
18.2: Especiación
- Page ID
- 57333

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Una de las mejores definiciones de especies es la del biólogo evolutivo Ernst Mayr: “Una especie es una población real o potencialmente entrelazada que no se cruza con otras poblaciones de este tipo cuando hay oportunidad de hacerlo”. Sin embargo, a veces la cría puede tener lugar (como puede ocurrir entre un caballo y un burro) pero si es así, las crías no son tan fértiles y/o bien adaptadas como los padres (la mula producida es estéril).
Especiación alopátrica: el papel del aislamiento en la especiación
La formación de dos o más especies a menudo (¡algunos trabajadores piensan siempre!) requiere el aislamiento geográfico de las subpoblaciones de la especie. Sólo entonces la selección natural o tal vez la deriva genética puede producir reservas genéticas distintivas. No es casualidad que las diversas razas (o "subespecies “) de los animales casi nunca ocupen el mismo territorio. Su distribución es alopátrica (“otro país”).

Las siete subespecies o razas distintas de la garganta amarilla Geothlypis trichas (una curruca) en Estados Unidos continental pronto se fusionarían en una sola población homogénea si ocupaban el mismo territorio y se criaban entre sí.
Pinzones de Darwin
De joven de 26 años, Charles Darwin visitó las Islas Galápagos frente a la costa de Ecuador. Entre los animales que estudió se encontraban lo que parecían ser 13 especies * de pinzones que no se encuentran en ningún otro lugar de la tierra.
- Algunos tienen picos fuertes para comer semillas de un tamaño u otro (#2, #3, #6).
- Otros tienen picos adaptados para comer insectos o néctar.
- Uno (#7) tiene pico como el de un pájaro carpintero. Lo usa para perforar agujeros en madera, pero al carecer de la larga lengua de un verdadero pájaro carpintero, utiliza una espina de cactus sostenida en su pico para excavar al insecto.
- Uno (#12) se parece más a una curruca que a un pinzón, pero sus huevos, nido y comportamiento de cortejo es como el de los otros pinzones.

Pinzones de Darwin. Los pinzones numerados 1—7 son pinzones molidos. Buscan su alimento en el suelo o en arbustos bajos. Los numerados 8—13 son pinzones arbóreos. Viven principalmente de insectos.
1. Pinzón grande de cactus (Geospiza conirostris)
2. Pinzón de tierra grande (Geospiza magnirostris)
3. Pinzón medio molido (Geospiza fortis)
4. Pinzón cactus (Geospiza scandens)
5. Pinzón molido de pico afilado (Geospiza difficilis)
6. Pinzón molido pequeño (Geospiza fuliginosa)
7. Pinzón pájaro carpintero (Cactospiza pallida)
8. Pinzón arbóreo vegetariano (Platyspiza crassirostris)
9. Pinzón arbóreo mediano (Camarhynchus pauper)
10. Pinzón arbóreo grande (Camarhynchus psittacula)
11. Pinzón arbóreo pequeño (Camarhynchus parvulus)
12. Pinzón curruca (Certhidia olivacea)
13. Pinzón de manglar (Cactospiza heliobates)
(De BSCS, Ciencias Biológicas: Moléculas al Hombre, Houghton Mifflin Co., 1963)
* El análisis genético proporciona evidencia de que:
- En realidad hay dos especies de pinzón curruca: Certhidia olivacea ahora llamado pinzón curruca verde y Certhidia fusca, el pinzón curruca gris.
- Las diversas poblaciones de Geospiza difficilis que se encuentran en las diferentes islas pertenecen a uno u otro de tres clados tan genéticamente distintos que merecen el estatus de especie completo.
Ya sea que el número sea 13 o 17, desde la época de Darwin, estas aves han proporcionado un estudio de caso de cómo una sola especie que llega a las Galápagos desde Centroamérica o Sudamérica podría -a lo largo de unos pocos millones de años- dar lugar a las diversas especies que viven allí hoy en día. Se han identificado varios factores que pueden contribuir a la especiación.
Oportunidad ecológica
Cuando el antepasado de los pinzones de Darwin llegó a las Galápagos, no encontró depredadores (No había mamíferos y pocos reptiles en las islas.) y pocos, si los hay, competidores. Sólo había un puñado de otros tipos de pájaros cantores. De hecho, si hubieran estado presentes verdaderas currucas o pájaros carpinteros, su eficiencia en la explotación de sus nichos seguramente habría impedido la evolución de pinzones currucas y pájaros carpinteros.
