
30.6: Sistemas sensoriales y respuestas de plantas
- Page ID
- 59320

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Habilidades para Desarrollar
- Describir cómo la luz roja y azul afectan el crecimiento vegetal y las actividades metabólicas
- Discutir el gravitropismo
- Entender cómo las hormonas afectan el crecimiento y desarrollo de las plantas
- Describir el tigmotropismo, el tigmonastismo y la tigmogénesis
- Explicar cómo las plantas se defienden de los depredadores y responden a las heridas
Los animales pueden responder a factores ambientales al mudarse a una nueva ubicación. Las plantas, sin embargo, están enraizadas en su lugar y deben responder a los factores ambientales circundantes. Las plantas cuentan con sistemas sofisticados para detectar y responder a la luz, la gravedad, la temperatura y el tacto físico. Los receptores detectan los factores ambientales y transmiten la información a los sistemas efectores, a menudo a través de mensajeros químicos intermedios, para generar respuestas de las plantas.
Respuestas de las plantas a la luz
Las plantas tienen una serie de usos sofisticados para la luz que van mucho más allá de su capacidad para fotosintetizar azúcares de bajo peso molecular usando solo dióxido de carbono, luz y agua. La fotomorfogénesis es el crecimiento y desarrollo de las plantas en respuesta a la luz. Permite a las plantas optimizar su uso de la luz y el espacio. El fotoperiodismo es la capacidad de usar la luz para rastrear el tiempo. Las plantas pueden decir la hora del día y la época del año al detectar y usar varias longitudes de onda de la luz solar. El fototropismo es una respuesta direccional que permite que las plantas crezcan hacia, o incluso lejos de, la luz.
La detección de luz en el ambiente es importante para las plantas; puede ser crucial para la competencia y supervivencia. La respuesta de las plantas a la luz está mediada por diferentes fotorreceptores, los cuales están compuestos por una proteína unida covalentemente a un pigmento absorbente de luz llamado cromóforo. Juntos, a los dos se les llama cromoproteína.
Las regiones rojo/rojo lejano y violeta-azul del espectro de luz visible desencadenan el desarrollo estructural en las plantas. Los fotorreceptores sensoriales absorben la luz en estas regiones particulares del espectro de luz visible debido a la calidad de la luz disponible en el espectro de luz diurna. En hábitats terrestres, la absorción de luz por clorofilas alcanza su punto máximo en las regiones azul y roja del espectro. A medida que la luz se filtra a través del dosel y se absorben las longitudes de onda azul y roja, el espectro se desplaza hacia el extremo rojo lejano, desplazando la comunidad vegetal a aquellas plantas mejor adaptadas para responder a la luz roja lejana. Los receptores de luz azul permiten a las plantas medir la dirección y la abundancia de la luz solar, que es rica en emisiones azul-verde. El agua absorbe la luz roja, lo que hace que la detección de luz azul sea esencial para las algas y plantas acuáticas.
El Sistema Fitocromo y la Respuesta Rojo/Rojo Lejano
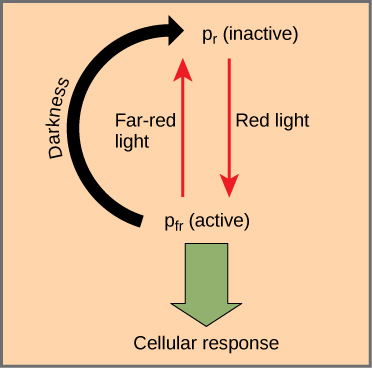
Los fitocromos son una familia de cromoproteínas con un cromóforo tetrapirrol lineal, similar al grupo principal de clorofila absorbente de luz de tetrapirrol anillado. Los fitocromos tienen dos formas foto-interconvertibles: Pr y Pfr. Pr absorbe la luz roja (~667 nm) y se convierte inmediatamente a Pfr. Pfr absorbe la luz roja extrema (~730 nm) y se convierte rápidamente de nuevo a Pr. La absorción de luz roja o roja lejana provoca un cambio masivo en la forma del cromóforo, alterando la conformación y actividad de la proteína fitocromática a la que está unida. Pfr es la forma fisiológicamente activa de la proteína; por lo tanto, la exposición a la luz roja produce actividad fisiológica. La exposición a la luz roja extrema inhibe la actividad del fitocromo. En conjunto, las dos formas representan el sistema de fitocromo (Figura\(\PageIndex{1}\)).
