
25.3: Historias de vida y selección natural
- Page ID
- 58245
La historia de vida de una especie describe la serie de eventos a lo largo de su vida, como la forma en que se asignan los recursos para el crecimiento, el mantenimiento y la reproducción. Los rasgos de la historia de vida afectan la tabla de vida de un organismo. La historia de vida de una especie está determinada genéticamente y conformada por el entorno y la selección natural.
Patrones de historia de vida y presupuestos energéticos
La energía es requerida por todos los organismos vivos para su crecimiento, mantenimiento y reproducción; al mismo tiempo, la energía es a menudo un factor limitante importante para determinar la supervivencia de un organismo. Las plantas, por ejemplo, adquieren energía del sol a través de la fotosíntesis pero deben gastar esta energía para crecer, mantener la salud y producir semillas ricas en energía para producir la próxima generación. Los animales tienen la carga adicional de usar algunas de sus reservas de energía para adquirir alimentos. Además, algunos animales deben gastar energía cuidando a sus crías. Así, todas las especies tienen un presupuesto energético: deben equilibrar la ingesta de energía con su uso de energía para el metabolismo, la reproducción, el cuidado parental y el almacenamiento de energía (como los osos que construyen grasa corporal para la hibernación invernal).
Cuidado parental y fecundidad
La fecundidad es la capacidad reproductiva potencial de un individuo dentro de una población. La fecundidad describe cuántas crías idealmente podrían producirse si un individuo tiene tantas crías como sea posible, repitiendo el ciclo reproductivo lo antes posible después del nacimiento de la descendencia. En los animales, la fecundidad está inversamente relacionada con la cantidad de cuidado parental que se da a una descendencia individual. Las especies, como muchos invertebrados marinos, que producen muchas crías suelen proporcionar poco o ningún cuidado para la descendencia (de todos modos no tendrían la energía ni la capacidad de hacerlo). La mayor parte de su presupuesto energético se utiliza para producir muchas crías diminutas. Los animales con esta estrategia suelen ser autosuficientes a una edad muy temprana. Esto se debe a la compensación energética que estos organismos han hecho para maximizar su aptitud evolutiva. Debido a que su energía se utiliza para producir descendencia en lugar del cuidado de los padres, tiene sentido que estas crías tengan alguna capacidad para poder moverse dentro de su entorno y encontrar alimento y tal vez refugio. Incluso con estas habilidades, su pequeño tamaño las hace extremadamente vulnerables a la depredación, por lo que la producción de muchas crías permite que suficientes de ellas sobrevivan para mantener a la especie.
Las especies animales que tienen pocas crías durante un evento reproductivo suelen dar un amplio cuidado parental, dedicando gran parte de su presupuesto energético a estas actividades, a veces a expensas de su propia salud. Este es el caso de muchos mamíferos, como humanos, canguros y pandas. Las crías de estas especies están relativamente indefensas al nacer y necesitan desarrollarse antes de alcanzar la autosuficiencia.
Las plantas con baja fecundidad producen pocas semillas ricas en energía (como los cocos y las castañas) y cada una tiene una buena oportunidad de germinar en un nuevo organismo; las plantas con alta fecundidad suelen tener muchas semillas pequeñas y pobres en energía (como las orquídeas) que tienen una probabilidad relativamente baja de sobrevivir. Si bien puede parecer que los cocos y las castañas tienen más posibilidades de sobrevivir, la compensación energética de la orquídea también es muy efectiva. Se trata de dónde se utiliza la energía, para grandes cantidades de semillas o para menos semillas con más energía.
Reproducción temprana versus tardía
El momento de la reproducción en una historia de vida también afecta la supervivencia de las especies. Los organismos que se reproducen a temprana edad tienen una mayor probabilidad de producir descendencia, pero esto suele ser a expensas de su crecimiento y del mantenimiento de su salud. Por el contrario, los organismos que comienzan a reproducirse más tarde en la vida suelen tener mayor fecundidad o están en mejores condiciones de brindar atención parental, pero corren el riesgo de que no sobrevivan hasta la edad reproductiva. Ejemplos de esto se pueden ver en peces. Los peces pequeños como los guppies utilizan su energía para reproducirse rápidamente, pero nunca alcanzan el tamaño que les daría defensa contra algunos depredadores. Los peces más grandes, como el agallas azules o el tiburón, utilizan su energía para alcanzar un tamaño grande pero lo hacen con el riesgo de que mueran antes de poder reproducirse o al menos reproducirse al máximo. Estas diferentes estrategias energéticas y compensaciones son clave para entender la evolución de cada especie ya que maximiza su aptitud y llena su nicho. En términos de presupuestación energética, algunas especies “lo arruinan todo” y agotan la mayor parte de sus reservas de energía para reproducirse temprano antes de morir. Otras especies retrasan tener reproducción para convertirse en individuos más fuertes, más experimentados y para asegurarse de que son lo suficientemente fuertes como para brindar atención parental si es necesario.
Eventos Reproductivos Únicos versus Múltiples
Algunos rasgos de la historia de vida, como la fecundidad, el momento de la reproducción y el cuidado parental, se pueden agrupar en estrategias generales que son utilizadas por múltiples especies. La semelparidad ocurre cuando una especie se reproduce solo una vez durante su vida y luego muere. Dichas especies utilizan la mayor parte de su presupuesto de recursos durante un solo evento reproductivo, sacrificando su salud hasta el punto de que no sobreviven. Ejemplos de semelparidad son el bambú, que florece de vez en cuando muere, y el salmón Chinook (Figura\(\PageIndex{1}\)), que utiliza la mayor parte de sus reservas de energía para migrar del océano a su área de anidación de agua dulce, donde se reproduce y luego muere. Los científicos han postulado explicaciones alternativas para la ventaja evolutiva de la muerte post-reproducción del Chinook: un suicidio programado causado por una liberación masiva de hormonas corticosteroides, presumiblemente por lo que los padres pueden convertirse en alimento para la descendencia, o simple agotamiento causado por las demandas de energía de reproducción; estos aún se están debatiendo.
