
22.3: Evolución adaptativa
- Page ID
- 58267
La selección natural solo actúa sobre los rasgos heredables de la población: seleccionar alelos beneficiosos y así aumentar su frecuencia en la población, al tiempo que selecciona contra alelos deletéreos y con ello disminuye su frecuencia, un proceso conocido como evolución adaptativa. La selección natural no actúa sobre alelos individuales, sin embargo, sino sobre organismos enteros. Un individuo puede portar un genotipo muy beneficioso con un fenotipo resultante que, por ejemplo, aumenta la capacidad de reproducción (fecundidad), pero si ese mismo individuo también porta un alelo que da como resultado una enfermedad infantil fatal, ese fenotipo de fecundidad no se transmitirá al siguiente generación porque el individuo no vivirá para llegar a la edad reproductiva. La selección natural actúa a nivel del individuo; selecciona para individuos con mayores contribuciones al acervo genético de la próxima generación, conocido como la aptitud evolutiva de un organismo (darwiniano).
La aptitud física suele ser cuantificable y es medida por científicos en el campo. Sin embargo, no es la aptitud absoluta de un individuo lo que cuenta, sino cómo se compara con los otros organismos de la población. Este concepto, llamado aptitud relativa, permite a los investigadores determinar qué individuos están aportando descendencia adicional a la siguiente generación, y así, cómo podría evolucionar la población.
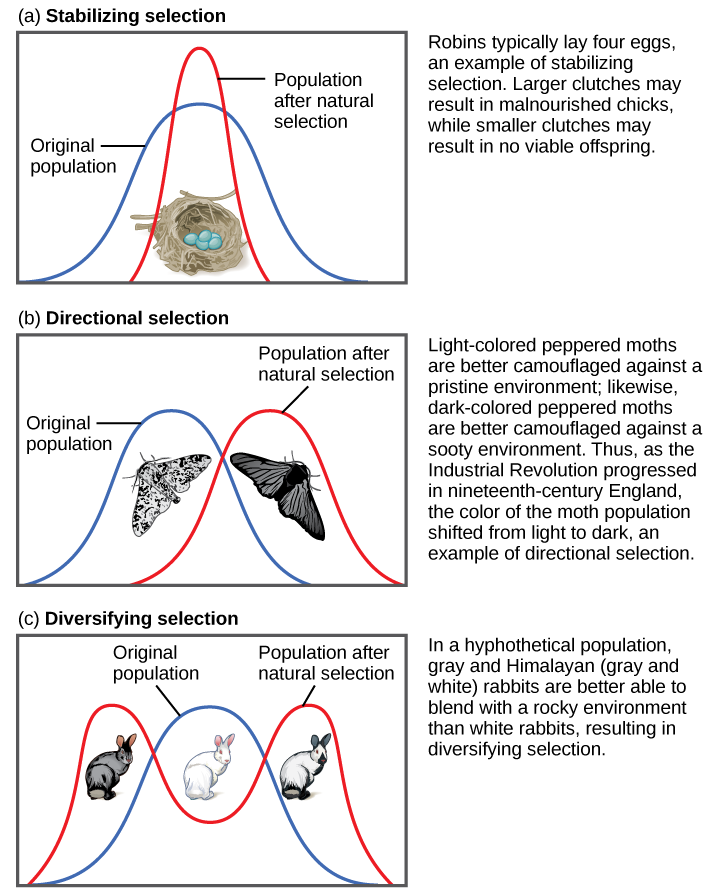
Hay varias formas en que la selección puede afectar la variación poblacional: la selección estabilizadora, la selección direccional, la selección diversificadora, la selección dependiente de la frecuencia y la selección sexual. A medida que la selección natural influye en las frecuencias alélicas en una población, los individuos pueden volverse más o menos genéticamente similares y los fenotipos mostrados pueden volverse más similares o más dispares.
Selección estabilizadora
Si la selección natural favorece un fenotipo promedio, seleccionando contra variación extrema, la población se someterá a selección estabilizadora (Figura\(\PageIndex{1}\)). En una población de ratones que viven en el bosque, por ejemplo, es probable que la selección natural favorezca a los individuos que mejor se mezclan con el suelo del bosque y tienen menos probabilidades de ser vistos por depredadores. Asumiendo que el suelo es un tono marrón bastante consistente, aquellos ratones cuyo pelaje se empareja más a ese color tendrán más probabilidades de sobrevivir y reproducirse, transmitiendo sus genes por su pelaje marrón. Los ratones que portan alelos que los hacen un poco más claros o un poco más oscuros destacarán contra el suelo y serán más propensos a ser víctimas de la depredación. Como resultado de esta selección, la varianza genética de la población disminuirá.
