
25.2: Dinámica y Regulación de la Población
- Page ID
- 58250
El modelo logístico de crecimiento poblacional, si bien es válido en muchas poblaciones naturales y un modelo útil, es una simplificación de la dinámica poblacional del mundo real. Implícito en el modelo es que la capacidad de carga del entorno no cambia, lo que no es el caso. La capacidad de carga varía anualmente: por ejemplo, algunos veranos son calurosos y secos mientras que otros son fríos y húmedos. En muchas zonas, la capacidad de carga durante el invierno es mucho menor que durante el verano. Además, eventos naturales como sismos, volcanes e incendios pueden alterar un ambiente y de ahí su capacidad de carga. Adicionalmente, las poblaciones no suelen existir en aislamiento. Se dedican a una competencia interespecífica: es decir, comparten el ambiente con otras especies, compitiendo con ellas por los mismos recursos. Estos factores también son importantes para entender cómo crecerá una población específica.
La naturaleza regula el crecimiento de la población de diversas maneras. Estos se agrupan en factores dependientes de la densidad, en los que la densidad de la población en un momento dado afecta la tasa de crecimiento y mortalidad, y factores independientes de la densidad, que influyen en la mortalidad en una población independientemente de la densidad poblacional. Obsérvese que en la primera, el efecto del factor en la población depende de la densidad de la población al inicio. Los biólogos de la conservación quieren entender ambos tipos porque esto les ayuda a manejar poblaciones y prevenir la extinción o sobrepoblación.
Regulación dependiente de la densidad
La mayoría de los factores dependientes de la densidad son de naturaleza biológica (bióticos) e incluyen depredación, competencia inter e intraespecífica, acumulación de desechos y enfermedades como las causadas por parásitos. Por lo general, cuanto más densa es una población, mayor es su tasa de mortalidad. Por ejemplo, durante la competencia intra e interespecífica, las tasas reproductivas de los individuos suelen ser menores, reduciendo la tasa de crecimiento de su población. Además, la baja densidad de presas aumenta la mortalidad de su depredador debido a que tiene más dificultades para localizar su fuente de alimento.
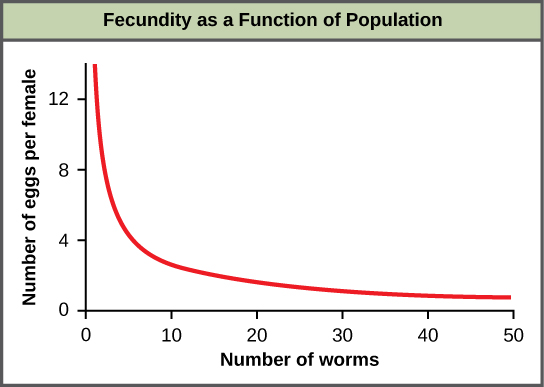
Un ejemplo de regulación dependiente de la densidad se muestra en la Figura\(\PageIndex{1}\) con resultados de un estudio centrado en la lombriz intestinal gigante (Ascaris lumbricoides), un parásito de humanos y otros mamíferos (Croll et al. 1982). Las poblaciones más densas del parásito presentaron menor fecundidad: contenían menos huevos. Una posible explicación para esto es que las hembras serían más pequeñas en poblaciones más densas (debido a los recursos limitados) y que las hembras más pequeñas tendrían menos huevos. Esta hipótesis fue probada y desmentida en un estudio de 2009 que demostró que el peso femenino no tuvo influencia (Walker et al, 2009). La causa real de la densidad-dependencia de la fecundidad en este organismo aún no está clara y a la espera de una mayor investigación.
Regulación independiente de la densidad e interacción con factores dependientes de la densidad
La mayoría de los factores independientes de la densidad son de naturaleza física o química (abióticos) Estos factores influyen en la mortalidad de una población independientemente de su densidad, incluyendo el clima, los desastres naturales y la contaminación. Un ciervo individual puede morir en un incendio forestal independientemente de cuántos venados se encuentren en esa zona. Sus posibilidades de supervivencia son las mismas tanto si la densidad poblacional es alta como baja. Lo mismo ocurre con el clima frío de invierno.
