
26.2: Flujo de Energía a través de Ecosistemas
- Page ID
- 58298
Todos los seres vivos requieren energía de una forma u otra. La energía es requerida por las vías metabólicas más complejas (a menudo en forma de trifosfato de adenosina, ATP), especialmente las responsables de construir moléculas grandes a partir de compuestos más pequeños, y la vida misma es un proceso impulsado por la energía. Los organismos vivos no serían capaces de ensamblar macromoléculas (proteínas, lípidos, ácidos nucleicos y carbohidratos complejos) a partir de sus subunidades monoméricas sin un aporte constante de energía.
Es importante entender cómo los organismos adquieren energía y cómo esa energía pasa de un organismo a otro a través de las redes alimentarias y sus cadenas alimentarias constituyentes. Las redes alimentarias ilustran cómo la energía fluye direccionalmente a través de los ecosistemas, incluyendo la eficiencia con la que los organismos la adquieren, la usan y cuánto queda para su uso por otros organismos de la red alimentaria.
Cómo los organismos adquieren energía en una red alimentaria
La energía es adquirida por los seres vivos de tres maneras: fotosíntesis, quimiosíntesis, y el consumo y digestión de otros organismos vivos o previamente vivos por heterótrofos.
Los organismos fotosintéticos y quimiosintéticos se agrupan en una categoría conocida como autótrofos: organismos capaces de sintetizar su propio alimento (más específicamente, capaces de utilizar carbono inorgánico como fuente de carbono). Los autótrofos fotosintéticos (fotoautótrofos) utilizan la luz solar como fuente de energía, mientras que los autótrofos quimiosintéticos (quimioautótrofos) utilizan moléculas inorgánicas como fuente de energía. Los autótrofos son críticos para todos los ecosistemas. Sin estos organismos, la energía no estaría disponible para otros organismos vivos y la vida misma no sería posible.
Los fotoautótrofos, como las plantas, las algas y las bacterias fotosintéticas, sirven como fuente de energía para la mayoría de los ecosistemas del mundo. Estos ecosistemas suelen ser descritos por las redes alimenticias de pastoreo. Los fotoautótrofos aprovechan la energía solar del sol convirtiéndola en energía química en forma de ATP (y NADP). La energía almacenada en ATP se utiliza para sintetizar moléculas orgánicas complejas, como la glucosa.
Los quimioautótrofos son principalmente bacterias que se encuentran en ecosistemas raros donde no se dispone de luz solar, como en aquellos asociados con cuevas oscuras o respiraderos hidrotermales en el fondo del océano (Figura\(\PageIndex{1}\)). Muchos quimioautótrofos en respiraderos hidrotermales utilizan sulfuro de hidrógeno (H 2 S), que se libera de los respiraderos como fuente de energía química. Esto permite a los quimioautótrofos sintetizar moléculas orgánicas complejas, como la glucosa, para su propia energía y a su vez suministra energía al resto del ecosistema.

