
5.1: Replicación de ADN
- Page ID
- 53519

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)La única manera de hacer nuevas células es mediante la división de células preexistentes. Esto significa que todos los organismos dependen de la división celular para su existencia continuada. El ADN, como ustedes saben, lleva la información genética que necesita cada célula. Cada vez que una célula se divide, todo su ADN debe copiarse fielmente para que una copia de esta información pueda transmitirse a la célula hija. Este proceso se llama replicación del ADN. Antes de examinar el proceso real de replicación del ADN, es útil pensar en lo que se necesita para lograr esta tarea con éxito. Considere los desafíos que enfrenta una célula en este proceso:
- El gran número de nucleótidos a copiar es enorme: e.g., en células humanas, del orden de varios miles de millones.
- Se debe desenrollar una molécula de ADN parental de doble hélice para exponer cadenas individuales de ADN que puedan servir como moldes para la síntesis de nuevas cadenas de ADN.
- Este desenrollado debe realizarse sin introducir distorsión topológica significativa en la molécula.
- Se debe evitar que las cadenas individuales desenrolladas de ADN se vuelvan a unir el tiempo suficiente para que las nuevas cadenas sean sintetizadas.
- Las ADN polimerasas no pueden comenzar la síntesis de una nueva cadena de ADN de novo y requieren un OH 3' libre al que pueden agregar nucleótidos de ADN.
- Las ADN polimerasas solo pueden extender una cadena en la dirección 5' a 3'. La extensión de 5' a 3' de ambas nuevas hebras en una sola horquilla de replicación significa que una de las hebras está hecha en trozos.
- El uso de cebadores de ARN requiere que los nucleótidos de ARN se eliminen y reemplacen con nucleótidos de ADN y se deben unir los fragmentos de ADN resultantes.
- Asegurar exactitud en la copia de tanta información.
Con esto en mente, podemos comenzar a examinar cómo las células afrontan cada uno de estos desafíos. Nuestra comprensión del proceso de replicación del ADN se deriva de estudios que utilizan bacterias, levaduras y otros sistemas, como huevos de Xenopus. Estas investigaciones han revelado que la replicación del ADN se lleva a cabo por la acción de un gran número de proteínas que actúan juntas como una compleja máquina proteica llamada replisoma. Numerosas proteínas involucradas en la replicación han sido identificadas y caracterizadas, incluyendo múltiples ADN polimerasas diferentes tanto en procariotas como eucariotas. Si bien las proteínas específicas involucradas son diferentes en bacterias y eucariotas, es útil comprender las consideraciones básicas que son relevantes en todas las células, antes de intentar abordar los detalles de cada sistema.
A continuación se presenta un relato generalizado de los pasos en la replicación del ADN, enfocado en los desafíos mencionados anteriormente.
-
El gran número de nucleótidos a copiar es enorme.
Por ejemplo, en las células humanas, el número de nucleótidos a copiar es del orden de varios miles de millones. Incluso en bacterias, el número está en millones. Las células, ya sean bacterianas o eucariotas, tienen que replicar todo su ADN antes de que puedan dividirse. En células como la nuestra, la gran cantidad de ADN se descompone en muchos cromosomas, cada uno de los cuales está compuesto por una cadena lineal de ADN. En células como las de E. coli, hay un solo cromosoma circular.
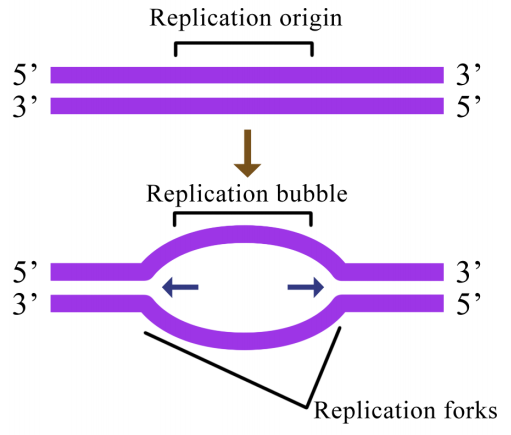
En cualquier situación, la replicación del ADN se inicia en sitios llamados orígenes de replicación. Estas son regiones de la molécula de ADN que son reconocidas por proteínas especiales de reconocimiento de origen que se unen al ADN. La unión de estas proteínas ayuda a abrir una región de ADN monocatenario donde puede comenzar la síntesis de nuevo ADN. En el caso de E. coli, existe un único origen de replicación en su cromosoma circular. En las células eucariotas, puede haber muchos miles de orígenes de replicación, teniendo cada cromosoma cientos. La replicación del ADN se inicia así en múltiples puntos a lo largo de cada cromosoma en eucariotas como se muestra en la Figura 5.1.3. Las micrografías de electrones de ADN replicante de células eucariotas muestran muchas “burbujas” de replicación en un solo cromosoma.