Aislamiento geográfico (alopatría)
La proximidad de las diversas islas ha permitido una migración suficiente de pinzones de Darwin entre ellas para permitir que surjan distintas poblaciones insulares. Pero las distancias entre las islas son lo suficientemente grandes como para limitar el mestizaje entre poblaciones en diferentes islas. Esto ha hecho posible la formación de subespecies distintivas (= razas) en las diversas islas.
La importancia del aislamiento geográfico está iluminada por una sola, decimocuarta, especie de pinzones de Darwin que vive en la Isla de los Cocos, a unas 500 millas (800 km) al noreste de las Galápagos. Los primeros inmigrantes allí también deben haber encontrado presiones de selección relajadas con pocos depredadores o competidores. Sin embargo, qué diferente es el resultado. Donde una especie inmigrante dio lugar a al menos 13 en las islas Galápagos dispersas, no se ha producido tal divergencia en la única isla aislada del Cocos.
Cambio evolutivo
De forma aislada, los cambios en el acervo genético pueden ocurrir a través de alguna combinación de selección natural, deriva genética y efecto fundador. Estos factores pueden producir distintas subpoblaciones en las diferentes islas. Mientras permanezcan separados (alopátricos) los consideramos razas o subespecies. De hecho, tal vez no puedan cruzarse con otras razas pero mientras no sepamos, asumimos que podrían.
¿Cuánto cambio genético se necesita para crear una nueva especie? Quizás no tanto como se podría pensar. Por ejemplo, los cambios en uno o solo unos pocos loci génicos podrían hacer el truco. Por ejemplo, una sola mutación alterando el color de la flor o la forma del pétalo podría impedir inmediatamente la polinización cruzada entre los tipos nuevos y los parentales (una forma de mecanismo de aislamiento precigótico).
Reunion
La cuestión de su estatus -subespecie o especie verdadera- se resuelve si alguna vez vuelven a ocupar el mismo territorio (se vuelven sintomáticos). Si se produce un mestizaje exitoso, las diferencias desaparecerán gradualmente, y se volverá a formar una sola población. La especiación no habrá ocurrido. Si, por otro lado, dos subespecies se reúnen pero no logran reanudar la reproducción, se ha producido la especiación y se han convertido en especies separadas.
Un ejemplo: El pinzón arbóreo mediano Camarhynchus pauper se encuentra sólo en la isla Floreana. Su pariente cercano, el pinzón arbóreo grande, Camarhynchus psittacula, se encuentra en todas las islas centrales incluyendo Floreana. De no ser por su presencia en Floreana, ambas formas serían consideradas subespecies de la misma especie. Debido a que sí conviven y mantienen su identidad separada en Floreana, sabemos que la especiación ha ocurrido.
Mecanismos de aislamiento
¿Qué podría impedir que dos subpoblaciones se crucen cuando se reúnan geográficamente? Existen varios mecanismos.
Los Mecanismos de Aislamiento Precigótico actúan antes de que ocurra la fertilización. Selección sexual - un fracaso para provocar el comportamiento de apareamiento. En Floreana, Camarhynchus psittacula tiene un pico más largo que Camarhynchus pauper, y los equipos de investigación dirigidos por Peter y Rosemary Grant han demostrado que el tamaño del pico es un criterio importante por el cual los pinzones de Darwin eligen a sus compañeros. Dos subpoblaciones pueden ocupar diferentes hábitats en la misma zona y, por lo tanto, no se encuentran en el momento de la reproducción. En las plantas, un cambio en el tiempo de floración puede impedir la polinización entre las dos subpoblaciones. Las diferencias estructurales en los órganos sexuales pueden convertirse en un mecanismo de aislamiento. El esperma puede no alcanzar o fusionarse con el óvulo.