El sistema de fitocromo actúa como un interruptor de luz biológico. Monitorea el nivel, intensidad, duración y color de la luz ambiental. El efecto de la luz roja es reversible al brillar inmediatamente luz roja lejana sobre la muestra, lo que convierte la cromoproteína en la forma Pr inactiva. Además, Pfr puede volver lentamente a Pr en la oscuridad, o descomponerse con el tiempo. En todos los casos, se invierte la respuesta fisiológica inducida por la luz roja. La forma activa del fitocromo (Pfr) puede activar directamente otras moléculas en el citoplasma, o bien puede traficarse al núcleo, donde activa o reprime directamente la expresión génica específica.
Una vez que el sistema de fitocromo evolucionó, las plantas lo adaptaron para satisfacer una variedad de necesidades. Sin filtrar, la luz solar completa contiene mucha más luz roja que la luz roja extrema. Debido a que la clorofila absorbe fuertemente en la región roja del espectro visible, pero no en la región roja lejana, cualquier planta a la sombra de otra planta en el suelo del bosque estará expuesta a luz empobrecida en rojo y enriquecida en rojo lejano. La preponderancia de la luz roja lejana convierte el fitocromo en las hojas sombreadas a la forma Pr (inactiva), lo que ralentiza el crecimiento. Las áreas más cercanas no sombreadas (o incluso menos sombreadas) en el suelo del bosque tienen más luz roja; las hojas expuestas a estas áreas perciben la luz roja, lo que activa la forma Pfr e induce el crecimiento. En definitiva, los brotes de las plantas utilizan el sistema fitocromo para crecer lejos de la sombra y hacia la luz. Debido a que la competencia por la luz es tan feroz en una comunidad vegetal densa, las ventajas evolutivas del sistema fitocromo son obvias.
En las semillas, el sistema de fitocromo no se utiliza para determinar la dirección y calidad de la luz (sombreada versus no sombreada). En cambio, se utiliza simplemente para determinar si hay alguna luz en absoluto. Esto es especialmente importante en especies con semillas muy pequeñas, como la lechuga. Por su tamaño, las semillas de lechuga tienen pocas reservas de alimentos. Sus plántulas no pueden crecer por mucho tiempo antes de que se les quede sin combustible. Si germinaran incluso un centímetro bajo la superficie del suelo, la plántula nunca llegaría a la luz del sol y moriría. En la oscuridad, el fitocromo está en la Pr (forma inactiva) y la semilla no germinará; solo germinará si se expone a la luz en la superficie del suelo. Al exponerse a la luz, Pr se convierte en Pfr y procede la germinación.
Las plantas también utilizan el sistema fitocromo para detectar el cambio de estación. El fotoperiodismo es una respuesta biológica al momento y duración del día y de la noche. Controla la floración, el ajuste de los cogollos de invierno y el crecimiento vegetativo. La detección de cambios estacionales es crucial para la supervivencia de las plantas. Aunque la temperatura y la intensidad de la luz influyen en el crecimiento de las plantas, no son indicadores confiables de temporada porque pueden variar de un año a otro. La duración del día es un mejor indicador de la época del año.
Como se indicó anteriormente, la luz solar sin filtrar es rica en luz roja pero deficiente en luz roja lejana. Por lo tanto, al amanecer, todas las moléculas de fitocromo en una hoja se convierten rápidamente a la forma activa de Pfr, y permanecen en esa forma hasta el atardecer. En la oscuridad, la forma Pfr tarda horas en volver lentamente a la forma Pr. Si la noche es larga (como en invierno), toda la forma Pfr revierte. Si la noche es corta (como en verano), puede quedar una cantidad considerable de Pfr al amanecer. Al detectar la relación Pr/Pfr al amanecer, una planta puede determinar la duración del ciclo día/noche. Además, las hojas conservan esa información durante varios días, permitiendo una comparación entre la duración de la noche anterior y las noches anteriores. Las noches más cortas indican la primavera a la planta; cuando las noches se alargan, se acerca el otoño. Esta información, junto con la detección de temperatura y disponibilidad de agua, permite a las plantas determinar la época del año y ajustar su fisiología en consecuencia. Las plantas de día corto (noche larga) utilizan esta información para florecer a fines del verano y principios del otoño, cuando las noches superan una duración crítica (a menudo ocho horas o menos). Las plantas de día largo (noche corta) florecen durante la primavera, cuando la oscuridad es menor que una longitud crítica (a menudo de ocho a 15 horas). No todas las plantas utilizan el sistema fitocromo de esta manera. La floración en plantas de día neutro no está regulada por la duración del día.