- Se debe desenrollar una molécula parental de doble hélice para exponer cadenas individuales de ADN que puedan servir como moldes para la síntesis de nuevas cadenas de ADN.
- Este desenrollado debe realizarse sin introducir distorsión topológica en la molécula.
- Se debe evitar que las cadenas individuales desenrolladas de ADN se vuelvan a unir el tiempo suficiente para que las nuevas cadenas sean sintetizadas.
Una vez separadas las dos cadenas de la molécula de ADN parental, se debe evitar que vuelvan a estar juntas para formar ADN bicatenario. Para garantizar que las regiones desenrolladas del ADN parental permanezcan monocatenarias y estén disponibles para copiarse, las cadenas separadas del ADN parental están unidas por muchas moléculas de una proteína llamada proteína de unión al ADN monocatenario (SSB).
-
Las ADN polimerasas no pueden comenzar la síntesis de una nueva cadena de ADN de novo y requieren un OH 3' libre al que pueden agregar nucleótidos de ADN.
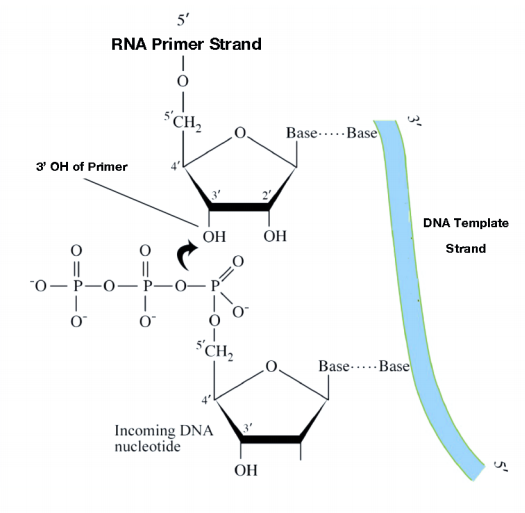
Aunque el ADN parental monocatenario ya está disponible para copiar, las ADN polimerasas no pueden comenzar la síntesis de una cadena complementaria de novo. Esto se debe a que todas las ADN polimerasas solo pueden agregar nuevos nucleótidos al extremo 3' de una cadena preexistente. Esto significa que alguna enzima que no sea una ADN polimerasa debe primero hacer una pequeña región de ácido nucleico, complementaria a la cadena parental, que pueda proporcionar un OH 3' libre al que la ADN polimerasa puede agregar un desoxirribonucleótido.
Esta tarea se logra mediante una enzima llamada primasa, que ensambla un tramo corto de ARN, llamado cebador, frente al molde de ADN parental. Esto proporciona una región corta de pares de bases con un grupo 3'OH libre al que la ADN polimerasa puede agregar el primer nucleótido de ADN nuevo (ver figura en la página anterior). Una vez que un cebador proporciona un 3'OH libre para extensión, otras proteínas entran en el acto. Estas proteínas están involucradas en la carga de la ADN polimerasa en el molde cebado y ayudan a mantenerla unida al ADN una vez que está encendida.
El primero de ellos es el cargador de abrazaderas. Como su nombre indica, la pinza cargadora ayuda a cargar un complejo proteico llamado pinza deslizante en el ADN en la horquilla de replicación. La pinza deslizante se une entonces por la ADN Polimerasa. La función de la pinza deslizante es aumentar la procesividad de la ADN polimerasa. Esta es una forma elegante de decir que mantiene la polimerasa asociada con la horquilla de replicación al evitar que se caiga; de hecho, la abrazadera deslizante ha sido descrita como un cinturón de seguridad para la ADN polimerasa.