Iteroparity describe especies que se reproducen repetidamente durante sus vidas. Algunos animales solo pueden aparearse una vez al año pero sobreviven a múltiples temporadas de apareamiento. El antílope pronghorn es un ejemplo de un animal que entra en un ciclo estacional del estro (“calor”): una condición fisiológica inducida por hormonas que prepara al cuerpo para el apareamiento exitoso (Figura\(\PageIndex{1}\)). Las hembras de estas especies se aparean solo durante la fase estro del ciclo. Se observa un patrón diferente en primates, incluyendo humanos y chimpancés, que pueden intentar reproducirse en cualquier momento durante sus años reproductivos, aunque sus ciclos menstruales hacen que el embarazo sea probable solo unos pocos días al mes durante la ovulación (Figura\(\PageIndex{1}\)).



Conexión Evolutiva — Presupuestos Energéticos, Costos Reproductivos y Selección Sexual en Drosophila
La investigación sobre cómo los animales asignan sus recursos energéticos para el crecimiento, mantenimiento y reproducción ha utilizado una variedad de modelos animales experimentales. Parte de este trabajo se ha realizado utilizando la mosca común de la fruta, Drosophila melanogaster. Los estudios han demostrado que no sólo la reproducción tiene un costo en cuanto a cuánto tiempo viven las moscas macho de la fruta, sino que también las moscas de la fruta que ya se han apareado varias veces tienen espermatozoides limitados que quedan para la reproducción. Las moscas de la fruta maximizan sus últimas posibilidades de reproducción seleccionando parejas óptimas.
En un estudio de 1981, se colocaron moscas macho de la fruta en recintos con hembras vírgenes o inseminadas. Los machos que se aparearon con hembras vírgenes tuvieron períodos de vida más cortos que los que estaban en contacto con el mismo número de hembras inseminadas con las que no pudieron aparearse. Este efecto ocurrió independientemente de cuán grandes (indicativo de su edad) fueran los machos. Así, los machos que no se apareaban vivieron más tiempo, lo que les permitió más oportunidades de encontrar pareja en el futuro.
Estudios más recientes, realizados en 2006, muestran cómo los machos seleccionan a la hembra con la que se aparearán y cómo esto se ve afectado por los apareamientos previos (Figura\(\PageIndex{2}\)) (Byrne & Rice, 2006). A los machos se les permitió seleccionar entre hembras más pequeñas y grandes. Los hallazgos mostraron que las hembras más grandes tuvieron mayor fecundidad, produciendo el doble de crías por apareamiento que las hembras más pequeñas. Los machos que previamente se habían apareado y, por lo tanto, tenían menores suministros de esperma, se denominaban “agotados en recursos”, mientras que los machos que no se habían apareado se denominaban “no agotados en recursos”. El estudio mostró que aunque los machos no agotados en recursos se aparearon preferencialmente con hembras más grandes, esta selección de parejas fue más pronunciada en los machos con escasez de recursos. Así, los machos con suministros de esperma agotados, los cuales fueron limitados en el número de veces que podían aparearse antes de reponer su suministro de esperma, seleccionaron hembras más grandes y fecundadas, maximizando así sus posibilidades de descendencia. Este estudio fue uno de los primeros en demostrar que el estado fisiológico del macho afectó su comportamiento de apareamiento de manera que claramente maximiza su uso de recursos reproductivos limitados.
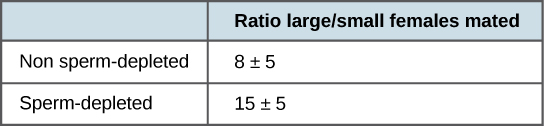
Estos estudios demuestran dos formas en las que el presupuesto energético es un factor de reproducción. Primero, la energía gastada en el apareamiento puede reducir la vida útil de un animal, pero para entonces ya se han reproducido, por lo que en el contexto de la selección natural esta muerte temprana no es de mucha importancia evolutiva. Segundo, cuando los recursos como el esperma (y la energía necesaria para reponerlo) son bajos, el comportamiento de un organismo puede cambiar para darles la mejor oportunidad de pasar sus genes a la siguiente generación. Estos cambios en el comportamiento, tan importantes para la evolución, se estudian en una disciplina conocida como biología conductual, o etología, en la interfaz entre biología poblacional y psicología.
Resumen
Todas las especies han evolucionado un patrón de vida, llamado estrategia de historia de vida, en el que reparten la energía para su crecimiento, mantenimiento y reproducción. Estos patrones evolucionan a través de la selección natural; permiten que las especies se adapten a su entorno para obtener los recursos que necesitan para reproducirse con éxito. Existe una relación inversa entre fecundidad y cuidado parental. Una especie puede reproducirse temprano en la vida para asegurar la supervivencia hasta una edad reproductiva o reproducirse más tarde en la vida para hacerse más grande y saludable y estar en mejores condiciones de brindar el cuidado de los padres. Una especie puede reproducirse una vez (semelparidad) o muchas veces (iteroparidad) en su vida.
Referencias
Byrne, P. y Rice, R. 2006. “Evidencia de elección adaptativa de pareja masculina en la mosca de la fruta Drosophila melanogaster”, Proc Biol Sci. 273, núm. 1589:917-922, doi: 10.1098/rspb.2005.3372.
OpenStax, Biología. OpenStax CNX. 26 de junio de 2020. https://cnx.org/contents/GFy_h8cu@10.137:noBcfThl@7/Understanding-Evolution.