
1.4: Forma del organismo- composición, tamaño y forma
- Page ID
- 56442

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
En la famosa escena de bares de Star Wars, se intentó ilustrar diversos organismos. Fue un esfuerzo notablemente inimaginativo: todas las formas de vida ilustradas tenían un parecido sorprendente con los humanos, todas aproximadamente del mismo tamaño y con los mismos componentes básicos (por ejemplo, cabeza, piernas, brazos). Sin salir de este planeta se puede llegar a un conjunto mucho más diverso de organismos, como espero lo demuestre este capítulo. Los organismos varían entre sí de múltiples maneras pero en este capítulo, me centraré en tres aspectos, dos de los cuales, el tamaño y la forma, son muy sencillos, pero el tercer aspecto, la composición, es menos familiar. Estas características han sido moldeadas por la evolución, y abundan la convergencia (grupos no relacionados que convergen en una forma común) y la divergencia (grupos relacionados que muestran una variedad de formas). En consecuencia, aunque la forma, el tamaño y la composición son extremadamente importantes para la biología de un organismo, son indicadores notablemente pobres de filogenia. Esto hace que intentar caracterizar la forma de entidades taxonómicas más grandes (clase, orden, filo) sea desafiante, si no imposible. La composición, el tamaño y la forma están interrelacionados y son significativos con otros aspectos de la biología de un organismo. Además, también suelen ser dinámicas, cambiando durante la existencia de un organismo.

TEMAS
- Composición
- Unicelular
- Multicelular
- Colonial
- Coenocítico
- Tamaño
- Consideraciones de tamaño y forma
- Forma del organismo: esferas, aplanadas, cilindros
Composición
¿De qué están compuestos los organismos, es decir, de qué están construidos? Un capítulo posterior considerará la composición en términos químicos y moleculares. En este capítulo 'composición' se refiere a la naturaleza celular de un organismo. La mayoría estaría de acuerdo en que todos los organismos están hechos de células, pero ¿es una célula o muchas? ¿Y las células son todas iguales o difieren? ¿Y las células son 'típicas' en su organización? Aquí hay cuatro tipos de composición, dos de los cuales son muy familiares, y dos de los cuales son menos:
- organismos unicelulares — el organismo es una sola célula uninucleada
- organismos multicelulares ('multicelularidad compleja'): el organismo está hecho de múltiples (generalmente muchas) células pero las células son de diferentes formas y tienen un arreglo organizado específico que involucra tejidos y órganos
- composición colonial ('multicelularidad simple') — el organismo está hecho de múltiples células pero las células son generalmente similares en tamaño y forma sin tejidos y órganos obvios
- composición coenocítica — el organismo está hecho de células 'atípicas', atípicas en que tienen múltiples núcleos, a veces miles de ellos. Todo el organismo a veces es solo una célula que tiene muchos núcleos, o puede estar compuesto por múltiples unidades, que podrían llamarse células, excepto que generalmente son más grandes de lo normal y tienen múltiples núcleos.
Organismos Unicelulares
A pesar de que las plantas y los animales, los dos grupos de organismos con los que la mayoría de la gente está familiarizada, son multicelulares, la composición unicelular es con mucho el tipo de organismo más común. Claramente es una forma exitosa de construir un organismo y la mayor parte de lo 'vivo' que sucede en la tierra es consecuencia de la vida unicelular. Como demostrarán los ejemplos de unicelulares que aquí se tratan, existe una tremenda diversidad en los organismos unicelulares.
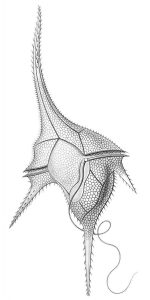
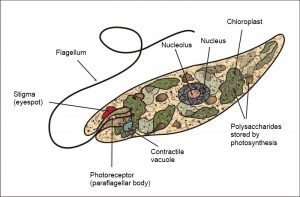
Los organismos unicelulares NO siempre son simples en forma y función. Pueden ser muy elaborados en forma por ejemplo, diatomeas (por ejemplo, Thalassiosira) y dinoflagelados (Figura 2). Si bien cualquier célula es una entidad compleja, existen muchos organismos unicelulares con componentes funcionalmente análogos a los órganos (multicelulares) de organismos más familiares. Por ejemplo, las células de Euglena poseen una notable cantidad de organización interna, incluyendo orgánulos (mancha ocular, vacuola contráctil) que realizan funciones cubiertas por órganos enteros (ojo, riñón) en organismos multicelulares. Además, Euglena (Figura 3) (junto con muchas bacterias y algas unicelulares) puede moverse como resultado del reposicionamiento coordinado de extensiones que no son patas, alas o aletas multicelulares sino una extensión de la célula individual llamada flagelo; adicionalmente, Euglena y otros organismos unicelulares también pueden moverse alterando la forma del cuerpo celular, comparable a lo que hacen las lombrices de tierra, pero sin la ayuda de los músculos y otros tejidos.
La mayoría de los organismos pasan por una etapa o etapas que es/son unicelulares (por ejemplo, esporas, gametos, cigotos) y estas unicelulas, muchas de las cuales pueden considerarse organismos porque a menudo son distintas espacial y temporalmente, suelen ser muy elaboradas en estructura y función. Los ejemplos que se discutirán más adelante son los espermatozoides de múltiples grupos de plantas (por ejemplo, helechos, musgos), a menudo denominados espermatozoides (Figura 4).