Conexión de carrera: Horticulturalista
La palabra “horticultor” proviene de las palabras latinas para jardín (hortus) y cultura (cultura). Esta carrera ha sido revolucionada por los avances en la comprensión de las respuestas de las plantas a los estímulos ambientales. Los productores de cultivos, frutas, hortalizas y flores estaban previamente restringidos por tener que cronometrar su siembra y cosecha según la temporada. Ahora, los horticultores pueden manipular las plantas para aumentar la producción de hojas, flores o frutos al comprender cómo los factores ambientales afectan el crecimiento y desarrollo de las plantas.
El manejo de invernaderos es un componente esencial de la educación de un horticultor. Para alargar la noche, las plantas se cubren con un paño de sombra opaca. Las plantas de día largo se irradian con luz roja en invierno para promover la floración temprana. Por ejemplo, la luz fluorescente (blanca fría) alta en longitudes de onda azules fomenta el crecimiento frondoso y es excelente para iniciar plántulas. Las lámparas incandescentes (bombillas estándar) son ricas en luz roja y promueven la floración en algunas plantas. El momento de maduración de los frutos puede aumentarse o retrasarse mediante la aplicación de hormonas vegetales. Recientemente, se han logrado avances considerables en el desarrollo de razas de plantas adecuadas a diferentes climas y resistentes a plagas y daños en el transporte. Tanto el rendimiento como la calidad de los cultivos han aumentado como resultado de las aplicaciones prácticas del conocimiento de las respuestas de las plantas a estímulos externos y hormonas.
Los horticultores encuentran empleo en laboratorios privados y gubernamentales, invernaderos, jardines botánicos y en los campos de producción o investigación. Mejoran los cultivos aplicando sus conocimientos de genética y fisiología vegetal. Para prepararse para una carrera de horticultura, los estudiantes toman clases de botánica, fisiología vegetal, patología vegetal, diseño del paisaje y mejoramiento de plantas. Para complementar estos cursos tradicionales, las especializaciones de horticultura agregan estudios en economía, negocios, informática y comunicaciones.
Las respuestas de la luz azul
El fototropismo, la flexión direccional de una planta hacia o lejos de una fuente de luz, es una respuesta a las longitudes de onda azules de la luz. El fototropismo positivo es el crecimiento hacia una fuente de luz (Figura\(\PageIndex{2}\)), mientras que el fototropismo negativo (también llamado skototropismo) es crecimiento lejos de la luz.
Las fototropinas apropiadamente llamadas son receptores basados en proteínas responsables de mediar la respuesta fototrópica. Como todos los fotorreceptores vegetales, las fototropinas consisten en una porción de proteína y una porción absorbente de luz, llamada cromóforo. En las fototropinas, el cromóforo es una molécula de flavina unida covalentemente; de ahí que las fototropinas pertenecen a una clase de proteínas llamadas flavoproteínas.
Otras respuestas bajo el control de las fototropinas son la apertura y cierre de las hojas, el movimiento de los cloroplastos y la apertura de los estomas. Sin embargo, de todas las respuestas controladas por fototropinas, el fototropismo ha sido estudiado el más largo y es el mejor entendido.
En su tratado de 1880 El poder de los movimientos en las plantas, Charles Darwin y su hijo Francis describieron por primera vez el fototropismo como la flexión de las plántulas hacia la luz. Darwin observó que la luz era percibida por la punta de la planta (el meristemo apical), pero que la respuesta (flexión) se dio en una parte diferente de la planta. Concluyeron que la señal tuvo que viajar desde el meristemo apical hasta la base de la planta.