Los Mecanismos de Aislamiento Postcigótico actúan incluso si se produce la fertilización. Incluso si se forma un cigoto, las diferencias genéticas pueden haberse vuelto tan grandes que los híbridos resultantes son menos viables o menos fértiles que los tipos parentales. La mula estéril producida al aparearse un caballo con un burro es un ejemplo. La esterilidad en los machos producida por hibridación es más común que en las hembras. De hecho, es el mecanismo de aislamiento postcigótico más común. Cuando Drosophila melanogaster intenta aparearse con su pariente Drosophila simulans, ni siquiera se producen machos viables. Las mutaciones en un solo gen (que codifica un componente del complejo de poro nuclear) son las responsables.
Refuerzo
Cuando dos especies que se han separado en alopatría se reúnen, sus mecanismos de aislamiento precigótico y postcigótico pueden volverse más estrictos que aquellos entre las mismas especies que existen separados entre sí. A este fenómeno se le llama refuerzo. Surge de la selección natural trabajando para favorecer a individuos que evitan apareamientos interespecíficos, lo que produciría híbridos menos aptos, cuando las dos especies se reúnen por primera vez.
Especiación por Hibridación
La hibridación entre angiospermas relacionadas a veces va seguida de una duplicación del número de cromosomas. Los poliploides resultantes ahora son completamente fértiles entre sí aunque no pueden reproducirse con ninguno de los dos tipos parentales; se ha creado una nueva especie. Esto parece haber sido un mecanismo frecuente de especiación en angiospermas. Incluso sin formar un poliploide, la hibridación interespecífica puede conducir ocasionalmente a una nueva especie de angiosperma. Dos especies de girasol, el “girasol común”, Helianthus annuus, y el “girasol de la pradera”, H. petiolaris, crecen ampliamente en la mitad occidental de Estados Unidos. Pueden cruzarse, pero sólo en raras ocasiones se producen crías fértiles.
Sin embargo, Rieseberg y sus colegas han encontrado evidencia de que la hibridación exitosa entre ellos ha ocurrido naturalmente en el pasado. Han demostrado que otras tres especies de girasol (cada una creciendo en un hábitat demasiado duro para cualquiera de los dos tipos parentales) son cada una producto de una antigua hibridación entre Helianthus annuus y H. petiolaris. Aunque cada una de estas especies tiene el mismo número diploide de cromosomas que los progenitores (2 n = 34), cada una tiene un patrón de segmentos cromosómicos que se han derivado, por recombinación genética, de ambos genomas parentales. Lo demostraron de varias maneras, notablemente combinando el análisis RFLP con la reacción en cadena de la polimerasa (PCR).
Incluso llegaron a producir (a baja frecuencia) híbridos annuus x petiolaris en el invernadero que imitaban los fenotipos y genotipos de los híbridos naturales. (Estos estudios monumentales fueron descritos en la edición del 29 de agosto de 2003 de Science.)
Otro ejemplo. En Pensilvania, los híbridos entre una especie de mosca de la fruta (no Drosophila) que se alimenta de arándanos y otra especie (de nuevo, no Drosophila) que se alimenta de bayas de nieve se alimentan de madreselva donde no encuentran competencia de sus especies parentales ni tienen oportunidad de reproducirse con ellas (no introgresión). Este estudio fue publicado en la edición del 28 de julio de 2005 de Nature. Por lo que la especiación puede ocurrir como resultado de la hibridación entre dos especies relacionadas, si el híbrido
- recibe un genoma que le permite reproducirse con otros híbridos similares pero no reproducirse con ninguna de las especies parentales,
- puede escapar a un hábitat donde no tiene que competir con ninguno de los padres,
- está adaptado para vivir bajo esas nuevas condiciones.
Radiación Adaptativa
Los procesos descritos en esta página pueden ocurrir una y otra vez. En el caso de los pinzones de Darwin, deben haberse repetido varias veces formando nuevas especies que dividieron gradualmente los hábitats disponibles entre ellos. Desde la primera llegada han llegado una variedad de pinzones que se alimentan de tierra y que se alimentan de árboles, así como el pinzón parecido a la curruca y el pinzón carpintero que usa herramientas. La formación de una serie de especies diversas a partir de una sola ancestral se llama radiación adaptativa.
Un informe publicado en la edición del 13 de enero de 2000 de Nature describe un estudio de las poblaciones de ratones domésticos en la isla de Madeira frente a la costa noroeste de África. Estos trabajadores (Janice Britton-Davidian et al) examinaron los cariotipos de 143 ratones caseros (Mus musculus domesticus) de diversos lugares a lo largo de la costa de esta isla montañosa.