Si bien ser unicelulares se considera legítimamente una condición primitiva, las levaduras son hongos unicelulares y demuestran que la condición unicelular a veces puede 'reaparecer' en un linaje que anteriormente tenía algún otro tipo de composición, es decir, que la unicelularidad es a veces una condición 'derivada'. La gran mayoría de los hongos son coloniales/multicelulares pero las levaduras han aparecido independientemente varias veces en varios grupos fúngicos diferentes y NO representan la condición primitiva. En muchas levaduras el hábito de crecimiento unicelular está ligado a las condiciones ambientales y el organismo puede ser inducido a crecer en una materia colonial ajustando las condiciones.
Organismos coloniales

Las colonias de células (organismos coloniales) pueden formarse de dos maneras. La forma menos común es que las células individuales se unan para formar una colonia. Este comportamiento ha evolucionado múltiples veces, varias veces en las bacterias (en grupos descritos como 'bacterias sociales') y también en los mohos celulares del limo como Dictyostelium. (Figura 5) Las colonias producidas de esta manera contendrán células que no necesariamente son todas iguales genéticamente. La segunda y mucho más común forma de formar una colonia es el resultado de repetidas divisiones celulares donde las células hijas se adhieren a su progenitor. Para una célula con pared celular, la adhesión entre células hijas se logra con una capa adhesiva depositada entre las nuevas paredes celulares que se producen durante la citocinesis.

En los organismos coloniales, las divisiones celulares repetidas producen un organismo con muchas células unidas entre sí pero las células son todas (o casi todas) idénticas. Este tipo de composición a veces se describe como 'multicelularidad simple' o 'plurocelularidad' que se distingue de 'multicelularidad verdadera' o 'compleja multicelularidad ', o a veces simplemente' multicelularidad 'por un conjunto variable de criterios que suelen incluir la diferenciación celular y la producción de tejidos (más sobre esto en el siguiente capítulo). Una amplia variedad de formas coloniales son posibles y resultan de los patrones en los planos de división celular, por ejemplo, se forman filamentos (Figura 6) si el plano de división celular es siempre el mismo, produciendo una cadena de células conectadas de arriba a abajo.

Los filamentos pueden crecer desde el extremo basal (si la célula original continúa dividiéndose pero las células hijas NO se dividen) o pueden crecer desde la punta (si la célula original NO se divide y la célula hija se divide repetidamente) o pueden crecer en todo el filamento. Los filamentos pueden ramificarse si algunas celdas de un filamento se dividen en una dirección paralela a la dirección del filamento; láminas aplanadas (Figura 7) se producen si las células se dividen repetidamente en dos planos perpendiculares; se producen cúmulos globulares de células si la célula original y sus derivadas se dividen repetidamente en múltiples planos.
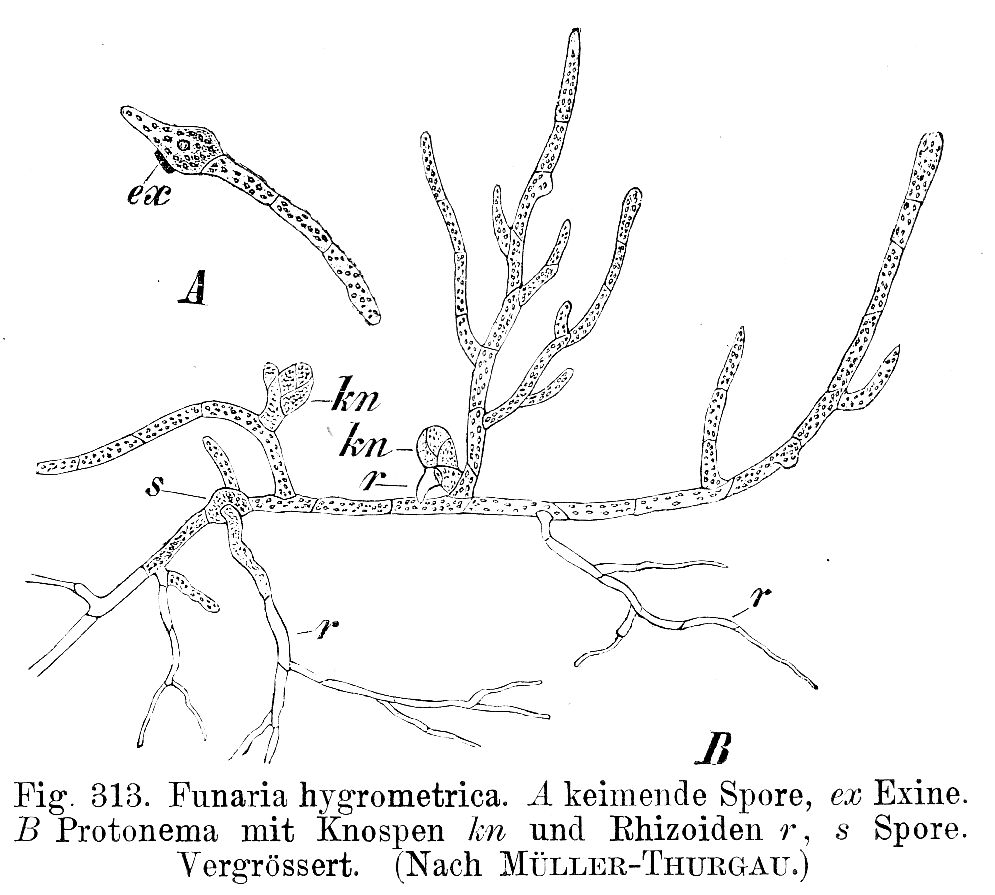
El crecimiento filamentoso se encuentra en arqueas, bacterias, (Figura 8) y hongos y muchos de los grupos que antes se ponían en el Reino Protista, en particular los grupos de algas verdes, rojas y pardas y algunas diatomeas. También se encuentra como una etapa breve en la mayoría de musgos y helechos (Fig. 10).

La mayoría de los hongos exhiben un tipo distinto de crecimiento filamentoso donde los filamentos, llamados hifas, crecen desde la punta pero comúnmente se ramifican, tanto en la punta como por debajo de ella, y con las ramas capaces de fusionarse con otras hifas para formar lo que se llama micelio, un interconectado, masa anastomosante de filamentos de hifa l (Figura 10).

El crecimiento colonial bidimensional, formando láminas de una a varias celdas de espesor, se encuentra más comúnmente en formas fotosintéticas, principalmente en los grupos de algas. También está presente en algunas de las plantas no vasculares: todas las hornworts y algunas de las hepáticas (Figura 11), donde se presenta en forma haploide (formadora de gametos) pero no en forma diploide (productora de esporas) (ver Capítulo 13). Ocasionalmente se encuentran colonias esféricas, particularmente en las algas verdes.

