
5.1: Introducción
- Page ID
- 56361

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)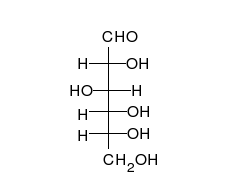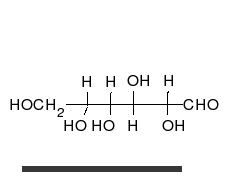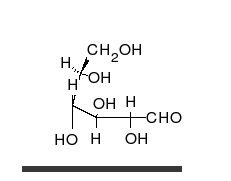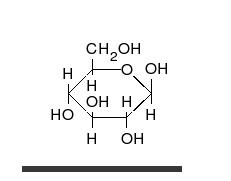
La glucosa es el carbohidrato preferido de las células. En solución, puede cambiar de una cadena lineal a un anillo.
La energía se almacena en los enlaces de los carbohidratos. Romper estos lazos libera esa energía. La trituración de cristales de azúcar crea pequeños campos eléctricos que desprenden luz ultravioleta invisible. El químico wintergreen (salicilato de metilo) se excita por estos electrones excitados y fluoresce en una longitud de onda azul visible. Este fenómeno se llama triboluminiscencia.
Glicólisis
La glucosa es el carbohidrato preferido de las células. La glucólisis (glicólico-azúcar; lisis - división) es un proceso universal de todas las células que ocurre en el citosol mediante el cual la glucosa (un azúcar de 6 carbonos) se divide en dos moléculas de piruvatos (una molécula de 3 carbonos) para generar ATP y NADH reducido. El ATP (trifosfato de adenosina) es la moneda de energía de la célula que almacena energía química en 3 enlaces fosfato de alta energía. El NADH (dinucleótido reducido de nicotinamida adenina) es un portador de electrones de alta energía que actúa como coenzima en las reacciones y como una especie de batería recargable. El estado sin carga que no lleva electrones de alta energía se llama NAD +.
La glucólisis es la división de la glucosa en 2 moléculas de piruvato para generar 2 moléculas de NADH y 2ATP.

El ATP contiene 3 fosfatos de alta energía y actúa como moneda de energía celular.

NADH es la forma reducida de NAD+. Los electrones de alta energía asociados con la forma reducida vienen con un átomo de hidrógeno.
Fermentación
En ausencia de oxígeno, las células pueden decidir utilizar el piruvato de la glucólisis para generar rápidamente moléculas de ATP adicionales en un proceso llamado fermentación. La fermentación es el proceso anaeróbico de reducción del piruvato para generar ATP. Este proceso utiliza el NADH generado a partir de la glucólisis como los agentes reductores. La fermentación es un proceso familiar que ocurre en la levadura para generar etanol. En otros organismos, como los humanos, la fermentación da como resultado la producción de ácido láctico. Tanto el ácido láctico como el etanol son tóxicos, pero esto ayuda a las células a generar ATP cuando se requiere energía rápidamente. La fermentación también genera CO 2 como molécula de desecho ya que el piruvato se descompone en un compuesto de 2 carbonos.
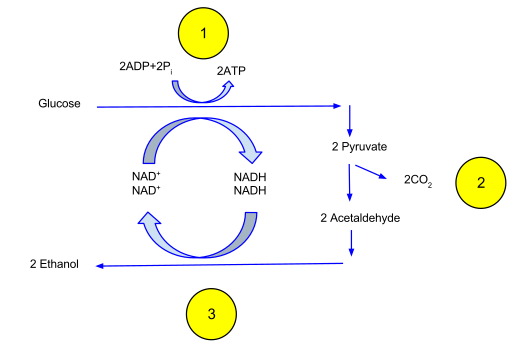
La fermentación en levadura genera ATP en ausencia de oxígeno pero produce poco ATP a costa del NADH reducido. Crédito: Davidcarmack (CC-BY-SA)
La reacción preparatoria
En presencia de O 2, los organismos aeróbicos utilizarán una reacción de descarboxilación de piruvato en el citosol. Esta reacción genera una molécula de acetil-CoA a partir de la Coenzima A que puede ingresar a las mitocondrias.
La coenzima A (CoA) se carga con un grupo acetilo (compuesto de 2 carbonos) para generar acetil-CoA y un CO 2.
Cuando hay un exceso de carbohidratos, el acetil-CoA se utiliza como punto de partida para el almacenamiento de energía a largo plazo en la síntesis de lípidos.
Mitocondrias
Las mitocondrias son la central eléctrica de las células eucariotas. Se derivan de un proceso descrito por la teoría endosimbiótica mediante el cual procariotas aeróbicos fueron engullidos por un proto-eucariota. En este arreglo mutualista, el procariota desintoxicó el mortal gas O 2 en el ambiente y lo utilizó para descomponer completamente la glucosa y producir muchas moléculas de ATP. La evidencia de esta teoría proviene de la replicación independiente de las mitocondrias, el ADN mitocondrial similar a bacterias, los ribosomas mitocondriales similares a bacterias, los lípidos bacterianos que se encuentran en la membrana interna y la naturaleza eucariota de la membrana externa. Las mitocondrias son genómicamente similares a las bacterias del orden Rickettsiales. Algunas bacterias de este orden siguen viviendo en libertad y otras son patógenos intracelulares.
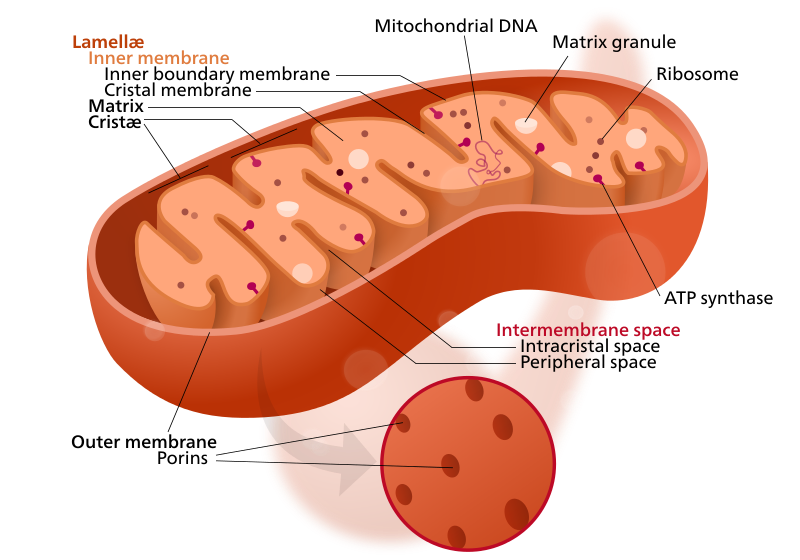
Crédito: Kelvinsong (CC-BY- SA 3.0)
Respiración aeróbica