En situaciones de la vida real, la regulación poblacional es muy complicada y los factores dependientes de la densidad e independientes pueden interactuar. Una población densa que se reduce de manera independiente de la densidad por algún factor (s) ambiental (s) podrá recuperarse de manera diferente a una población dispersa. Por ejemplo, una población de venados afectados por un duro invierno se recuperará más rápido si quedan más venados por reproducirse.
Evolution Connection — ¿Por qué se extinguió el mamut lanudo?
Es fácil perderse en la discusión de dinosaurios y teorías sobre por qué se extinguieron hace 65 millones de años. ¿Se debió a que un meteorito se estrellaba contra la Tierra cerca de la costa del México moderno, o era de algún ciclo meteorológico de largo plazo que aún no se entiende? Una hipótesis que nunca se propondrá es que los humanos tuvieron algo que ver con ello. Los mamíferos eran criaturas pequeñas e insignificantes del bosque hace 65 millones de años, y no existían humanos.



Los mamuts lanudos, sin embargo, comenzaron a extinguirse hace unos 10 mil años, cuando compartieron la Tierra con humanos que no eran diferentes anatómicamente a los humanos de hoy (Figura\(\PageIndex{2}\)). Los mamuts sobrevivieron en poblaciones aisladas de islas tan recientemente como 1700 a.C. Sabemos mucho de estos animales a partir de cadáveres encontrados congelados en el hielo de Siberia y otras regiones del norte. Los científicos han secuenciado al menos el 50 por ciento de su genoma y creen que los mamuts son entre 98 y 99 por ciento idénticos a los elefantes modernos.
Comúnmente se piensa que el cambio climático y la caza humana llevaron a su extinción. Un estudio de 2008 estimó que el cambio climático redujo el rango del mamut de 3.000.000 de millas cuadradas hace 42,000 años a 310,000 millas cuadradas hace 6,000 años (Nogués-Bravo et al. 2008). También está bien documentado que los humanos cazaban a estos animales. Un estudio de 2012 mostró que ningún factor fue el responsable exclusivo de la extinción de estas magníficas criaturas (MacDonald et al. 2012). Además de la caza humana, el cambio climático y la reducción del hábitat, estos científicos demostraron que otro factor importante en la extinción del mamut fue la migración de humanos a través del Estrecho de Bering a América del Norte durante la última edad de hielo hace 20 mil años.
El mantenimiento de poblaciones estables fue y es muy complejo, con muchos factores que interactúan determinando el desenlace. Es importante recordar que los humanos también son parte de la naturaleza. Una vez contribuimos al declive de una especie utilizando únicamente tecnología de caza primitiva.
Historias de vida de especies seleccionadas por K y seleccionadas por r
Si bien las estrategias reproductivas juegan un papel clave en las historias de vida, no tienen en cuenta factores importantes como los recursos limitados y la competencia. La regulación del crecimiento poblacional por estos factores puede ser utilizada para introducir un concepto clásico en biología poblacional, el de especies seleccionadas por K versus especies seleccionadas por r.
Teorías tempranas sobre la historia de vida: Especie K -seleccionada y r -seleccionada
Para la segunda mitad del siglo XX, el concepto de especies seleccionadas K y r se utilizó ampliamente y con éxito para estudiar poblaciones. El concepto se relaciona no solo con estrategias reproductivas sino también con el hábitat y comportamiento de una especie, especialmente en la forma en que obtiene recursos y cuidado de sus crías. Incluye también la duración de la vida y los factores de supervivencia. Para este análisis, los biólogos poblacionales han agrupado especies en las dos grandes categorías, seleccionadas por K y seleccionadas por r, aunque en realidad son dos extremos de un continuo.
Las especies seleccionadas por K son especies seleccionadas por ambientes estables y predecibles. Las poblaciones de especies seleccionadas por K tienden a existir cerca de su capacidad de carga (de ahí el término K -selected) donde la competencia intraespecífica es alta. Estas especies tienen pocas crías grandes, un largo período de gestación, y a menudo dan cuidados a largo plazo a sus crías (Tabla\(\PageIndex{1}\)). Si bien son más grandes en tamaño al nacer, las crías son relativamente indefensas e inmaduras al nacer. Para cuando lleguen a la edad adulta, deben desarrollar habilidades para competir por los recursos naturales. En las plantas, los científicos piensan en el cuidado de los padres de manera más amplia: cuánto tiempo tarda en desarrollarse el fruto o cuánto tiempo permanece en la planta son factores determinantes en el tiempo hasta el siguiente evento reproductivo. Ejemplos de especies seleccionadas por K son primates (incluyendo humanos), elefantes y plantas como los robles (Figura\(\PageIndex{3}\) a).