Selección Direccional
Cuando el ambiente cambia, las poblaciones a menudo se someterán a selección direccional (Figura\(\PageIndex{1}\)), que selecciona fenotipos en un extremo del espectro de variación existente. Un ejemplo clásico de este tipo de selección es la evolución de la polilla salpimentada en la Inglaterra de los siglos XVIII y XIX. Antes de la Revolución Industrial, las polillas eran predominantemente de color claro, lo que les permitió mezclarse con los árboles y líquenes de color claro en su entorno. Pero a medida que el hollín comenzó a arrojar de las fábricas, los árboles se oscurecieron y las polillas de color claro se volvieron más fáciles de detectar para las aves depredadoras. Con el tiempo, la frecuencia de la forma melánica de la polilla aumentó debido a que tuvieron una mayor tasa de supervivencia en hábitats afectados por la contaminación del aire debido a que su coloración más oscura se mezcló con los árboles hollín. De igual manera, la hipotética población de ratones puede evolucionar para adquirir una coloración diferente si algo provocara que el suelo del bosque donde viven cambie de color. El resultado de este tipo de selección es un cambio en la varianza genética de la población hacia el nuevo fenotipo fit.
Diversificar la selección
En ocasiones, dos o más fenotipos distintos pueden tener cada uno sus ventajas y ser seleccionados por selección natural, mientras que los fenotipos intermedios son, en promedio, menos aptos. Conocida como selección diversificadora (Figura\(\PageIndex{1}\)), esto se observa en muchas poblaciones de animales que tienen múltiples formas masculinas. Los machos alfa grandes y dominantes obtienen parejas por fuerza bruta, mientras que los machos pequeños pueden colarse para copulaciones furtivas con las hembras en el territorio de un macho alfa. En este caso, tanto los machos alfa como los machos “furtivos” serán seleccionados para, pero los machos medianos, que no pueden superar a los machos alfa y son demasiado grandes para escabullirse cópulaciones, son seleccionados en contra. La diversificación de la selección también puede ocurrir cuando los cambios ambientales favorecen a los individuos en ambos extremos del espectro fenotípico. Imagina una población de ratones que viven en la playa donde hay arena de color claro intercalada con parches de pasto alto. En este escenario, se favorecería a los ratones de color claro que se mezclan con la arena, así como a los ratones de color oscuro que pueden esconderse en la hierba. Los ratones de color medio, por otro lado, no se mezclarían ni con la hierba ni con la arena, y por lo tanto serían más propensos a ser comidos por depredadores. El resultado de este tipo de selección es el aumento de la varianza genética a medida que la población se vuelve más diversa.
Ejercicio\(\PageIndex{1}\)
En los últimos años, las fábricas se han vuelto más limpias y se libera menos hollín al medio ambiente. ¿Qué impacto cree que esto ha tenido en la distribución del color de la polilla en la población?
- Responder
-
Las polillas han cambiado a un color más claro.
Selección dependiente de la frecuencia
Otro tipo de selección, llamada selección dependiente de la frecuencia, favorece fenotipos que son comunes (selección positiva dependiente de la frecuencia) o raros (selección dependiente de la frecuencia negativa). Un ejemplo interesante de este tipo de selección se observa en un grupo único de lagartos del noroeste del Pacífico. Los lagartos machos con manchas laterales comunes vienen en tres patrones de color garganta: naranja, azul y amarillo. Cada una de estas formas tiene una estrategia reproductiva diferente: los machos anaranjados son los más fuertes y pueden luchar contra otros machos para acceder a sus hembras; los machos azules son de tamaño mediano y forman fuertes lazos de pareja con sus parejas; y los machos amarillos (Figura\(\PageIndex{2}\)) son los más pequeños, y se parecen un poco a las hembras, que les permite colarse cópulaciones. Al igual que un juego de piedra-papel-tijeras, el naranja supera al azul, el azul late al amarillo y el amarillo le gana al naranja en la competencia por las hembras. Es decir, los machos anaranjados grandes y fuertes pueden luchar contra los machos azules para aparearse con las hembras unidas por parejas del azul, los machos azules tienen éxito en proteger a sus compañeros contra los machos de zapatilla amarilla, y los machos amarillos pueden escabullirse cópulaciones de los posibles compañeros de los machos anaranjados grandes y políginos.