Respiración Celular. El lado izquierdo es la glucólisis (anaeróbica). El lado derecho es lo que ocurre en presencia de oxígeno en eucariotas. Las reacciones aeróbicas ocurren dentro de las mitocondrias después de ser alimentadas con moléculas de acetil-CoA de la reacción preparatoria citoplásmica. Crédito: RegisFrey (CC-BY-SA 3.0)
El acetil-CoA ingresa a la matriz mitocondrial donde se usa en el Ciclo Krebs (también conocido como Ciclo del ácido tricarboxílico (TCA), también conocido como Ciclo del ácido cítrico). Por cada piruvato, hay 2 vueltas del ciclo donde se generan NADH adicional y otro portador de electrones de alta energía FADH 2 (flavina adenina dinucleótido). Los electrones almacenados por NADH y FADH 2 son transferidos a proteínas llamadas citocromos que tienen centros metálicos para conducir estos electrones. En el proceso de mover estos electrones, los citocromos en estas Cadenas de Transporte de Electrones (ETC) alimentan el movimiento de protones hacia el espacio intermembrana. El término de estos electrones es una molécula O 2 que se reduce en moléculas 1/2 H 2 O. Este aparente movimiento de las moléculas de agua a partir de la síntesis química se denomina quimiósmosis. Un canal en la membrana llamado ATP sintasa actúa como puerta de entrada para que el H + vuelva a la matriz, pero usa este movimiento para convertir ADP en ATP.
Primer plano de la Cadena de Transporte de Electrones (ETC) que tiene lugar en la membrana interna de las mitocondrias. Aquí es donde se utiliza oxígeno como aceptor de electrones final. La reducción de 1/2 O 2 da como resultado la generación de una molécula de agua (quimiósmosis). Crédito: Jeremy Seto (CC-BY-NC-SA 3.0)
Alberca Metabólica
Las vías catabólicas involucradas en la glucólisis y el ciclo de Krebs constituyen el pool metabólico que suministra bloques de construcción para otras reacciones anabólicas en la célula. Un exceso de carbohidratos puede resultar en una acumulación de moléculas de acetil-CoA. Si hay un gran exceso de acetil-CoA, los grupos acetilo pueden comprometerse con la síntesis de ácidos grasos para el almacenamiento de energía a largo plazo. Los productos glicolíticos también pueden ser el punto de partida para la síntesis de aminoácidos. El 3-fosfoglicerato se puede utilizar para sintetizar glicina, cisteína y serina. El piruvato se puede usar para generar alanina, valina y leucina. El oxaloacetato del ciclo de Krebs se puede utilizar como punto de partida para aspartato, lisina, asparagina, metionina, treonina e isoleucina. El glutamato y la glutamina se sintetizan a partir de α-cetoglutarato formado durante el ciclo de Krebs. Si bien la mayoría de los 20 aminoácidos se pueden sintetizar de novo, existen 9 aminoácidos esenciales en humanos que no pueden sintetizarse en cantidad suficiente y por lo tanto deben obtenerse de la dieta. Estos aminoácidos esenciales incluyen histidina, isoleucina, leucina, lisina, metionina, fenilalanina, treonina, triptófano y valina.