Los encinos crecen muy lentamente y tardan, en promedio, 20 años en producir sus primeras semillas, conocidas como bellotas. Hasta 50,000 bellotas pueden ser producidas por un árbol individual, pero la tasa de germinación es baja ya que muchas de estas se pudren o son consumidas por animales como las ardillas. En algunos años, los encinos pueden producir un número excepcionalmente grande de bellotas, y estos años pueden estar en un ciclo de dos o tres años dependiendo de la especie de encino (r -selección).
A medida que los encinos crecen a un gran tamaño y durante muchos años antes de que comiencen a producir bellotas, dedican un gran porcentaje de su presupuesto energético al crecimiento y mantenimiento. La altura y el tamaño del árbol le permiten dominar otras plantas en la competencia por la luz solar, el principal recurso energético del roble. Además, cuando se reproduce, el roble produce semillas grandes y ricas en energía que utilizan su reserva energética para establecerse rápidamente (selección K).
En contraste, las especies seleccionadas por r tienen un gran número de crías pequeñas (de ahí su designación r (Tabla\(\PageIndex{1}\))). Esta estrategia suele emplearse en entornos impredecibles o cambiantes. Los animales seleccionados por r no dan cuidados parentales a largo plazo y las crías son relativamente maduras y autosuficientes al nacer. Ejemplos de especies seleccionadas por r son invertebrados marinos, como medusas, y plantas, como el diente de león (Figura\(\PageIndex{3}\) b). Los dientes de león tienen semillas pequeñas que se dispersan por el viento a largas distancias. Muchas semillas se producen simultáneamente para asegurar que al menos algunas de ellas lleguen a un ambiente hospitalario. Las semillas que aterrizan en ambientes inhóspitos tienen pocas posibilidades de supervivencia ya que sus semillas son bajas en contenido energético. Tenga en cuenta que la supervivencia no es necesariamente una función de la energía almacenada en la propia semilla.
| Tabla\(\PageIndex{1}\): Características de las especies seleccionadas por K y las especies seleccionadas por r | |
|---|---|
| Características de las especies seleccionadas por K | Características de las especies seleccionadas por r |
| \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas">Maduras tardías | \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas">Maduras tempranas |
| \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas K-">Mayor longevidad | \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas por r">Menor longevidad |
| \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas K-">Aumento del cuidado parental | \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas">Disminución del cuidado parental |
| \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas K-">Mayor competencia | \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas">Disminución de la competencia |
| \ (\ PageIndex {1}\): Características de las especies seleccionadas por K y lascaracterísticas de las especies seleccionadas">Menos crías | \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas">Más crías |
| \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas">Cría mayor | \ (\ PageIndex {1}\): Características de las especies seleccionadas K y r-seleccionadasCaracterísticas de las especies seleccionadas por r">descendencia más pequeña |

Teorías modernas de la historia de la vida
La teoría de la selección r y K, aunque aceptada durante décadas y utilizada para muchas investigaciones innovadoras, ahora ha sido reconsiderada, y muchos biólogos de población la han abandonado o modificado. A lo largo de los años, varios estudios intentaron confirmar la teoría, pero estos intentos han fracasado en gran medida. Se identificaron muchas especies que no siguieron las predicciones de la teoría. Además, la teoría ignoró la mortalidad específica por edad de las poblaciones que los científicos ahora saben que es muy importante. Se han desarrollado nuevos modelos demográficos de evolución de la historia de vida que incorporan muchos conceptos ecológicos incluidos en la teoría de la selección r y K, así como la estructura de edad de la población y los factores de mortalidad.