Las ventajas del crecimiento colonial, es decir, de que las células se agreguen o que las células hijas permanezcan unidas a las células parentales, pueden ser diferentes en diferentes situaciones. Una ventaja es el tamaño; permite organismos mucho más grandes que una sola célula y el tamaño más grande es a veces, pero no siempre, ventajoso. Como se analiza a continuación, el tamaño afecta la interacción entre los organismos y su entorno. El crecimiento colonial puede ser exitoso porque permite la exploración del hábitat: un organismo en un hábitat marginal puede ser sostenido mientras explora pastos más verdes. El mayor tamaño de las colonias hace posible la explotación de diferentes hábitates/regiones. Por ejemplo, las algas son capaces de explotar tanto un sustrato (para fijación) como la columna de agua (para luz y nutrientes). La mayoría de los trabajadores sienten que el crecimiento colonial (multicelularidad simple) es un paso en el camino hacia la multicelularidad compleja, pero la abundancia de formas coloniales altamente exitosas (es decir, diversas y abundantes) indican que el crecimiento colonial es en sí mismo un tipo de composición ventajosa.
Organismos coenocíticos

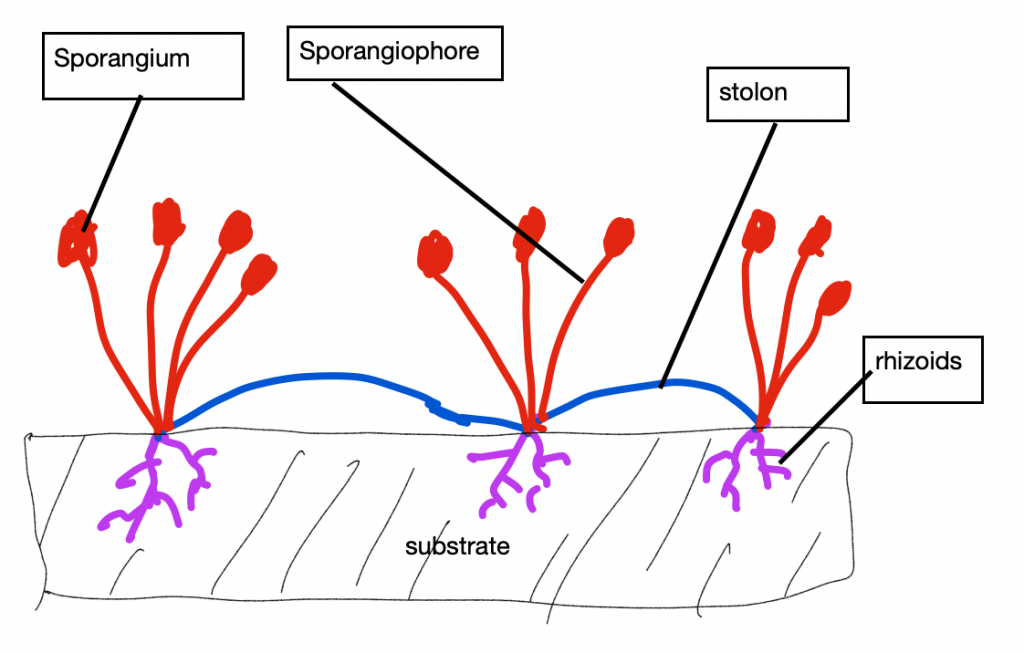
Probablemente el tipo de composición menos familiar es el coenocítico, donde las células del organismo, o a veces su única célula, es multinucleada. Esto es el resultado de divisiones mitóticas que no van acompañadas de citocinesis, a menudo repetidamente así. Parte de la significación de la condición coenocítica tiene que ver con el tamaño. El tamaño celular puede ser limitado porque un solo núcleo solo puede controlar un volumen limitado de citosol y para que la célula se haga más grande es necesario que haya núcleos adicionales. Las células coenocíticas suelen ser grandes y a veces extraordinariamente. Un caso extremo se encuentra en los moldes de limo plasmodial (por ejemplo, Physarum), un organismo similar a la ameba que consiste en una sola célula sin pared que puede tener hasta un metro de longitud con cientos de miles de núcleos (Figura 12). La forma de los mohos de limo es dinámica, debido a la capacidad del citoplasma para fluir en canales dentro de la célula individual, permitiendo así que el organismo explore su entorno, engulle comida e incluso trepe obstáculos. A diferencia de los mohos plasmodiales sin pared, los organismos coenocíticos con paredes pueden producir formas específicas y permanentes, pero no la forma en que esa forma se crea típicamente en organismos multicelulares (agregando células para crear una forma), sino como resultado de la creación de forma en células grandes individuales. Los moldes de pan (Rhizopus). producen rizoides, estolones y estructuras reproductivas acechadas, todas formadas a partir de una sola célula con múltiples núcleos (Figura 13).
De igual manera, la notoria alga verde invasora Caulerpa, puede aparecer muy parecida a la planta, es decir, con 'tallo y hojas', pero su notable tamaño y complejidad ocurre sin recurrir a células y tejidos.
Muchos organismos coenocíticos son muy similares en forma a los organismos coloniales, más comúnmente filamentos pero también láminas y esferas. La estructura 'fúngica' de la hifaa/micelio que es celular (i.e., colonial) en los hongos saco (Ascomycota) y en los hongos club (Basidiomycota) es coenocítica en los mohos del pan (Zygomycota) y en la micorriza que forma Glomeromycota. Y esta misma forma coenocítica similar a hongos se encuentra en un grupo no fúngico, los mohos de agua, por ejemplo, el patógeno vegetal Phytophthora. La estructura de los moldes de agua explica por qué el grupo solía colocarse en el filo fúngico y también por qué agrupar las cosas por 'miradas', es decir, forma, ¡es peligroso! Los mohos de agua ahora se colocan en un grupo que incluye algas pardas y coccolítoforos, dos grupos que no son para nada fúngicos ni coenocíticos.