Productividad dentro de niveles tróficos
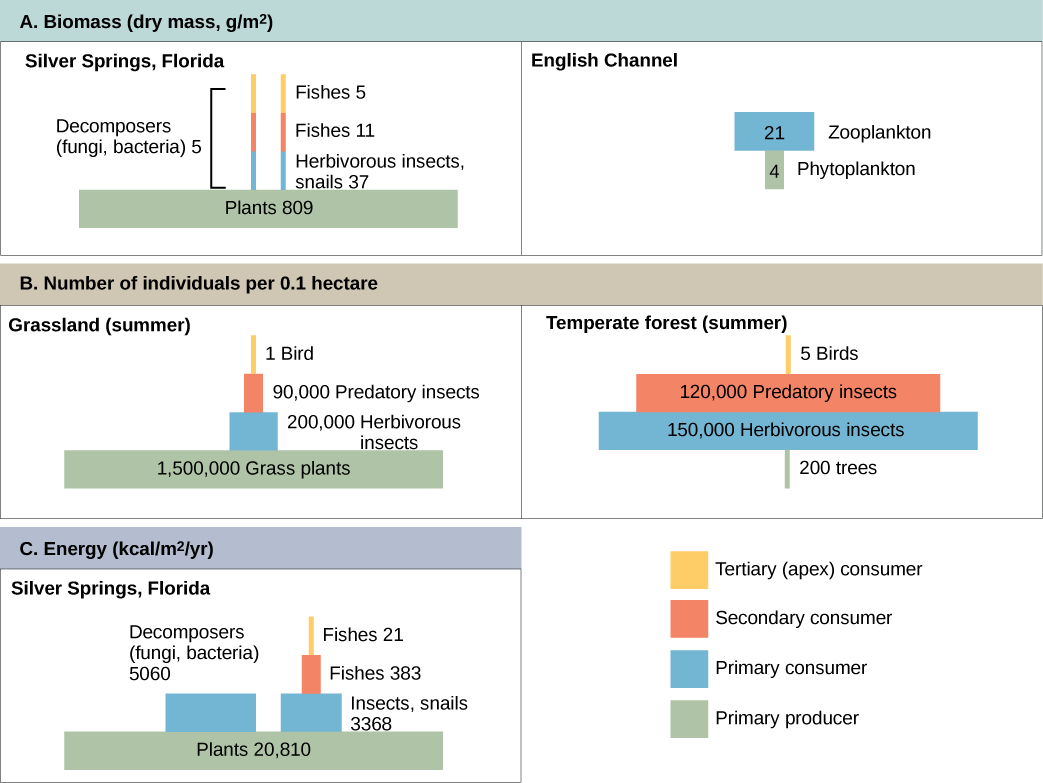
La productividad dentro de un ecosistema puede definirse como el porcentaje de energía que ingresa al ecosistema incorporada a la biomasa en un nivel trófico particular. La biomasa es la masa total, en un área unitaria al momento de la medición, de organismos vivos o previamente vivos dentro de un nivel trófico. Los ecosistemas tienen cantidades características de biomasa en cada nivel trófico. Por ejemplo, en el ecosistema del Canal de la Mancha, los productores primarios representan una biomasa de 4 g/m 2 (gramos por metro cuadrado), mientras que los consumidores primarios exhiben una biomasa de 21 g/m 2.
La productividad de los productores primarios es especialmente importante en cualquier ecosistema porque estos organismos aportan energía a otros organismos vivos por fotoautotrofia o quimioautotrofia. La tasa a la que los productores primarios fotosintéticos incorporan energía del sol se denomina productividad primaria bruta. Un ejemplo de productividad primaria bruta se muestra en el diagrama compartimental del flujo de energía dentro del ecosistema acuático Silver Springs como se muestra (Figura\(\PageIndex{2}\)). En este ecosistema, se demostró que la energía total acumulada por los productores primarios (productividad primaria bruta) era de 20,810 kcal/m 2 /año.
Debido a que todos los organismos necesitan usar parte de esta energía para sus propias funciones (como la respiración y la pérdida de calor metabólica resultante) los científicos a menudo se refieren a la productividad primaria neta de un ecosistema. La productividad primaria neta es la energía que permanece en los productores primarios después de contabilizar la respiración y la pérdida de calor de los organismos. La productividad neta está entonces disponible para los consumidores primarios en el siguiente nivel trófico. En nuestro ejemplo de Silver Spring, 13,187 de los 20,810 kcal/m 2 /año se utilizaron para la respiración o se perdieron como calor, dejando 7,632 kcal/m 2 /año de energía para su uso por los consumidores primarios.
Eficiencia ecológica: La transferencia de energía entre niveles tróficos
Como se ilustra en la Figura\(\PageIndex{2}\), grandes cantidades de energía se pierden del ecosistema de un nivel trófico al siguiente nivel a medida que la energía fluye de los productores primarios a través de los diversos niveles tróficos de consumidores y descomponedores.