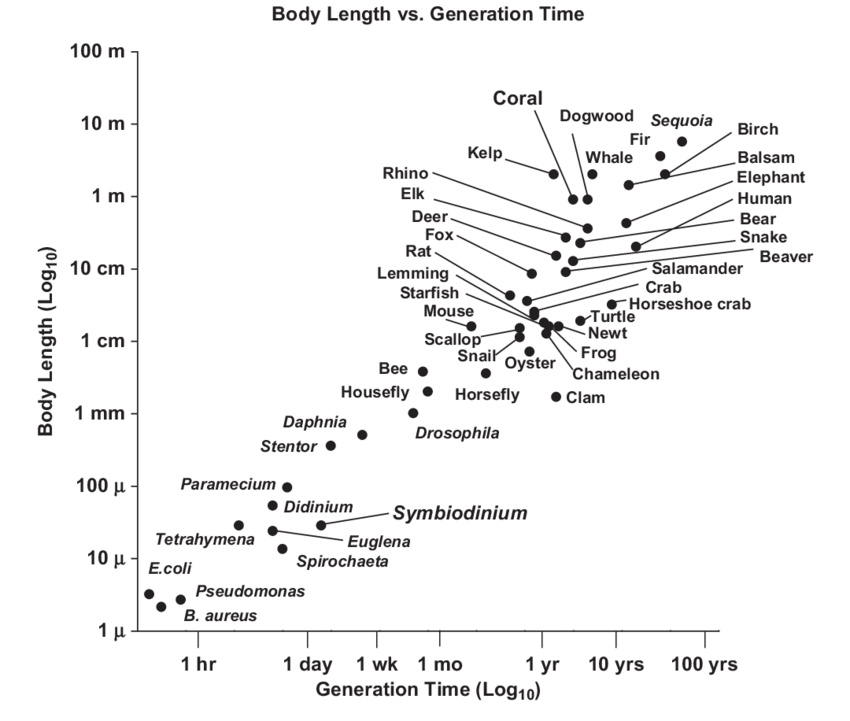
El tamaño corporal se correlaciona con el tiempo de generación
El tiempo de generación es el lapso promedio de tiempo entre el nacimiento de un individuo y el nacimiento de su descendencia. La tendencia general es que cuanto mayor sea la especie, mayor será el tiempo de generación (Figura\(\PageIndex{4}\)).
Resumen
Las poblaciones están reguladas por una variedad de factores dependientes de la densidad e independientes de la densidad. Las especies se dividen en dos categorías en función de una variedad de características de sus patrones de historia de vida: especies seleccionadas por r, que tienen un gran número de crías, y especies seleccionadas por K, que tienen pocas crías. La teoría de la selección r y K ha caído fuera de uso; sin embargo, muchas de sus características clave todavía se utilizan en modelos más nuevos de dinámica demográfica de base demográfica.
Referencias
Croll, N.A. et al. 1982. “La biología poblacional y el control de Ascaris lumbricoides en una comunidad rural en Irán”. Transacciones de la Real Sociedad de Medicina e Higiene Tropical 76, núm. 2:187-197, doi:10.1016/0035-9203 (82) 90272-3.
MacDonald, G.M., et al. 2012. “Patrón de extinción del mamut lanudo en Beringia”. Naturaleza Comunicaciones 3, núm. 893, doi:10.1038/ncomms1881.
Nogués-Bravo, D. et al. 2008. “El cambio climático, los humanos y la extinción del mamut lanudo”. PLoS Biol 6: e79, doi:10.1371/journal.pbio.0060079.
OpenStax, Biología. OpenStax CNX. 26 de junio de 2020. https://cnx.org/contents/GFy_h8cu@10.137:noBcfThl@7/Understanding-Evolution.
Walker, M. et al. 2009. “Efectos dependientes de la densidad sobre el peso de las infecciones femeninas de Ascaris lumbricoides en humanos y su impacto en los patrones de producción de huevos”. Parásitos y vectores 2, núm. 11, doi:10.1186/1756-3305-2-11.
Sammarco, P., & Strychar, K. 2009. Efectos del Cambio Climático/Calentamiento Global en Arrecifes de Coral: Adaptación/Exaptación en Corales, Evolución en Zooxantelas y Cambios Biogeográficos. Bioindicadores Ambientales, 4, 9—45. https://doi.org/10.1080/15555270902905377