Muchos no considerarían que estos organismos coenocíticos fueran unicelulares, aunque técnicamente muchos lo son. Están más cerca de un organismo colonial. Si se define órgano como un componente de un organismo que proporciona una función particular (por ejemplo, anclaje, elevación) entonces es evidente que los órganos pueden formarse tanto en organismos coloniales como coenocíticos (Figura 14), en ambos casos sin la presencia de células y tejidos especializados.
Organismos Multicelulares
La mayoría de los trabajadores no unificarían todos los organismos compuestos por dos o más células como 'multicelulares' pero desarrollar criterios para definiciones útiles (por ejemplo, multicelularidad 'simple vs. compleja') es un desafío y no hay un acuerdo universal sobre dónde trazar líneas. Para la mayoría de los investigadores, la multicelularidad 'verdadera' implica la especialización de los tipos celulares, tema que se considerará en el próximo capítulo.
Tamaño del organismo
Los organismos varían enormemente tanto en volumen como en su extensión en tres dimensiones. Los organismos más pequeños (Cuadro 1) son procariotas (bacterias y arqueas), organismos unicelulares que carecen de núcleos u otros orgánulos celulares, con dimensiones más largas típicamente de unos pocos um (= 10 -6 mm) y volúmenes de menos de 1 um 3 (= 1 femolitro, donde mil millones (10 9 ) femolitres es igual a 1 ul; un cuatrillón (10 15) femolitres equivale a 1 litro). Estos pequeños organismos suelen tener la forma de esferas, varillas o espirales. Aunque la mayoría de las células procariotas varían de 0.5 a 10 um, hay algunos procariotas gigantes cuyas células podrían tener hasta 500 um de longitud, lo que significa que se pueden ver a simple vista, aunque como una mota.

Los eucariotas unicelulares poseen células con núcleos y otros orgánulos celulares. Generalmente son más grandes (10-100 um) que los procariotas aunque hay una serie de eucariotas unicelulares con células pequeñas, en particular las levaduras (hongos unicelulares generalmente con células menores de 10 um). El hecho de que los organismos unicelulares son casi siempre de tamaño pequeño generalmente se piensa que es una consecuencia de la necesidad de control nuclear de la actividad celular; mayor tamaño es difícil porque en las células grandes los mensajes (ARNm, proteínas) que se mueven por difusión tardan demasiado en llegar del 'control center' (el núcleo, el ribosoma) a todas las partes de la célula. Esta idea se sustenta en las siguientes observaciones:
- Las células coenocíticas, que tienen múltiples núcleos, suelen ser mucho más grandes que las células con un solo núcleo
- unicelulas más grandes a menudo exhiben flujo citoplásmico que proporciona un movimiento más rápido de las señales a través de la célula
- unicelulares más grandes (que se encuentran en plantas, hongos y algas) poseen una gran vacuola central que ocupa gran parte del volumen celular y que es de alguna manera un 'espacio muerto' metabólico que es relativamente inactivo (o al menos no tan dinámico como el citosol); en consecuencia, la cantidad de citosol que el núcleo debe ' control' es en realidad mucho más pequeño de lo que parece basado en el tamaño de celda
Existen algunos organismos unicelulares extremadamente grandes, Acetabularia, (Figura 16) de pie de hasta 5 cm de altura, siendo uno de ellos. La acetabularia es única no sólo por su tamaño sino también porque demuestra que los 'órganos' ('rizoides', 'tallos', 'hojas') son posibles incluso en organismos que son unicelulares. Las células de acetabularia son más de 1000 veces más grandes que las células eucariotas típicas, con dimensiones máximas de más de 50, 000 um (= 50 mm = 5 cm).

Otras unicelulas grandes, por ejemplo, moho de pan (Rhizopus), Physarum (moho de limo plasmodial) son coenocíticas y se podría hacer el caso de que realmente no deberían considerarse unicelulares.
La mayoría de los organismos de más de 100 um de tamaño son coloniales, coenocíticos o multicelulares. Si bien las células del organismo coenocítico varían mucho en tamaño, las células de los organismos coloniales y multicelulares son comúnmente de 10 a 100 um en su dimensión máxima y el tamaño general del organismo está determinado por cuántas células se producen/acumulan. Algunos de los organismos particularmente grandes que consideraremos son: secuoyas, de hasta 85 m de altura y con un peso estimado de 2100 toneladas métricas (= 2100 * 10 3 kg = 4620 * 10 3 libras = 2310 toneladas); algas gigantes (algas pardas) de hasta 50 m de longitud; 'Pando', un clon de álamo tembloroso que pesan s más de 6 millones de kg, se extienden s sobre 43 hectáreas y pueden tener una edad de 80, 000 años (más en la discusión de Populus); y un hongo de miel (Armillaria) que se extiende sobre 4 millas cuadradas (1000 hectáreas), pesa aproximadamente 55, 000 kg y se considera que tiene 2400 años de antigüedad.