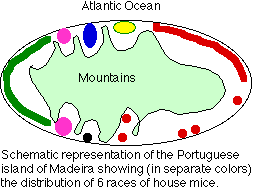
Sus hallazgos:
- Hay 6 poblaciones distintas (mostradas por diferentes colores)
- Cada uno de estos tiene un cariotipo distinto, con un número diploide menor que el “normal” (2 n = 40).
- La reducción del número de cromosomas se ha producido a través de fusiones Robertsonias. Los cromosomas de ratón tienden a ser acrocéntricos; es decir, el centrómero conecta un brazo largo y otro muy corto. Los cromosomas acrocéntricos están en riesgo de translocaciones que fusionan los brazos largos de dos cromosomas diferentes con la pérdida de los brazos cortos.
- Las diferentes poblaciones son alopátricas; aisladas en diferentes valles que conducen al mar.
- El cariotipo distinto y uniforme encontrado en cada población probablemente surgió de la deriva genética más que de la selección natural.
- Las 6 poblaciones diferentes son técnicamente descritas como razas porque no hay oportunidad para que intenten el mestizaje.
- Sin embargo, seguramente cumplen con la definición de especies verdaderas. Si bien los híbridos se formarían fácilmente (sin mecanismos de aislamiento precigóticos), estos probablemente serían infértiles ya que la sinapsis adecuada y la segregación de cromosomas tan diferentes sería difícil cuando los híbridos intentaran formar gametos por meiosis.
Especiación simpátrica
Especiación simpátrica se refiere a la formación de dos o más especies descendientes de una sola especie ancestral que ocupan la misma ubicación geográfica. Algunos biólogos evolutivos no creen que alguna vez ocurra. Sienten que el mestizaje pronto eliminaría cualquier diferencia genética que pudiera aparecer. Pero hay alguna evidencia convincente (aunque indirecta) de que la especiación simpátrica puede ocurrir.
Las espinillas de tres espinas, peces de agua dulce que han sido estudiados por Dolph Schluter (quien recibió su doctorado por su trabajo sobre los pinzones de Darwin con Peter Grant) y sus actuales colegas en Columbia Británica, proporcionan un ejemplo intrigante que mejor se explica por la especiación simpátrica.
Han encontrado:
- Dos especies diferentes de espinillas espinosas en cada uno de cinco lagos diferentes.
- una especie bentónica grande de boca grande que se alimenta de presas grandes en la zona litoral
- una especie limnética más pequeña con una boca más pequeña que se alimenta del plancton pequeño en aguas abiertas.
- El análisis de ADN indica que cada lago fue colonizado de forma independiente, presumiblemente por un antepasado marino, después de la última edad de hielo.
- El análisis de ADN también muestra que las dos especies en cada lago están más estrechamente relacionadas entre sí que con cualquiera de las especies en los otros lagos.
- Sin embargo, las dos especies en cada lago están aisladas reproductivamente; ninguna se aparea con la otra.
- Sin embargo, las pruebas en acuarios mostraron que
- Las especies bentónicas de un lago desovan con las especies bentónicas de los otros lagos y de igual manera las especies limneticas de los diferentes lagos desovan entre sí.
- Estas especies bentónicas y limnéticas incluso muestran sus preferencias de apareamiento cuando se presentan con sticklebacks de lagos japoneses; es decir, un béntico canadiense prefiere un bentónico japonés sobre su primo limnético cercano de su propio lago.
- Su conclusión: en cada lago, lo que comenzó como una sola población enfrentó tal competencia por recursos limitados que
- selección disruptiva: competencia que favorece a los peces en cualquier extremo del tamaño corporal y el tamaño de la boca sobre aquellos más cercanos a la media, junto con
- apareamiento surtido: cada tamaño preferido se aparea como él
- El hecho de que este patrón de especiación ocurrió de la misma manera en tres ocasiones distintas sugiere fuertemente que los factores ecológicos en una población simpátrica pueden causar especiación.
La especiación simpátrica impulsada por factores ecológicos también puede explicar la extraordinaria diversidad de crustáceos que viven en las profundidades del lago Baikal de Siberia.
¿Cuántos genes se necesitan para comenzar el camino hacia la especiación simpátrica?