En 1913, Peter Boysen-Jensen demostró que una señal química producida en la punta de la planta era la responsable de la flexión en la base. Cortó la punta de una plántula, cubrió la sección cortada con una capa de gelatina, y luego reemplazó la punta. La plántula se inclinó hacia la luz cuando se iluminaba. Sin embargo, cuando se insertaron escamas de mica impermeables entre la punta y la base cortada, la plántula no se dobló. Un refinamiento del experimento mostró que la señal viajó en el lado sombreado de la plántula. Cuando se insertó la placa de mica en el lado iluminado, la planta sí se dobló hacia la luz. Por lo tanto, la señal química fue un estimulante del crecimiento debido a que la respuesta fototrópica implicó un alargamiento celular más rápido en el lado sombreado que en el lado iluminado. Ahora sabemos que a medida que la luz pasa a través del tallo de una planta, se difracta y genera activación de fototropina a través del tallo. La mayor parte de la activación ocurre en el lado iluminado, provocando que la hormona vegetal ácido indol acético (IAA) se acumule en el lado sombreado. Las células madre se alargan bajo la influencia de IAA.
Los criptocromos son otra clase de fotorreceptores absorbentes de luz azul que también contienen un cromóforo basado en flavina. Los criptocromos establecen el ciclo de actividad de 24 horas de las plantas, también conocido como su ritema circadiano, utilizando señales de luz azul. Existe cierta evidencia de que los criptocromos trabajan junto con fototropinas para mediar la respuesta fototrópica.
Enlace al aprendizaje
Utilice el menú de navegación en el panel izquierdo de este sitio web para ver imágenes de plantas en movimiento.
Respuestas de las plantas a la gravedad
Ya sea que germinen o no a la luz o en la oscuridad total, los brotes suelen brotar del suelo, y las raíces crecen hacia abajo hacia el suelo. Una planta colocada de costado en la oscuridad enviará brotes hacia arriba cuando se le dé suficiente tiempo. El gravitropismo asegura que las raíces crezcan en el suelo y que los brotes crezcan hacia la luz solar. El crecimiento de la punta apical del brote hacia arriba se llama gravitropismo negativo, mientras que el crecimiento de las raíces hacia abajo se llama gravitropismo positivo.
Los amiloplastos (también conocidos como estatolitos) son plastidios especializados que contienen gránulos de almidón y se asientan hacia abajo en respuesta a la gravedad. Los amiloplastos se encuentran en brotes y en células especializadas del casquete radicular. Cuando una planta se inclina, los estatolitos caen a la nueva pared celular del fondo. Unas horas después, el brote o raíz mostrará crecimiento en la nueva dirección vertical.
El mecanismo que media el gravitropismo es razonablemente bien entendido. Cuando los amiloplastos se asientan en el fondo de las células detectoras de gravedad en la raíz o brote, contactan físicamente con el retículo endoplásmico (ER), provocando la liberación de iones de calcio desde el interior de la sala de emergencias. Esta señalización de calcio en las células provoca el transporte polar de la hormona vegetal IAA al fondo de la célula. En las raíces, una alta concentración de IAA inhibe la elongación celular. El efecto ralentiza el crecimiento en el lado inferior de la raíz, mientras que las células se desarrollan normalmente en el lado superior. El IAA tiene el efecto contrario en los brotes, donde una mayor concentración en el lado inferior del brote estimula la expansión celular, haciendo que el brote crezca. Después de que el brote o raíz comienzan a crecer verticalmente, los amiloplastos vuelven a su posición normal. Se han propuesto otras hipótesis, que involucran a toda la célula en el efecto gravitropismo, para explicar por qué algunos mutantes que carecen de amiloplastos aún pueden exhibir una respuesta gravitropica débil.
Respuestas de crecimiento
La respuesta sensorial de una planta a estímulos externos se basa en mensajeros químicos (hormonas). Las hormonas vegetales afectan todos los aspectos de la vida vegetal, desde la floración hasta el cuajado y maduración del fruto y desde el fototropismo hasta la caída de las hojas. Potencialmente, cada célula de una planta puede producir hormonas vegetales. Pueden actuar en su célula de origen o ser transportados a otras porciones del cuerpo de la planta, con muchas respuestas vegetales que involucran la interacción sinérgica o antagónica de dos o más hormonas. En contraste, las hormonas animales se producen en glándulas específicas y se transportan a un sitio distante para la acción, y actúan solas.
Las hormonas vegetales son un grupo de sustancias químicas no relacionadas que afectan la morfogénesis de las plantas. Tradicionalmente se describen cinco hormonas vegetales principales: auxinas (particularmente IAA), citoquininas, giberelinas, etileno y ácido abscísico. Además, otros nutrientes y condiciones ambientales se pueden caracterizar como factores de crecimiento.