Tamaño y forma: influencia en el área de superficie y las interacciones con el medio ambiente
El tamaño y la forma son particularmente significativos porque dictan el grado de interacción entre el organismo y el ambiente exterior. Apreciar que las condiciones dentro de los organismos son distintas del exterior; esto es parte de lo que define la vida. La segunda ley de la termodinámica (que consideraremos en el Capítulo 24) dicta que las diferencias entre el interior y el exterior disminuyen con el tiempo: si algún químico se concentra dentro de un organismo tenderá a filtrarse; si algo se excluye de un organismo tenderá a filtrarse; si un organismo es más cálido que su ambiente se enfriará; si un organismo es más fresco que su ambiente se calentará. Cualesquiera que sean las condiciones que un organismo desarrolle para promover sus funciones vitales, tenderán a desaparecer porque la segunda ley dicta que los sistemas cambien para desarrollar uniformidad, es decir, hay una tendencia a que el interior se vuelva más como el exterior. Ante esto, uno podría pensar que tener poca interacción con el entorno externo es 'lo mejor'. Sin embargo, la interacción con el medio ambiente es esencial: (1) para obtener materiales —alimentos, oxígeno, minerales— que son necesarios para mantener la vida, y (2) librar al organismo de los 'materiales' que produce, por ejemplo, calor y dióxido de carbono, que le causarán daño si se permite que se acumule. El tamaño y la forma de un organismo controlan cómo interactúa con el entorno en el que se encuentra. Significativamente, el tamaño y la forma afectan dos parámetros importantes que están relacionados pero no exactamente lo mismo, la superficie de contacto con el ambiente y el volumen del ambiente en proximidad con el organismo. Ambos parámetros son importantes para controlar la interacción entre el organismo y su entorno.
La importancia de la superficie es familiarizada.Más superficie permite una mayor interacción con el ambiente, lo que puede o no ser beneficioso.Para cualquier forma, los objetos más pequeños siempre tienen una mayor superficie por unidad de volumen que los objetos grandes (Tabla 1).
|
forma |
dimensión (en unidades arbitrarias) r = radio, s = lado |
volumen (unidades 3) |
superficie (unidades 2) |
superficie: volumen (unidades -1) |
|---|---|---|---|---|
|
esfera |
r = 0.1 unidades |
0.004 |
0.126 |
31.5 |
|
esfera |
r = 1.0 unidades |
4.19 |
12.6 |
3.01 |
|
esfera |
r = 10 unidades |
4190 |
1257 |
0.3 |
|
cubo |
s = 0.1 unidades |
0.001 |
0.06 |
60 |
|
cubo |
s =1.0 unidades |
1 |
6 |
6 |
|
cubo |
s = 10 unidades |
1000 |
600 |
0.6 |
|
cilindro |
r = 0.1, altura = 10 x r |
0.314 |
0.691 |
22 |
|
cilindro |
r= 1.0, altura = 10 x r |
3.14 |
69.1 |
2.20 |
|
cilindro |
r= 10, altura = 10 x r |
31.4 |
6911 |
0.22 |
|
fórmulas |
esfera: volumen = 4/3 (pi) (r) 3; superficie = 4 (pi) (r) 2 |
|||
|
cubo: volumen = (s) 3; superficie = 6 (s) 2 |
||||
|
cilindro: volumen = altura (pi) (r) 2; superficie específica = 2 (pi) (r) (altura) + 2 (pi) (r) 2 |
||||
Manteniendo el volumen constante, una esfera tiene el área superficial mínima de cualquier forma, y las desviaciones de una forma isodiamétrica aumentan el área superficial y, por lo tanto, la relación área superficial:volumen (Cuadro 2). Si se compara el área superficial de dos formas comunes, 'filamentos' (alargados en una dimensión) y 'láminas' (alargadas en dos dimensiones) mientras se mantiene constante el volumen, el grado de elongación aumenta el área superficial y el alargamiento en dos dimensiones tiene un mayor efecto que el alargamiento en una dimensión (Cuadro 3).
|
forma |
dimensiones |
área de superficie = superficie/volumen |
|
esfera |
radio = 0.62 |
4.83 |
|
cubo |
lado = 1 |
6.0 |
|
cilindro |
diámetro = largo = 1.08 |
5.49 |
|
forma |
dimensiones |
área de superficie = superficie/volumen |
|
hilo |
largo = 10 x diámetro |
8.34 |
|
hilo |
largo = 100 x diámetro |
17.28 |
|
disco |
diámetro = 10 x largo |
10.32 |
|
disco |
diámetro = 100 x largo |
40.69 |
|
rosca cuboidal |
0.464 x 0.464 x 4.64 |
9.04 |
|
rosca cuboidal |
0.215 x 0.251 x 21.54 |
21.73 |
|
lámina cuboidal |
0.215 x 2.15 x 2.15 |
11.09 |
|
lámina cuboidal |
0.0464 x 4.64 x 4.64 |
43.92 |
Consecuencias de forma y tamaño
Para ver cómo son significativas la superficie y la forma, usaremos el ejemplo del flujo de calor de un ambiente más cálido a una celda más fría, pero el mismo principio se aplicaría al flujo de calor de la celda al ambiente o al movimiento de materiales, por ejemplo, nutrientes hacia la celda o productos de desecho fuera de la celda. Debido a que los objetos más pequeños tienen una superficie relativamente mayor que los más grandes, los organismos más pequeños se calientan más rápidamente que los más grandes. De hecho, debido al efectivo intercambio de calor entre ellos y su entorno, los organismos pequeños siempre están muy cerca de la misma temperatura que su ambiente. Solo los organismos grandes, con una pequeña relación de superficie a volumen, pueden desarrollar temperaturas sustancialmente diferentes de su entorno. Considerando la forma, los cuerpos esféricos, con la menor superficie por unidad de volumen, se calientan más lentamente que cualquier otra forma cuando se ponen en un ambiente que es más caliente que él; cuanto más desviación de una forma esférica, más rápido ganará calor. Si tuvieras tres pedazos de hielo, uno esférico, uno filamentoso y otro en forma de disco, todos del mismo volumen, el disco se derretiría primero, luego el filamento y duraría la esfera. Asumiendo volúmenes iguales para cubitos de hielo, los mejores cubitos de hielo, si quieres que duren (no se derritan), son esféricos, los mejores cubitos de hielo si quieres que enfríen la bebida en la que están, son formas que más se desvían de las esferas
Una segunda influencia de la forma: la extensión del ambiente que se explora
Un hecho que a menudo se pasa por alto es que los organismos cambian su entorno a su alrededor. En el ejemplo que se acaba de dar, la transferencia de calor a la celda da como resultado un enfriamiento del entorno adyacente a la celda. El enfriamiento del ambiente junto a la celda reducirá la ganancia de calor por parte del organismo y disminuirá la importancia de la superficie para la transferencia de calor. Debido a esto, una segunda característica relacionada con la forma se vuelve importante: el volumen del ambiente que se encuentra dentro de la cierta edistancia (la distancia depende de las características de transferencia) del organismo. Para ver por qué, considera dos organismos esféricos con unas 'distorsiones' cilíndricas de su límite por lo demás esférico: uno tiene un 'outie' (una proyección que se extiende hacia afuera), el otro tiene una 'innie' (una invaginación que penetra en el organismo) (Figura 18).
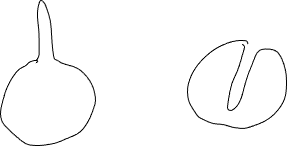
Siempre y cuando las dimensiones de la proyección/invaginación sean las mismas, ambas células tendrán la misma superficie, sin embargo, el intercambio de calor entre la célula 'outie' será más rápido que el de una celda 'innie'. Esto se debe a que las condiciones del ambiente dentro de la proyección 'innie' se volverán más similares a las del organismo que a las condiciones del ambiente a granel fuera de la célula. El volumen de 'el ambiente' que se encuentra en el 'innie' es pequeño y la superficie entre el innie y la celda es relativamente grande, en consecuencia, a medida que la parte 'innie' del ambiente pierde su calor a la celda se volvería cada vez menos significativa en una fuente de calor. Por lo tanto, la superficie adicional resultante del 'innie' sería de poca importancia en términos de interacción con el entorno. Por el contrario, la superficie adicional del 'outie' puede seguir siendo más efectiva para facilitar la transferencia de calor porque esta área está rodeada por el ambiente, no por la celda. De ahí que el área superficial por sí misma no siempre sea la mejor medida de cuánta interacción podría tener un organismo (u objeto) con su entorno.
Una consecuencia de esto es que la forma es importante para influir en la transferencia de materiales entre el organismo y el medio ambiente de dos maneras: (1) determinando el área superficial de un volumen dado de organismo, y (2) influyendo en el volumen del ambiente que está en estrecho contacto con la célula. Aunque un 'innie' sí aumenta la superficie, esto hace poco para influir en el volumen de ambiente cercano a una celda; un 'outie' hace mucho más. La importancia de la cantidad de volumen ambiental que se explora depende de varios factores, entre ellos la velocidad a la que el calor o el material se conduce a través del ambiente y la velocidad a la que el calor o el material pueden transferirse del ambiente al organismo. Si el ambiente transfiere calor o material fácilmente, o si la velocidad de transferencia a la celda es lenta, la importancia de cuánto ambiente se explora es de menor importancia.
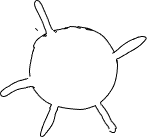