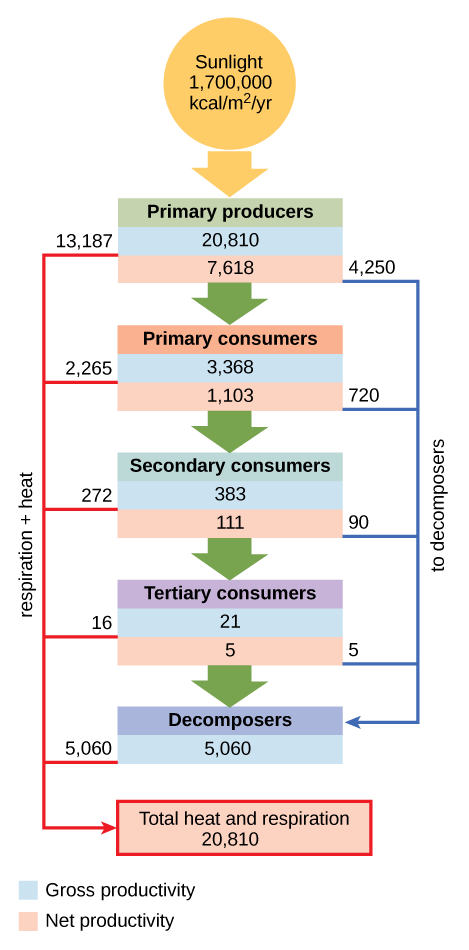
La razón principal de esta pérdida es la segunda ley de la termodinámica, que establece que cada vez que la energía se convierte de una forma a otra, hay una tendencia hacia el desorden (entropía) en el sistema. En los sistemas biológicos, esto significa que una gran cantidad de energía se pierde como calor metabólico cuando los organismos de un nivel trófico consumen el siguiente nivel. En el ejemplo del ecosistema de Silver Springs, vemos que los consumidores primarios produjeron 1103 kcal/m 2 /año a partir de los 7618 kcal/m 2 /año de energía disponibles para ellos de los productores primarios. La medición de la eficiencia de transferencia de energía entre dos niveles tróficos sucesivos se denomina eficiencia de transferencia de nivel trófico (TLTE) y se define por la fórmula:
\[\text{TLTE} = \dfrac{\text{production at present trophic level}}{\text{production at previous trophic level}} \times 100 \nonumber\]
En Silver Springs, el TLTE entre los dos primeros niveles tróficos fue de aproximadamente 14.8 por ciento. La baja eficiencia de la transferencia de energía entre niveles tróficos suele ser el factor principal que limita la longitud de las cadenas alimentarias observadas en una red alimentaria. El hecho es que, después de cuatro a seis transferencias de energía, no queda suficiente energía para soportar otro nivel trófico. En el ejemplo del lago Ontario, solo se produjeron tres transferencias de energía entre el productor primario, (algas verdes), y el consumidor ápice (salmón Chinook).
Los ecologistas tienen muchos métodos diferentes para medir las transferencias de energía dentro de los ecosistemas. Algunas transferencias son más fáciles o más difíciles de medir dependiendo de la complejidad del ecosistema y de cuánto acceso tienen los científicos para observar el ecosistema. Es decir, algunos ecosistemas son más difíciles de estudiar que otros, y a veces hay que estimar la cuantificación de las transferencias de energía.
Otro parámetro principal que es importante para caracterizar el flujo de energía dentro de un ecosistema es la eficiencia neta de producción. La eficiencia neta de producción (NPE) permite a los ecologistas cuantificar con qué eficiencia los organismos de un determinado nivel trófico incorporan la energía que reciben a la biomasa; se calcula utilizando la siguiente fórmula:
\[\text{NPE} = \dfrac{\text{net consumer productivity}}{\text{assimilation}} \times 100 \nonumber\]
La productividad neta del consumidor es el contenido energético disponible para los organismos del siguiente nivel trófico. La asimilación es la biomasa (contenido energético generado por unidad de área) del nivel trófico actual después de contabilizar la pérdida de energía por ingestión incompleta de alimentos, energía utilizada para la respiración y energía perdida como desperdicio. La ingestión incompleta se refiere al hecho de que algunos consumidores comen solo una parte de sus alimentos. Por ejemplo, cuando un león mata a un antílope, se lo comerá de todo excepto la piel y los huesos. Al león le falta la médula ósea rica en energía dentro del hueso, por lo que el león no hace uso de todas las calorías que su presa podría proporcionar.
Así, el NPE mide la eficiencia con la que cada nivel trófico usa e incorpora la energía de su alimento a la biomasa para alimentar el siguiente nivel trófico. En general, los animales de sangre fría (ectotermos), como invertebrados, peces, anfibios y reptiles, utilizan menos de la energía que obtienen para la respiración y el calor que los animales de sangre caliente (endotermas), como aves y mamíferos. El calor extra generado en las endotermas, aunque una ventaja en cuanto a la actividad de estos organismos en ambientes más fríos, es una desventaja importante en términos de NPE. Por lo tanto, muchas endotermas tienen que comer con más frecuencia que los ectotermos para obtener la energía que necesitan para sobrevivir. En general, la NPE para ectotermos es un orden de magnitud (10x) mayor que para las endotermas. Por ejemplo, el NPE para una oruga que come hojas se ha medido en 18 por ciento, mientras que el NPE para una ardilla que come bellotas puede ser tan bajo como 1.6 por ciento.