Auxins
El término auxina se deriva de la palabra griega auxein, que significa “crecer”. Las auxinas son las principales hormonas responsables del alargamiento celular en fototropismo y gravitropismo. También controlan la diferenciación del meristemo en tejido vascular, y promueven el desarrollo y la disposición de las hojas. Si bien muchas auxinas sintéticas se utilizan como herbicidas, IAA es la única auxina natural que muestra actividad fisiológica. La dominancia apical, la inhibición de la formación lateral de los cogollos, es desencadenada por las auxinas producidas en el meristema apical. La floración, el cuajado y maduración del fruto, y la inhibición de la abscisión (caída de hojas) son otras respuestas de las plantas bajo el control directo o indirecto de las auxinas. Las auxinas también actúan como relevo para los efectos de la luz azul y las respuestas rojas/rojo lejano.
El uso comercial de auxinas está muy extendido en viveros de plantas y para la producción de cultivos. El IAA se utiliza como hormona de enraizamiento para promover el crecimiento de raíces adventicias en esquejes y hojas desprendidas. La aplicación de auxinas sintéticas a plantas de tomate en invernaderos promueve el desarrollo normal del fruto. La aplicación al aire libre de auxina promueve la sincronización del fraguado y caída de frutos para coordinar la temporada de cosecha Frutos como los pepinos sin semillas pueden inducirse a dar frutos mediante el tratamiento de flores de plantas no fertilizadas con auxinas.
Citininas
El efecto de las citoquininas se reportó por primera vez cuando se encontró que agregar el endospermo líquido de cocos al desarrollo de embriones vegetales en cultivo estimuló su crecimiento. Se encontró que el factor de crecimiento estimulante era la citoquinina, una hormona que promueve la citocinesis (división celular). Hasta la fecha se conocen casi 200 citoquininas naturales o sintéticas. Las citoquininas son más abundantes en los tejidos en crecimiento, como raíces, embriones y frutos, donde se produce la división celular. Se sabe que las citoquininas retrasan la senescencia en los tejidos foliares, promueven la mitosis y estimulan la diferenciación del meristemo en brotes y raíces. Muchos efectos sobre el desarrollo de las plantas están bajo la influencia de las citoquininas, ya sea en conjunto con la auxina u otra hormona. Por ejemplo, la dominancia apical parece ser el resultado de un equilibrio entre las auxinas que inhiben los cogollos laterales, y las citoquininas que promueven el crecimiento más ocupado.
Giberelinas
Las giberelinas (GA) son un grupo de alrededor de 125 hormonas vegetales estrechamente relacionadas que estimulan el alargamiento de los brotes, la germinación de las semillas y la maduración de frutos y flores. Los GAs se sintetizan en los meristemos apicales de raíz y tallo, hojas jóvenes y embriones de semillas. En áreas urbanas, a veces se aplican antagonistas de GA a árboles bajo líneas eléctricas para controlar el crecimiento y reducir la frecuencia de poda.
Los GAs rompen la latencia (un estado de crecimiento y desarrollo inhibido) en las semillas de plantas que requieren exposición al frío o la luz para germinar. El ácido abscísico es un fuerte antagonista de la acción de GA. Otros efectos de los GAs incluyen la expresión de género, el desarrollo de frutos sin semillas y el retraso de la senescencia en hojas y frutos. Las uvas sin semillas se obtienen a través de métodos de reproducción estándar y contienen semillas poco visibles que no se desarrollan. Debido a que los GA son producidos por las semillas, y debido a que el desarrollo del fruto y la elongación del tallo están bajo control de GA, estas variedades de uvas normalmente producirían frutos pequeños en racimos compactos. Las uvas en maduración se tratan rutinariamente con GA para promover un mayor tamaño de fruto, así como racimos más sueltos (tallos más largos), lo que reduce el caso de infección por moho (Figura\(\PageIndex{3}\)).

Ácido abscísico
La hormona vegetal ácido abscísico (ABA) se descubrió por primera vez como el agente que causa la abscisión o caída de las cápsulas de algodón. Sin embargo, estudios más recientes indican que el ABA juega solo un papel menor en el proceso de abscisión. El ABA se acumula como respuesta a condiciones ambientales estresantes, como deshidratación, temperaturas frías o duraciones diarias acortadas. Su actividad contrarresta muchos de los efectos promotores del crecimiento de las GAs y las auxinas. ABA inhibe la elongación del tallo e induce latencia en cogollos laterales.