Considere otro ejemplo de dos celdas con el mismo número de múltiples salidas, extensiones hacia afuera y las mismas áreas de superficie. Uno tiene los outies juntos, el otro los tiene espaciados (Figura 19). La celda con extensiones espaciadas explora más el ambiente que la otra y podrá adquirir más calor o material de su entorno (o perder más calor o material a su entorno), especialmente si la velocidad a la que el calor o los materiales se mueven a través del ambiente es relativamente lenta, o la velocidad a la que son absorbidos/perdidos es relativamente rápida. Se puede calcular un parámetro que es el volumen de entorno a cierta distancia de la superficie celular; las outies que están cerca unas de otras son menos efectivas para aumentar la cantidad de ambiente que es 'accesible' que las salidas más separadas.
Algunas situaciones (quizás) familiares demuestran la importancia de la forma y algunas de las complicaciones relacionadas con ella. Las microvillas, pequeñas proyecciones del intestino delgado que se extienden hacia la vía intestinal, a menudo se citan como importantes en la absorción de materiales del intestino porque proporcionan mayor área de superficie. Este es ciertamente el caso pero también hay que señalar que el movimiento del material a través de la vía intestinal, resultado de la peristalsis, es lo que permite que la superficie adicional sea significativa. Si no fuera por el peristaltismo llevando continuamente material 'fresco' al denso rodal de microvillas, el aumento de la superficie sería de poca utilidad. La peristalsis cambia el ambiente junto a las microvellosidades. Los pelos radiculares, extensiones cilíndricas de las células en el exterior de las raíces, son otra situación en la que el aumento de la superficie se cita como significativo para la función de absorción de agua de las raíces. Este puede no ser siempre el caso, especialmente cuando los pelos radiculares son extremadamente densos y si el agua es abundante, lo que le permite moverse más fácilmente por el suelo. Sin embargo, las raíces y los pelos radiculares no solo absorben agua, también adquieren nutrientes y el impacto de los pelos radiculares puede ser diferente para diferentes nutrientes en comparación con el impacto para el agua. Además, la conductividad del suelo al agua y a los minerales se ve muy fuertemente afectada por la cantidad de agua presente, una propiedad muy dinámica para la mayoría de los suelos. Los pelos radiculares probablemente hacen múltiples cosas que son significativas para la absorción de agua y minerales: (1) aumentar la superficie, (2) aumentar el volumen de suelo en estrecha proximidad a la raíz, (3) mejorar el contacto entre la raíz y el suelo evitando brechas (espacios de aire) que reducirían drásticamente la absorción de agua y nutrientes, (4) realizar funciones metabólicas que facilitan la absorción de nutrientes, por ejemplo, el transporte activo.
Formas de Organismos
Si bien hay una amplia variedad de formas de organismos, tres formas comunes son cilindros, láminas y esferas. Muchos organismos son compuestos de diferentes formas, es decir, tienen algunas piezas que son una forma y otras piezas que son de otra forma, por ejemplo, m cualquier animal tiene apéndices cilíndricos unidos a un núcleo esférico. La mayoría de las plantas sobre el suelo están compuestas por láminas aplanadas (hojas) unidas a tallos cilíndricos. Tanto la forma aérea como la subterránea de las plantas suelen ser filamentos que se ramifican repetidamente, una forma que también se encuentra en los hongos. Las formas comunes para los organismos cubiertos en este texto se esbozan en el Cuadro 4.
|
forma |
ejemplos |
notas |
|---|---|---|
|
esferas |
muchos organismos unicelulares, algunos coloniales y multicelulares |
baja relación de superficie a volumen y pequeña cantidad de volumen ambiental explorado por unidad de organismo |
|
aplanado |
muchas algas coloniales y multicelulares incluyendo lechuga de mar (Ulva) y algas marinas (Laminaria); la forma haploide de todos los hornworts y muchas hepáticas |
alta relación de superficie a volumen, a menudo significativa para que los organismos fotosintéticos absorban más luz |
|
cilindros (ramificados o no ramificados) |
la mayoría de los hongos, muchas algas verdes y rojas, algunas bacterias, las raíces de las plantas vasculares |
tanto una alta relación de superficie a volumen como potencialmente una gran cantidad de volumen ambiental explorado por unidad de organismo |
|
cilindro con apéndices (no cilíndricos) |
algunas macroalgas (rojas, marrones y verdes); la mayoría de las plantas, incluyendo la mayoría de los musgos, muchas hepáticas y casi todas las plantas vasculares |
los apéndices suelen ser aplanados y fotosintéticos y su forma aumenta la cantidad de superficie expuesta a la luz |
Esferas