La ineficiencia del uso de energía por parte de animales de sangre caliente tiene amplias implicaciones para el suministro de alimentos del mundo. Es ampliamente aceptado que la industria cárnica utiliza grandes cantidades de cultivos para alimentar al ganado, y debido a que el NPE es bajo, gran parte de la energía de la alimentación animal se pierde. Por ejemplo, cuesta alrededor de 1¢ producir 1000 calorías dietéticas (kcal) de maíz o soja, pero aproximadamente $0.19 producir un número similar de calorías cultivando ganado para el consumo de carne. El mismo contenido energético de la leche del ganado bovino también es costoso, a aproximadamente $0.16 por 1000 kcal. Gran parte de esta diferencia se debe al bajo NPE del ganado. Así, ha habido un movimiento creciente a nivel mundial para promover el consumo de alimentos no cárnicos y no lácteos para que se desperdicie menos energía alimentando animales para la industria cárnica.
Modelando Ecosistemas Flujo de Energía: Pirámides Ecológicas
La estructura de los ecosistemas se puede visualizar con pirámides ecológicas, que fueron descritas por primera vez por los estudios pioneros de Charles Elton en la década de 1920. Las pirámides ecológicas muestran las cantidades relativas de varios parámetros (como el número de organismos, energía y biomasa) a través de los niveles tróficos.
Las pirámides de números pueden ser verticales o invertidas, dependiendo del ecosistema. Como se muestra en la Figura\(\PageIndex{3}\), los pastizales típicos durante el verano tienen una base de muchas plantas y el número de organismos disminuye en cada nivel trófico. Sin embargo, durante el verano en un bosque templado, la base de la pirámide consta de pocos árboles en comparación con el número de consumidores primarios, en su mayoría insectos. Debido a que los árboles son grandes, tienen una gran capacidad fotosintética y dominan otras plantas en este ecosistema para obtener luz solar. Aún en menor número, los productores primarios en los bosques siguen siendo capaces de soportar otros niveles tróficos.
Otra forma de visualizar la estructura del ecosistema es con pirámides de biomasa. Esta pirámide mide la cantidad de energía convertida en tejido vivo en los diferentes niveles tróficos. Usando el ejemplo del ecosistema de Silver Springs, estos datos exhiben una pirámide vertical de biomasa (Figura\(\PageIndex{3}\)), mientras que la pirámide del ejemplo del Canal de la Mancha está invertida. Las plantas (productoras primarias) del ecosistema Silver Springs constituyen un gran porcentaje de la biomasa que se encuentra allí. Sin embargo, el fitoplancton en el ejemplo del Canal de la Mancha conforman menos biomasa que los consumidores primarios, el zooplancton. Al igual que con las pirámides invertidas de números, esta pirámide invertida no se debe a la falta de productividad de los productores primarios sino que resulta de la alta tasa de rotación del fitoplancton. Los fitoplancton son consumidos rápidamente por los consumidores primarios, minimizando así su biomasa en cualquier momento en particular. Sin embargo, el fitoplancton se reproduce rápidamente, por lo que son capaces de soportar el resto del ecosistema.
El modelado de ecosistemas piramidales también puede usarse para mostrar el flujo de energía a través de los niveles tróficos Las pirámides de energía siempre están en posición vertical, y no se puede soportar un ecosistema sin suficiente productividad primaria. Todos los tipos de pirámides ecológicas son útiles para caracterizar la estructura de los ecosistemas. Sin embargo, en el estudio del flujo de energía a través del ecosistema, las pirámides de energía son los modelos más consistentes y representativos de la estructura del ecosistema.