Quizás no muchos. El barrenador europeo del maíz, Ostrinia nubilalis, (que a pesar de su nombre común también es una plaga importante en Estados Unidos) existe como dos razas distintas designadas Z y E. Ambos se pueden encontrar en la misma zona; es decir, son sintomáticos. Pero en el campo, practican el apareamiento selectivo, solo criándose con compañeros de su propia raza.
Las hembras de ambas razas sintetizan y liberan una feromona que es un atrayente sexual para los machos. Ambas razas utilizan la misma sustancia pero diferentes isómeros de la misma. El isómero que se produce está bajo el control de un locus génico que codifica una sola enzima. La capacidad de los machos para responder a un isómero u otro está controlada por 2 loci.
El problema de los clines
Existe otra forma posible de que surjan nuevas especies ante la ausencia de barreras geográficas. Si una población abarca un área grande y si los individuos de esa población pueden dispersarse solo en una pequeña porción de este rango, entonces el flujo génico a través de estas grandes distancias se reduciría. La ocurrencia de diferencias fenotípicas graduales (y genotípicas) en una población a través de una gran área geográfica se llama clina. El mestizaje exitoso ocurre libremente en cada punto a lo largo de la clina, pero los individuos en los extremos de la clina pueden no ser capaces de cruzarse. Esto se puede probar en el laboratorio.
Y, en ocasiones, se prueba en la naturaleza. Si una clina se dobla para que los extremos se encuentren, y las poblaciones reunidas en el cruce no puedan cruzarse, entonces se ha cumplido la definición de especie separada. Estas especies se denominan especies de anillo y este tipo de especiación se llama especiación parapátrica.
Dos ejemplos:
1. La zapatilla caribeña tártago Euphorbia tithymaloides.
El análisis genético muestra que esta flor silvestre se originó en Centroamérica donde México y Guatemala comparten un límite común. A partir de ahí se extendió en dos direcciones
- al noreste a través de la península de Yucatán y luego saltó de isla a través de Jamaica, República Dominicana, Puerto Rico y hacia las Islas Vírgenes;
- al sur a través de Centroamérica, a través de Venezuela, y luego al norte a través de Barbados y las otras islas de las Antillas Menores finalmente llegando también a las Islas Vírgenes.
Reunidas en las Islas Vírgenes, las dos poblaciones han divergido lo suficiente como para conservar sus rasgos genotípicos y fenotípicos distintivos. Los estudios en curso determinarán en qué grado pueden estar aislados reproductivamente.
2. La salamandra de gran mancha Ensatina eschscholtzii.
Este animal se encuentra en California donde se presenta en una serie de diferentes subespecies o razas. Una sola subespecie se encuentra en el norte de California, y se cree que es la fundadora de todas las demás. Con el tiempo esa población original se extendió hacia el sur en dos direcciones:
- bajando las montañas de Sierra Nevada al este del gran valle central de San Joaquín y
- bajando por la cordillera de la costa al oeste del valle.
Al sur del valle, el grupo oriental se ha movido hacia el oeste y ahora se encuentra con el grupo occidental, cerrando el anillo. Aquí las dos poblaciones no logran cruzarse con éxito, manteniendo sus distintas identidades. Pero cada subespecie se cruza en una cadena ininterrumpida por los dos caminos que tomaron sus antepasados.
Las especies anulares presentan un problema difícil en la asignación de designaciones de especies. Es fácil decir que las poblaciones en los extremos del clino representan especies separadas, pero ¿dónde dio lugar una a la otra? En cada punto a lo largo de la clina, el mestizaje continúa con éxito.
El mismo problema se enfrenta a los paleontólogos que examinan los cambios fenotípicos graduales observados en una línea continua de fósiles cada vez más jóvenes de lo que se supone que es una sola línea de descenso. Si se pudiera resucitar la especie ancestral (A) y la especie descendiente (B) y no podían cruzarse, entonces cumplen con la definición de especie separada. Pero no hubo momento en el tiempo en el que se pudiera decir que A se convirtió en B. Así que los clines de hoy son un modelo en el espacio del descenso de Darwin con modificaciones ocurriendo a lo largo del tiempo.
Aunque los clines presentan un problema para los clasificadores, son una hermosa demostración de la convicción de Darwin de que la acumulación de pequeñas diferencias heredadas puede conducir a la formación de nuevas especies.