La ADN polimerasa ahora está preparada para iniciar la síntesis de la nueva cadena de ADN (en E. coli, la polimerasa replicativa primaria se llama ADN polimerasa III). Como ya sabrás, la síntesis de nuevo ADN se logra mediante la adición de nuevos nucleótidos complementarios a los de la cadena parental. La ADN polimerasa cataliza la reacción mediante la cual se agrega un desoxirribonucleótido entrante en el extremo 3' del nucleótido anterior, comenzando con el 3'OH en el extremo del cebador de ARN.
El fosfato 5' en cada nucleótido entrante se une por la ADN polimerasa al OH 3' en el extremo de la cadena de ácido nucleico en crecimiento. Como ya señalamos, las nuevas cadenas de ADN se sintetizan mediante la adición de nucleótidos de ADN al extremo de un cebador de ARN. Por lo tanto, la nueva molécula de ADN tiene una pieza corta de ARN al principio.
-
Las ADN polimerasas solo pueden extender una cadena en la dirección 5' a 3'. El crecimiento de 5' a 3' de ambas nuevas hebras significa que una de las hebras está hecha en trozos.
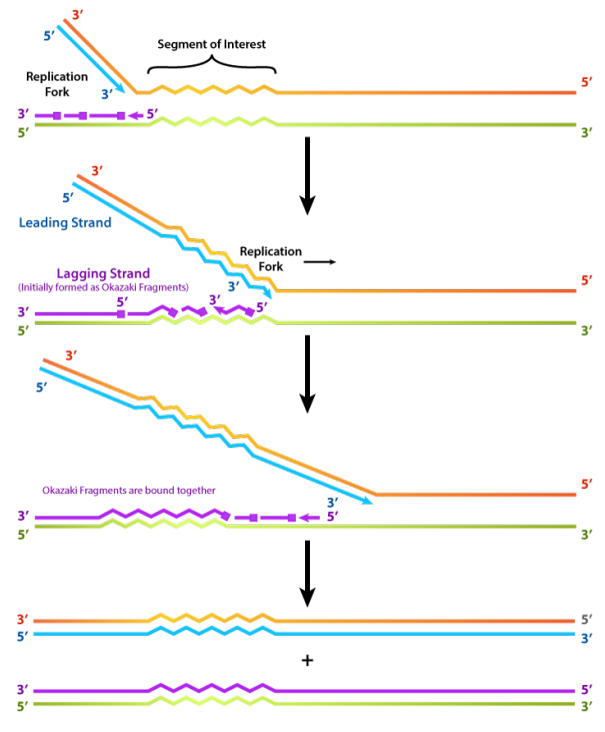
Hemos observado que la ADN polimerasa solo puede construir una nueva cadena de ADN en la dirección 5' a 3'. También sabemos que las dos cadenas parentales de ADN son antiparalelas. Esto significa que en cada horquilla de replicación, una nueva hebra, llamada la cadena principal, se puede sintetizar continuamente en la dirección 5' a 3' porque se está haciendo en la misma dirección en la que se está abriendo la horquilla de replicación.
La síntesis de la otra nueva cadena, llamada cadena rezagada, requiere que se establezcan múltiples cebadores de ARN y que el nuevo ADN se haga en muchas piezas cortas que se unen posteriormente. Estas piezas cortas de ácido nucleico, cada una compuesta por un pequeño tramo de cebador de ARN y alrededor de 1000-2000 nucleótidos de ADN, se denominan Fragmentos de Okazaki, para Reiji Okazaki, el científico que primero demostró su existencia.
- El uso de cebadores de ARN requiere que los nucleótidos de ARN se eliminen y reemplacen con nucleótidos de ADN.