Consecuencias de las redes alimentarias: aumento biológico
Una de las consecuencias ambientales más importantes de la dinámica de los ecosistemas es la biomagnificación. La biomagnificación es la concentración creciente de sustancias persistentes y tóxicas en los organismos en cada nivel trófico, desde los productores primarios hasta los consumidores ápice. Se ha demostrado que muchas sustancias se bioacumulan, incluyendo estudios clásicos con el pesticida diclorodifeniltricloroetano (DDT), que fue publicado en el bestseller de la década de 1960, Silent Spring, por Rachel Carson. El DDT era un pesticida de uso común antes de que se conocieran sus peligros. En algunos ecosistemas acuáticos, los organismos de cada nivel trófico consumieron muchos organismos del nivel inferior, lo que provocó que el DDT aumentara en las aves (consumidores ápice) que comían peces. Así, las aves acumularon cantidades suficientes de DDT para causar fragilidad en sus cáscaras de huevo. Este efecto incrementó la rotura del huevo durante la anidación y se demostró que tiene efectos adversos en estas poblaciones de aves. El uso del DDT fue prohibido en Estados Unidos en la década de 1970.
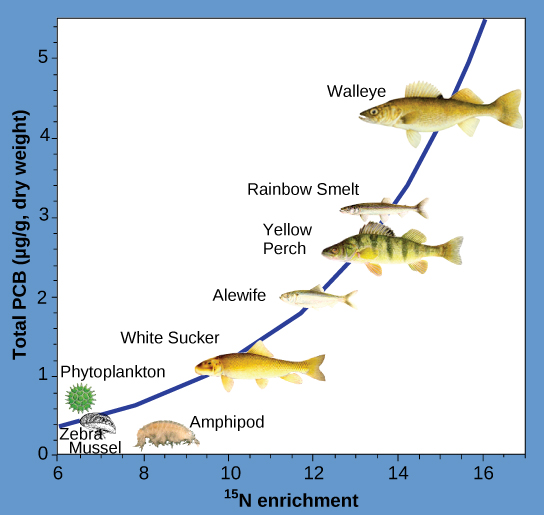
Otras sustancias que biomagnifican son los bifenilos policlorados (PCB), que se utilizaron en líquidos refrigerantes en Estados Unidos hasta que se prohibió su uso en 1979, y metales pesados, como mercurio, plomo y cadmio. Estas sustancias se estudiaron mejor en ecosistemas acuáticos, donde las especies de peces a diferentes niveles tróficos acumulan sustancias tóxicas traídas a través del ecosistema por los productores primarios. Como se ilustra en un estudio realizado por la Administración Nacional Oceánica y Atmosférica (NOAA) en la Bahía Saginaw del Lago Huron (Figura\(\PageIndex{4}\)), las concentraciones de PCB aumentaron desde los productores primarios del ecosistema (fitoplancton) a través de los diferentes niveles tróficos de las especies de peces. El consumidor ápice (walleye) tiene más de cuatro veces la cantidad de PCB en comparación con el fitoplancton. Además, con base en los resultados de otros estudios, las aves que comen estos peces pueden tener niveles de PCB al menos un orden de magnitud más altos que los encontrados en los peces del lago.
Otras preocupaciones han sido planteadas por la acumulación de metales pesados, como el mercurio y el cadmio, en ciertos tipos de mariscos. La Agencia de Protección Ambiental de Estados Unidos (EPA) recomienda que las mujeres embarazadas y los niños pequeños no consuman ningún pez espada, tiburón, caballa real o tilefish debido a su alto contenido de mercurio. A estos individuos se les aconseja comer pescado bajo en mercurio: salmón, tilapia, camarón, abadejo y bagre. La biomagnificación es un buen ejemplo de cómo la dinámica de los ecosistemas puede afectar nuestra vida cotidiana, incluso influyendo en los alimentos que comemos.
Resumen
Los organismos en un ecosistema adquieren energía de diversas maneras, la cual se transfiere entre niveles tróficos a medida que la energía fluye desde la parte inferior hasta la parte superior de la red alimentaria, perdiendo energía en cada transferencia. La eficiencia de estas transferencias es importante para comprender los diferentes comportamientos y hábitos alimenticios de los animales de sangre caliente versus de sangre fría. La modelización de la energía ecosistémica se realiza mejor con pirámides ecológicas de energía, aunque otras pirámides ecológicas proporcionan otra información vital sobre la estructura del ecosistema.
Referencias
OpenStax, Biología. OpenStax CNX. 26 de junio de 2020. https://cnx.org/contents/GFy_h8cu@10.137:noBcfThl@7/Understanding-Evolution.