En este escenario, los machos anaranjados se verán favorecidos por la selección natural cuando la población esté dominada por machos azules, los machos azules prosperarán cuando la población sea mayoritariamente machos amarillos, y los machos amarillos serán seleccionados para cuando los machos anaranjados sean los más poblados. Como resultado, las poblaciones de lagartos con manchas laterales circulan en la distribución de estos fenotipos; en una generación, el naranja podría ser predominante, y luego los machos amarillos comenzarán a aumentar en frecuencia. Una vez que los machos amarillos constituyen la mayoría de la población, los machos azules serán seleccionados para. Por último, cuando los machos azules se vuelvan comunes, los machos anaranjados volverán a ser favorecidos.
La selección negativa dependiente de la frecuencia sirve para aumentar la varianza genética de la población mediante la selección de fenotipos raros, mientras que la selección positiva dependiente de la frecuencia generalmente disminuye la varianza genética al seleccionar fenotipos comunes.
Selección Sexual
Los machos y hembras de ciertas especies suelen ser bastante diferentes entre sí en formas más allá de los órganos reproductivos. Los machos suelen ser más grandes, por ejemplo, y muestran muchos colores y adornos elaborados, como la cola del pavo real, mientras que las hembras tienden a ser más pequeñas y más opacas en la decoración. Tales diferencias se conocen como dimorfismos sexuales (Figura\(\PageIndex{3}\)), que surgen del hecho de que en muchas poblaciones, particularmente las poblaciones animales, hay más varianza en el éxito reproductivo de los machos que en las hembras. Es decir, algunos machos —a menudo los machos más grandes, más fuertes o más decorados— obtienen la gran mayoría de los apareamientos totales, mientras que otros no reciben ninguno. Esto puede ocurrir porque los machos son mejores para luchar contra otros machos, o porque las hembras elegirán aparearse con los machos más grandes o más condecorados. En cualquier caso, esta variación en el éxito reproductivo genera una fuerte presión de selección entre los machos para obtener esos apareamientos, lo que resulta en la evolución de un mayor tamaño corporal y elaborados ornamentos para llamar la atención de las mujeres. Las hembras, por otro lado, tienden a obtener un puñado de apareamientos seleccionados; por lo tanto, es más probable que seleccionen machos más deseables.
El dimorfismo sexual varía ampliamente entre las especies, por supuesto, y algunas especies incluso tienen un rol sexual invertido. En tales casos, las hembras tienden a tener una mayor varianza en su éxito reproductivo que los machos y se seleccionan correspondientemente por el tamaño corporal más grande y rasgos elaborados generalmente característicos de los machos.

Las presiones de selección sobre machos y hembras para obtener apareamientos se conocen como selección sexual; puede resultar en el desarrollo de características sexuales secundarias que no benefician la probabilidad de supervivencia del individuo sino que ayudan a maximizar su éxito reproductivo. La selección sexual puede ser tan fuerte que selecciona por rasgos que en realidad son perjudiciales para la supervivencia del individuo. Piensa, una vez más, en la cola del pavo real. Si bien es hermoso y el macho con la cola más grande y colorida tiene más probabilidades de ganar a la hembra, no es el apéndice más práctico. Además de ser más visible para los depredadores, hace que los machos sean más lentos en sus intentos de fuga. Hay alguna evidencia de que este riesgo, de hecho, es la razón por la que a las hembras les gustan las colas grandes en primer lugar. La especulación es que las colas grandes conllevan riesgo, y solo los mejores machos sobreviven a ese riesgo: cuanto más grande es la cola, más se ajusta el macho. Esta idea se conoce como el principio de handicap.
La hipótesis de los buenos genes establece que los machos desarrollan estos impresionantes ornamentos para mostrar su metabolismo eficiente o su capacidad para combatir enfermedades. Luego, las hembras eligen machos con los rasgos más impresionantes porque señala su superioridad genética, que luego pasarán a sus crías. Aunque se podría argumentar que las hembras no deben ser exigentes porque probablemente reducirá su número de crías, si mejores machos engendran descendencia más en forma, puede ser beneficioso. Menos crías más sanas pueden aumentar las posibilidades de supervivencia más que muchas crías más débiles. En 1915, el biólogo Ronald Fisher propuso otro modelo de selección sexual: el modelo fugado fisheriano, que sugiere que la selección de ciertos rasgos es resultado de la preferencia sexual.