ABA induce la latencia en las semillas al bloquear la germinación y promover la síntesis de proteínas de almacenamiento. Las plantas adaptadas a climas templados requieren un largo periodo de temperatura fría antes de que las semillas germinen. Este mecanismo protege a las plantas jóvenes de brotar demasiado pronto durante un clima inestacional cálido en invierno. A medida que la hormona se descompone gradualmente durante el invierno, la semilla se libera de la latencia y germina cuando las condiciones son favorables en primavera. Otro efecto de ABA es promover el desarrollo de cogollos de invierno; media la conversión del meristema apical en un cogollo latente. La baja humedad del suelo provoca un aumento en el ABA, lo que hace que los estomas se cierren, reduciendo la pérdida de agua en los brotes
Etileno
El etileno se asocia con maduración de frutos, marchitamiento de flores y caída de hojas. El etileno es inusual porque es un gas volátil (C 2 H 4). Hace cientos de años, cuando se instalaron farolas de gas en las calles de la ciudad, los árboles que crecieron cerca de los postes de las lámparas desarrollaron troncos retorcidos, engrosados y arrojaron sus hojas antes de lo esperado. Estos efectos fueron causados por la volatilización del etileno de las lámparas.
Los tejidos envejecidos (especialmente las hojas senescentes) y los nódulos de los tallos producen etileno. El efecto más conocido de la hormona, sin embargo, es la promoción de la maduración de los frutos. El etileno estimula la conversión de almidón y ácidos en azúcares. Algunas personas almacenan fruta inmadura, como aguacates, en una bolsa de papel sellada para acelerar la maduración; el gas liberado por la primera fruta en madurar acelerará la maduración del fruto restante. El etileno también desencadena la abscisión de hojas y frutos, la decoloración y caída de las flores, y promueve la germinación en algunos cereales y la brotación de bulbos y papas.
El etileno es ampliamente utilizado en la agricultura. Los fruticultores comerciales controlan el momento de maduración de los frutos con la aplicación del gas. Los horticultores inhiben la caída de hojas en plantas ornamentales mediante la eliminación de etileno de los invernaderos mediante ventiladores y ventilación.
Hormonas no tradicionales
Investigaciones recientes han descubierto una serie de compuestos que también influyen en el desarrollo de las plantas. Sus papeles son menos entendidos que los efectos de las principales hormonas descritas hasta ahora.
Los jasmonatos juegan un papel importante en las respuestas de defensa a la herbivoría. Sus niveles aumentan cuando una planta es herida por un depredador, resultando en un aumento de metabolitos secundarios tóxicos. Contribuyen a la producción de compuestos volátiles que atraen a los enemigos naturales de los depredadores. Por ejemplo, la masticación de plantas de tomate por las orugas conduce a un aumento en los niveles de ácido jasmónico, lo que a su vez desencadena la liberación de compuestos volátiles que atraen a los depredadores de la plaga.
Las oligosacarinas también juegan un papel en la defensa de las plantas contra infecciones bacterianas y fúngicas. Actúan localmente en el sitio de la lesión, y también pueden ser transportados a otros tejidos. Las strigolactonas promueven la germinación de semillas en algunas especies e inhiben el desarrollo apical lateral en ausencia de auxinas. Las strigolactonas también juegan un papel en el establecimiento de micorrizas, una asociación mutualista de raíces de plantas y hongos. Los brasinoesteroides son importantes para muchos procesos fisiológicos y de desarrollo. Las señales entre estos compuestos y otras hormonas, notablemente auxina y GA, amplifican su efecto fisiológico. La dominancia apical, la germinación de semillas, el gravitropismo y la resistencia a la congelación están influenciados positivamente por las hormonas. El crecimiento radicular y la caída de frutos son inhibidos por los esteroides
Respuestas de Plantas a Viento y Tacto
El brote de una planta de guisante serpentea alrededor de un enrejado, mientras que un árbol crece en ángulo en respuesta a los fuertes vientos predominantes. Estos son ejemplos de cómo las plantas responden al tacto o al viento.