Como se mencionó anteriormente, las esferas tienen una superficie mínima por unidad de volumen. Suponiendo que existe un límite especializado en el exterior del objeto, ya sea unicelular o multicelular, una esfera requeriría la cantidad mínima de límite, que a menudo se compone de materiales relativamente caros. Las formas esféricas también son más móviles en muchas situaciones debido a su menor arrastre, que en general aumenta con la superficie. Si bien hay una serie de animales aproximadamente esféricos, los organismos multicelulares esféricos de otros grupos, en particular, los grupos que estamos cubriendo, son poco comunes. Sin embargo, la forma se encuentra comúnmente organismos que son unicelulares: muchas bacterias, muchas algas verdes unicelulares, dinoflagelados, criptófitas y coccolitóforos son de forma aproximadamente esférica y la forma ocasionalmente ocurre en organismos coloniales (algunas algas verdes). Las esferas también son comunes en las unidades de dispersión: polen, semillas, esporas, todas las cuales son entidades que podrían considerarse organismos. Y las formas esféricas también son una forma común para las estructuras (órganos) que contienen elementos a dispersar: esporangios (recipientes de esporas), frutos (contenedores de semillas), anteras (contenedores de polen). La (s) ventaja (s) de las formas esféricas sin duda varía según las circunstancias y también puede reflejar otras limitaciones en el desarrollo. En raras ocasiones, semillas de forma redonda, frutos e incluso plantas enteras pueden ayudar en la dispersión de propágulos por el viento y la gravedad. Un ejemplo de planta entera de esto es la tumbleweed, cuya forma esférica promueve la dispersión por el viento de las semillas que se liberan de la planta rodante. Las formas esféricas son típicas de los órganos de almacenamiento (generalmente subterráneos) de las plantas con flores: bulbos, bulbos y tubérculos y esto es probablemente una consecuencia de consideraciones de superficie a volumen.
Estructuras aplanadas
Las estructuras aplanadas son especialmente comunes en los organismos fotosintéticos, indudablemente por la importancia de interceptar la luz. Si bien a menudo se afirma que las formas aplanadas 'interceptan más luz solar' que otras formas (por ejemplo, hojas de arce vs. agujas de pino), esto es engañoso; cantidades similares de luz pueden adquirirse dada cualquier forma en particular. Lo significativo es cuánto volumen se requiere para producir un área dada de superficie absorbente de luz, y también cuánta superficie total se requiere. En el Cuadro 5 se comparan las características morfológicas de tres formas diferentes de hojas: cubos, láminas y filamentos. En cada situación se hace la suposición de que una 'cara' de la forma está orientada hacia la fuente de luz y que la absorción de luz sólo ocurre en esta superficie. Como se puede observar la misma cantidad de luz podría ser interceptada por diez hojas cuboidales, cada una de 10 x 10 x 10 cm de tamaño, o diez hojas filamentosas, cada una de 100 x 1 x 1 cm o por diez hojas planas de 10 x 10 x 1 cm. Si bien tanto los filamentos como las hojas planas pueden producir la misma cantidad de superficie absorbente por unidad de volumen de hoja, las hojas cuboidales 'cuestan' mucho más (área iluminada: volumen total) (Cuadro 5). La diferencia entre las hojas filamentosas y las hojas planas es que las hojas planas reducen la superficie total requerida para producir una cantidad dada de superficie absorbente. Probablemente son estos dos factores los que son significativos para asegurar que las superficies absorbentes de luz sean generalmente planas: las hojas gruesas requieren un volumen que no se puede usar eficazmente para la fotosíntesis; las hojas filamentosas producen un exceso de superficie total que puede ser costoso en términos de otros factores, por ejemplo, la pérdida de agua para organismos terrestres.
La estructura aplanada puede ser todo el organismo (algunas algas verdes, rojas y pardas, algunas hepáticas (incluyendo Marchantia, hornworts) o, más comúnmente, las estructuras aplanadas ocurren como hojas/estructuras similares a hojas de un tallo cilíndrico.
|
forma |
número de hojas |
dimensiones |
volumen total |
superficie total |
área iluminada/ área total |
área iluminada/ volumen |
|
cuboidal |
10 |
10 x 10 x 10 |
10000 |
6000 |
1/6 |
1:100 |
|
filamentoso |
10 |
100 x 1 x 1 |
1000 |
4020 |
1/4.02 |
1:1 |
|
planar |
10 |
10 x 10 x 1 |
1000 |
2400 |
1:2.4 |
1:1 |
Ciertamente, el tratamiento anterior es superficial y hay que señalar algunos factores complicadores: la luz generalmente no solo es absorbida por una superficie de cualquier estructura ya que la posición del sol cambia durante el día y la estación; la luz penetra a través de una superficie a capas inferiores; aunque la mayoría de las hojas se mueven solo cantidades menores durante el día o la estación, algunas hojas cambian de posición para 'rastrear' el sol; y algunas hojas cambian de posición para reducir la exposición solar. Y, a pesar de la tendencia de superficies absorbentes de luz generalmente planas, existen múltiples ejemplos de 'hojas' filamentosas y también ejemplos de 'hojas' esféricas.
Una forma plana también se encuentra a veces en los propágulos, las etapas de dispersión de los organismos, donde el 'ala' aplanada ayuda a la dispersión en el aire. Curiosamente, las alas se encuentran principalmente en propágulos relativamente grandes, las semillas y frutos de las plantas semilleras, con dimensiones mayores a unos pocos milímetros. Las 'alas' son poco frecuentes en organismos acuáticos y en pequeños propágulos: e.g., esporas y polen. Aunque la mayor parte del polen de coníferas tiene dos 'alas', no están particularmente aplanadas y, a pesar de que las coníferas son polinizadas por el viento, el papel de las alas puede no ser ayudar en la dispersión sino más bien orientar el grano de polen después de que se haya dispersado.
Estructuras cilíndricas