Tanto en el principio de hándicap como en la hipótesis de los buenos genes, se dice que el rasgo es una señal honesta de la calidad de los machos, dando así a las hembras una manera de encontrar a los compañeros más aptos, machos que pasarán los mejores genes a sus crías.
Ningún organismo perfecto
La selección natural es una fuerza impulsora en la evolución y puede generar poblaciones que están mejor adaptadas para sobrevivir y reproducirse con éxito en sus entornos. Pero la selección natural no puede producir el organismo perfecto. La selección natural solo puede seleccionar sobre la variación existente en la población; no crea nada desde cero. Por lo tanto, está limitado por la varianza genética existente de una población y cualquier alelo nuevo que surja a través de la mutación y el flujo génico.
La selección natural también es limitada porque funciona a nivel de individuos, no alelos, y algunos alelos están vinculados debido a su proximidad física en el genoma, haciéndolos más propensos a transmitirse juntos (desequilibrio de ligamiento). Cualquier individuo dado puede portar algunos alelos beneficiosos y algunos alelos desfavorables. Es el efecto neto de estos alelos, o la aptitud del organismo, sobre el que puede actuar la selección natural. Como resultado, los alelos buenos se pueden perder si son portados por individuos que también tienen varios alelos abrumadoramente malos; de igual manera, los alelos malos se pueden mantener si son portados por individuos que tienen suficientes alelos buenos para resultar en un beneficio general de aptitud física.
Además, la selección natural puede estar limitada por las relaciones entre diferentes polimorfismos. Una morph puede conferir una mayor aptitud que otra, pero puede que no aumente de frecuencia debido a que pasar del rasgo menos beneficioso al más beneficioso requeriría pasar por un fenotipo menos beneficioso. Piensa en los ratones que viven en la playa. Algunos son de color claro y se mezclan con la arena, mientras que otros son oscuros y se mezclan con los parches de hierba. Los ratones de color oscuro pueden estar, en general, más en forma que los ratones de color claro, y a primera vista, uno podría esperar que los ratones de color claro sean seleccionados para una coloración más oscura. Pero recuerda que el fenotipo intermedio, un pelaje de color medio, es muy malo para los ratones, no pueden mezclarse ni con la arena ni con la hierba y es más probable que los depredadores los coman. Como resultado, los ratones de color claro no serían seleccionados para una coloración oscura debido a que aquellos individuos que comenzaron a moverse en esa dirección (comenzaron a ser seleccionados para un pelaje más oscuro) estarían menos en forma que aquellos que permanecieron claros.
Por último, es importante entender que no toda la evolución es adaptativa. Si bien la selección natural selecciona a los individuos más aptos y a menudo da como resultado una población más adecuada en general, otras fuerzas de evolución, incluyendo la deriva genética y el flujo génico, a menudo hacen lo contrario: introducir alelos deletéreos al acervo genético de la población. La evolución no tiene propósito, no es transformar una población en un ideal preconcebido. Es simplemente la suma de las diversas fuerzas descritas en este capítulo y cómo influyen en la varianza genética y fenotípica de una población.
Resumen
Debido a que la selección natural actúa para aumentar la frecuencia de alelos y rasgos beneficiosos mientras disminuye la frecuencia de cualidades deletéreas, es evolución adaptativa. La selección natural actúa a nivel del individuo, seleccionando para aquellos que tienen una mayor aptitud general en comparación con el resto de la población. Si los fenotipos de ajuste son los que son similares, la selección natural dará como resultado una selección estabilizadora y una disminución general en la variación de la población. La selección direccional trabaja para cambiar la varianza de una población hacia un nuevo fenotipo de ajuste, a medida que cambian las condiciones ambientales. Por el contrario, la diversificación de la selección da como resultado un aumento de la varianza genética al seleccionar dos o más fenotipos distintos.
Otros tipos de selección incluyen la selección dependiente de la frecuencia, en la que se seleccionan individuos con común (selección dependiente de la frecuencia positiva) o raros (selección dependiente de la frecuencia negativa) para. Finalmente, la selección sexual resulta del hecho de que un sexo tiene más varianza en el éxito reproductivo que el otro. Como resultado, machos y hembras experimentan diferentes presiones selectivas, lo que a menudo puede conducir a la evolución de diferencias fenotípicas, o dimorfismos sexuales, entre los dos.
Referencias
OpenStax, Biología. OpenStax CNX. 25 de junio de 2020. https://cnx.org/contents/GFy_h8cu@10.137:noBcfThl@7/Understanding-Evolution.