El movimiento de una planta sometida a presión direccional constante se llama tigmotropismo, de las palabras griegas thigma que significa “toque”, y tropismo que implica “dirección”. Los zaricillos son un ejemplo de ello. La región meristemática de los zarcillos es muy sensible al tacto; el tacto ligero evocará una respuesta rápida de enrollamiento. Las celdas en contacto con una superficie de soporte se contraen, mientras que las celdas en el lado opuesto del soporte se expanden. La aplicación de ácido jasmónico es suficiente para desencadenar el enrollamiento de zarcillos sin un estímulo mecánico.
Una respuesta tigmonástica es una respuesta táctil independiente de la dirección del estímulo. En el atrapamoscas Venus, dos hojas modificadas se unen en una bisagra y se alinean con púas delgadas en forma de tenedor a lo largo de los bordes exteriores. Los pelos diminutos se encuentran dentro de la trampa. Cuando un insecto se roza contra estos pelos de gatillo, tocando dos o más de ellos sucesivamente, las hojas se cierran rápidamente, atrapando a la presa. Las glándulas en la superficie foliar secretan enzimas que digieren lentamente al insecto. Los nutrientes liberados son absorbidos por las hojas, las cuales reabren para la siguiente comida.
La tigmomorfogénesis es un cambio de desarrollo lento en la forma de una planta sometida a estrés mecánico continuo. Cuando los árboles se doblan al viento, por ejemplo, el crecimiento suele atrofiarse y el tronco se espesa. El tejido fortalecedor, especialmente el xilema, se produce para agregar rigidez y resistir la fuerza del viento. Los investigadores plantean la hipótesis de que la tensión mecánica induce el crecimiento y la diferenciación para fortalecer los tejidos. El etileno y el jasmonato probablemente están involucrados en la tigmomorfogénesis.
Enlace al aprendizaje

Usa el menú de la izquierda para navegar a tres cortometrajes: una trampa para moscas Venus capturando presas, el cierre progresivo de folíolos sensibles de plantas y el entrelazado de zarcillos.
Respuestas de defensa contra herbívoros y patógenos
Las plantas se enfrentan a dos tipos de enemigos: herbívoros y patógenos. Los herbívoros tanto grandes como pequeños usan plantas como alimento, y las mastican activamente. Los patógenos son agentes de enfermedades. Estos microorganismos infecciosos, como hongos, bacterias y nematodos, viven de la planta y dañan sus tejidos. Las plantas han desarrollado una variedad de estrategias para desalentar o matar a los atacantes.
La primera línea de defensa en plantas es una barrera intacta e impenetrable. La corteza y la cutícula cerosa pueden proteger contra los depredadores. Otras adaptaciones contra la herbivoría incluyen las espinas, que son ramas modificadas, y las espinas, que son hojas modificadas. Desalientan a los animales al causar daño físico e inducir erupciones y reacciones alérgicas. La protección exterior de una planta puede verse comprometida por daños mecánicos, que pueden proporcionar un punto de entrada para patógenos. Si se incumple la primera línea de defensa, la planta debe recurrir a un conjunto diferente de mecanismos de defensa, como toxinas y enzimas.
Los metabolitos secundarios son compuestos que no se derivan directamente de la fotosíntesis y no son necesarios para la respiración o el crecimiento y desarrollo de las plantas. Muchos metabolitos son tóxicos, e incluso pueden ser letales para los animales que los ingieren. Algunos metabolitos son alcaloides, que desalientan a los depredadores con olores dañinos (como los aceites volátiles de menta y salvia) o sabores repelentes (como el amargor de la quinina). Otros alcaloides afectan a los herbívoros al provocar ya sea una estimulación excesiva (la cafeína es un ejemplo) o el letargo asociado a los opioides. Algunos compuestos se vuelven tóxicos después de la ingestión; por ejemplo, el cianuro de glicol en la raíz de yuca libera cianuro solo al ingerirlo por el herbívoro.
Las heridas mecánicas y los ataques de depredadores activan mecanismos de defensa y protección tanto en el tejido dañado como en sitios más alejados de la ubicación de la lesión. Algunas reacciones de defensa ocurren en cuestión de minutos: otras a lo largo de varias horas. Las células infectadas y circundantes pueden morir, deteniendo así la propagación de la infección.
La señalización de larga distancia provoca una respuesta sistémica dirigida a disuadir al depredador. A medida que se daña el tejido, los jasmonatos pueden promover la síntesis de compuestos que son tóxicos para los depredadores. Los jasmonatos también provocan la síntesis de compuestos volátiles que atraen a los parasitoides, que son insectos que pasan sus etapas de desarrollo en o sobre otro insecto, y eventualmente matan a su huésped. La planta puede activar la abscisión del tejido lesionado si se daña más allá de su reparación.