- Hemos visto que cada pieza de ADN recién sintetizada comienza con un cebador de ARN, haciendo efectivamente una nueva cadena de ácido nucleico que es parte ARN y parte ADN. No se puede permitir que la cadena de ADN terminada tenga fragmentos de ARN unidos. o se eliminan los nucleótidos de ARN y los huecos se rellenan con nucleótidos de ADN (por ADN polimerasa I en E. coli). Las piezas de ADN son luego unidas entre sí por la enzima ADN ligasa.
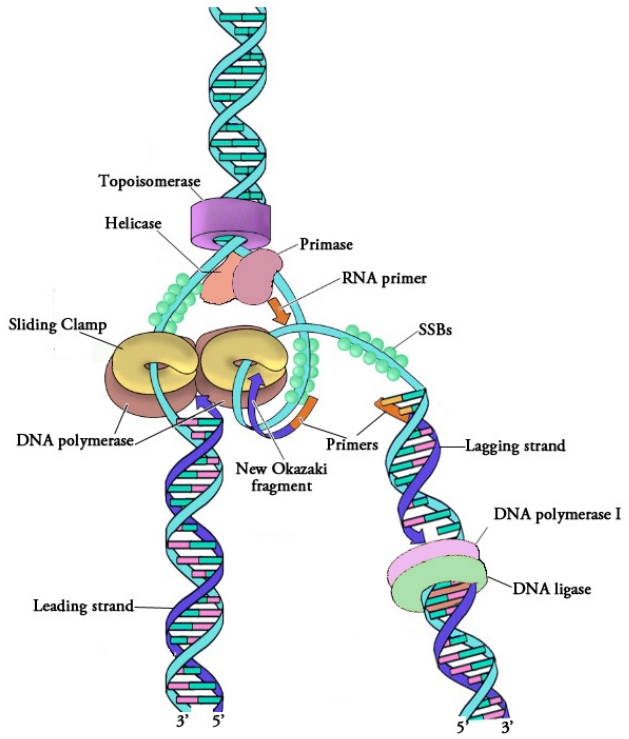
Figura 5.1.6: Proteínas en una horquilla de replicación
Los pasos descritos anteriormente completan esencialmente el proceso de replicación del ADN. La Figura 5.1.6 muestra una horquilla de replicación, completa con las proteínas asociadas que forman el replisoma.
- Asegurar precisión en la copia de tanta información
¿Qué tan precisa es la copia de la información en el ADN por la ADN polimerasa? Como ustedes saben, los cambios en la secuencia del ADN (mutaciones) pueden cambiar la secuencia de aminoácidos de las proteínas codificadas y que esto suele ser, aunque no siempre, deletéreo para el funcionamiento del organismo. Cuando se copian miles de millones de bases en el ADN durante la replicación, ¿cómo aseguran las células que el ADN recién sintetizado sea una copia fiel de la información original?
Las ADN polimerasas, como hemos señalado anteriormente, funcionan rápido (promediando 50 bases por segundo en células humanas y hasta 20 veces más rápido en E. coli). Sin embargo, tanto las células humanas como las bacterianas parecen replicar su ADN con bastante precisión. Esto se debe a la función de corrección de pruebas de las ADN polimerasas. La función de corrección de pruebas de una ADN polimerasa permite a la polimerasa detectar cuándo se ha insertado la base incorrecta frente a una cadena molde, retroceder y eliminar la base insertada erróneamente. Esto es posible porque la polimerasa es una enzima de doble función. Puede extender una cadena de ADN en virtud de su actividad polimerasa 5' a 3' pero también puede retroceder y eliminar la última base insertada porque tiene una actividad exonucleasa de 3' a 5' (una exonucleasa es una enzima que elimina bases, una a una, de los extremos de los ácidos nucleicos). La actividad exonucleasa de la ADN polimerasa le permite escindir una base insertada erróneamente, después de lo cual la actividad polimerasa inserta la base correcta y procede con la extensión de la cadena.
En otras palabras, la ADN polimerasa está monitoreando su propia precisión (también denominada su fidelidad) a medida que produce nuevo ADN, corrigiendo errores inmediatamente antes de pasar a agregar la siguiente base. Este mecanismo, que opera durante la replicación del ADN, corrige muchos errores a medida que ocurren, reduciendo en aproximadamente 100 veces los errores cometidos cuando se copia el ADN.