Las estructuras cilíndricas son extremadamente comunes, tanto como partes del organismo como como todo el organismo. Los organismos unicelulares cilíndricos se encuentran en bacterias y arqueas en forma de bastón; las formas coloniales filamentosas están representadas por algunas bacterias, muchas cianobacterias, diatomeas coloniales, muchas algas verdes y algunas algas rojas y pardas. Para la mayoría de estos organismos coloniales, los filamentos son todos de una célula de espesor, pero, especialmente en las algas rojas y pardas, los filamentos pueden ser más gruesos, a menudo varias células gruesas. Hay muchas algas verdes coenocíticas que son filamentosas.

Una estructura filamentosa ramificada es particularmente efectiva como una estructura de 'alimentación' (para hongos) o 'minera' (para raíces de plantas) que explora un ambiente para obtener recursos. Dicha forma también es efectiva para proporcionar anclaje porque permite una interacción extensa entre el organismo y su sustrato. A diferencia de las raíces, que tienen funciones duales de absorción y anclaje, los retenedores de algas y rizoides de musgos, hepáticas y hornworts realizan poca absorción pero principalmente cumplen un papel de anclaje. A través de la diversidad de organismos, lo que es lo que forma la estructura ramificada y el tamaño de la estructura ramificada varían enormemente. Los quítridos unicelulares (hongos) forman 'rizoides' para absorción y anclaje que son extensiones de una sola célula. Los 'arbusculos' (que significa literalmente 'pequeño árbol') de los hongos Glomeromycota también son extensiones de partes de la célula fúngica que ingresa a las células radiculares de las plantas. Los pelos radiculares también son extensiones de células individuales. Los rizoides de musgos, hepáticas y hornworts suelen ser un filamento formado por una cadena de células individuales. Las retenciones de macroalgas y las raíces de las plantas (a diferencia de los pelos radiculares), son multicelulares y están formadas por múltiples células.
Cilindros con apéndices no cilíndricos
Los cilindros también funcionan para posicionar órganos en ubicaciones favorables, por ejemplo, el tallo de las plantas o el estipo de las algas más grandes, distribuyendo hojas aplanadas/estructuras similares a hojas para obtener más luz. La gran mayoría de las plantas, que van desde musgos, la mayoría de las hepáticas y todas las plantas vasculares, tienen una estructura aérea que puede describirse como un 'tallo con hojas'. Por lo general, el tallo cilíndrico se ramifica y es erecto pero a veces no está ramificado y a veces está postrado, corriendo a lo largo de la superficie del suelo.
Phytopthora, un moho de agua, es un parásito que crece dentro de las hojas de las plantas y produce estructuras ramificadas que emergen de la hoja a través de estomas, poros regulados en la superficie foliar y forman estructuras cilíndricas ramificadas sobre las cuales se producen estructuras reproductivas que se dispersan.
Los cilindros (tallos) también sirven repetidamente para elevar estructuras que están produciendo propágulos (estructuras reproductivas móviles) y así facilitar la dispersión de los propágulos. La ubicuidad de la estructura refleja la significación de la estructura en múltiples situaciones en un rango de tamaño s: desde tallos tan cortos como 1 micra en mixobacterias, a 1 milímetro en mohos de limo, a 1 centímetro en musgos, a metros de altura (plantas con flores). Los tallos cilíndricos pueden ser partes de una célula (moldes de pan, algunos mohos de limo), hechos de un solo filamento de células (hongos), una colección de filamentos entrelazados (hongos) o verdaderamente multicelulares (algunos hongos, musgos, plantas vasculares). Generalmente, tales estructuras se encuentran en organismos terrestres con tallos que transportan estructuras productoras de propágulos hacia arriba hacia el aire. Al parecer, incluso una extensión ascendente muy corta puede mejorar la dispersión al permitir que los propágulos se liberen por encima de la parte más sedentaria de la capa límite, la capa de aire inmóvil que cubre todos los objetos, y en particular el suelo. Al parecer, tales estructuras no son tan ventajosas en situaciones acuáticas porque son mucho menos comunes. En algunas situaciones, un tallo es significativo no al mover la estructura productora de propágulos hacia arriba (en relación con la gravedad) sino porque mueve la estructura fuera del propio cuerpo del organismo o fuera del cuerpo del que está creciendo dentro (por ejemplo, hongos, algunos parásitos).

Como demuestra el último ejemplo, los cilindros son significativos para una serie de organismos como medio de movilidad, para llegar a nuevos lugares. Si bien la mayoría de los organismos que estudiamos son considerados inmóviles, son capaces de moverse debido al crecimiento, y un cilindro es una estructura efectiva para cubrir el territorio. Los ejemplos incluyen: que corren horizontalmente por encima del suelo tallos (estolones) y por debajo del suelo tallos (rizomas) de plantas, los 'estolones' de moldes de pan, rizomorfos de hongos, 'corredores' horizontales en Caulerpa, una alga verde.