Resumen
Las plantas responden a la luz por cambios en la morfología y actividad. La irradiación con luz roja convierte el fotorreceptor fitocromo a su forma de absorción de luz roja lejana: PFR. Esta forma controla la germinación y la floración en respuesta a la duración del día, así como desencadena la fotosíntesis en plantas latentes o aquellas que acaban de emerger del suelo. Los receptores de luz azul, los criptocromos y las fototropinas son responsables del fototropismo. Los amiloplastos, que contienen gránulos pesados de almidón, perciben la gravedad. Los brotes presentan gravitropismo negativo, mientras que las raíces presentan gravitropismo positivo. Las hormonas vegetales, compuestos naturales sintetizados en pequeñas cantidades, pueden actuar tanto en las células que las producen como en tejidos y órganos distantes. Las auxinas son responsables de la dominancia apical, el crecimiento radicular, el crecimiento direccional hacia la luz y muchas otras respuestas de crecimiento. Las citoquininas estimulan la división celular y contrarrestan la dominancia apical en brotes. Las giberelinas inhiben la latencia de las semillas y promueven el crecimiento del tallo. El ácido abscísico induce la latencia en semillas y brotes, y protege a las plantas de la pérdida excesiva de agua al promover el cierre estomático. El gas etileno acelera la maduración de los frutos y la caída de las hojas. Las plantas responden al tacto por movimientos rápidos (tigmotropía y tigmonastía) y crecimiento diferencial lento (tigmomorfogénesis). Las plantas han desarrollado mecanismos de defensa contra depredadores y patógenos. Barreras físicas como corteza y espinas protegen los tejidos tiernos. Las plantas también tienen defensas químicas, incluyendo metabolitos secundarios tóxicos y hormonas, que provocan mecanismos de defensa adicionales.
Glosario
- ácido abscísico (ABA)
- hormona vegetal que induce la latencia en semillas y otros órganos
- abscisión
- proceso fisiológico que conduce a la caída de un órgano de la planta (como la caída de hojas o pétalos)
- auxina
- hormona vegetal que influye en el alargamiento celular (en fototropismo), gravitropismo, dominancia apical y crecimiento radicular
- cromóforo
- molécula que absorbe la luz
- criptocromo
- proteína que absorbe la luz en las regiones azul y ultravioleta del espectro de luz
- citoquinina
- hormona vegetal que promueve la división celular
- etileno
- hormona vegetal volátil que se asocia con la maduración de los frutos, el marchitamiento de las flores y la caída de las hojas
- giberelina (GA)
- hormona vegetal que estimula el alargamiento de los brotes, la germinación de las semillas y la maduración y caída de frutos y flores
- jasmonates
- pequeña familia de compuestos derivados del ácido graso ácido linoleico
- gravitropismo negativo
- crecimiento lejos de la gravedad de la Tierra
- oligosacarina
- hormona importante en las defensas de las plantas contra infecciones bacterianas y fúngicas
- fotomorfogénesis
- crecimiento y desarrollo de plantas en respuesta a la luz
- fotoperiodismo
- ocurrencia de procesos vegetales, como germinación y floración, según la época del año
- fototropina
- receptor de luz azul que promueve el fototropismo, apertura y cierre estomático, y otras respuestas que promueven la fotosíntesis
- fototropismo
- flexión direccional de una planta hacia una fuente de luz
- fitocromo
- proteína pigmentaria vegetal que existe en dos formas reversibles (Pr y Pfr) y media los cambios morfológicos en respuesta a la luz roja
- gravitropismo positivo
- crecimiento hacia el centro gravitacional de la Tierra
- statolith
- (también, amiloplasto) orgánulo vegetal que contiene gránulos pesados de almidón
- strigolactona
- hormona que promueve la germinación de semillas en algunas especies e inhibe el desarrollo apical lateral en ausencia de auxinas
- tigmomorfogénesis
- respuesta del desarrollo al tacto
- tigmonástico
- crecimiento direccional de una planta independientemente de la dirección en la que se aplique el contacto
- Thigmotropismo
- crecimiento direccional de una planta en respuesta al contacto constante