
1.7: Desarrollo de formas productoras
- Page ID
- 56443

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Una característica de los seres vivos, y no sólo de los organismos, es que son dinámicos, cambian, y lo hacen de una manera predecible que puede describirse y observarse una y otra vez en una variedad de entidades biológicas. Este patrón de cambio a través del tiempo se denomina desarrollo y aunque el término se asocia principalmente con organismos, es significativo darse cuenta de que el cambio organizado (es decir, controlado de alguna manera) a través del tiempo se ve en las células (por ejemplo, el ciclo celular, apoptosis), poblaciones (por ejemplo, logística y exponencial crecimiento poblacional) y comunidades (e.g., sucesión).


TEMAS
- Desarrollo del organismo
- Desarrollo en organismos unicelulares
- Desarrollo de la forma celular
- Desarrollo en organismos coloniales
- Desarrollo en organismos cenocíticos/sifonáceos
- Desarrollo en organismos multicelulares
- Desarrollo en plantas
Desarrollo del organismo

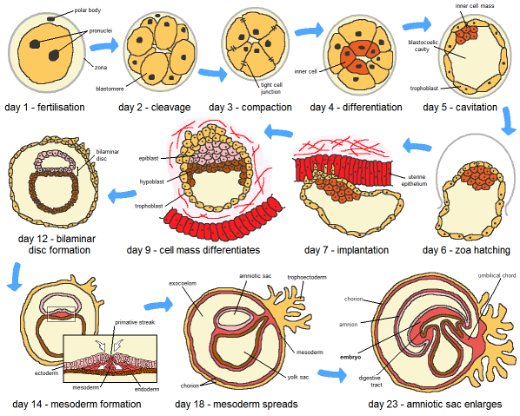
Si bien el desarrollo puede incluir cambios en una variedad de aspectos de un organismo, desde la estructura hasta la fisiología y el comportamiento, nos centraremos aquí en cómo el desarrollo produce y cambia la estructura y forma de un organismo a lo largo de su vida útil. Hemos definido a los organismos como distintos en el tiempo y el espacio con un punto de partida y un punto final; así, los aspectos de desarrollo son las alteraciones en el tamaño, la forma y la estructura que ocurren durante la vida útil del organismo. Para la mayoría de los organismos familiares el punto de partida es una célula especial que tiene la capacidad de proliferar, desarrollar una estructura compleja y crecer para producir un 'punto final', una forma específica que caracteriza a esa criatura en particular. Muchos podrían pensar que este es el fin del desarrollo, pero para muchos organismos hay cambios en el adulto con el tiempo y a veces los cambios son dramáticos y terminan con la vida del organismo. Algunos organismos se 'autodestruyen' como parte de su proceso de desarrollo, al igual que algunas células lo hacen. El salmón muere después del desove, al igual que las plantas de trigo. En ambos casos el punto final del proceso de desarrollo al es la muerte. Un corolario de esto es que el desarrollo no solo incluye el crecimiento y el aumento de la complejidad, también puede incluir, y de hecho a menudo termina con, disminuciones de tamaño y complejidad y, en última instancia, el fin del organismo.Figura 6

El campo del desarrollo ha explotado en los últimos años y se está examinando con gran detalle desde perspectivas moleculares y evolutivas. Estamos enfocados a un nivel muy diferente, considerando las características de desarrollo muy básicas que provocan la forma y estructura de las entidades biológicas que describimos como organismos. Se han descrito diversas formas de organismos: unicelulares, multicelulares simples (filamentosos, láminas), coenocíticos, multicelulares complejos. Consideraremos brevemente cómo se desarrolla cada una de estas formas. Un aspecto importante del desarrollo para muchos organismos es el proceso del sexo, que implica combinar la información genética de dos organismos y posteriormente reducirla a la mitad. El sexo y la reproducción se consideran posteriormente en el capítulo 11.
Desarrollo en organismos que son unicelulares

Varios organismos considerados aquí (por ejemplo, muchas bacterias, arqueas, diatomeas, dinoflagelados y muchas algas verdes) son unicelulares y, si bien su desarrollo es bastante diferente al de los organismos multicelulares, cambian con el tiempo y muestran patrones definidos de desarrollo. Universalmente, el desarrollo en organismos unicelulares tiene dos manifestaciones visibles (hay muchos más eventos de desarrollo que no son visibles): división celular (Figura 7) y crecimiento celular. Estos procesos son componentes del 'ciclo celular', una serie repetitiva donde las células adquieren materiales, sintetizan moléculas a partir de ellos y dividen estos materiales en dos células hijas en proceso de mitosis. Si bien la molécula más obvia que se necesita hacer es el ADN, hay una gran cantidad de otras: moléculas para membranas, ribosomas y (para células eucariotas) para mitocondrias y plastidios. La construcción de estos materiales requiere que la célula obtenga los elementos de los que están hechos: carbono, nitrógeno, fósforo, etc., y parte del ciclo celular implica la adquisición de estos materiales y la construcción de las moléculas requeridas.
La relación entre la división celular y el crecimiento celular no es la misma en todos los organismos. Las células pueden dividirse repetidamente sin ningún crecimiento (por ejemplo, en el desarrollo temprano de muchos animales). En esta situación la célula original tiene muchos 'materiales' y las divisiones celulares simplemente dividen estos materiales en células hijas dando como resultado un organismo multicelular que es básicamente del mismo tamaño que el óvulo fertilizado original. Pero esto no puede continuar para siempre, eventualmente tiene que haber adquisición de material, reflejada en un aumento de masa y tamaño (es decir, crecimiento), para permitir una división celular continua.
El componente principal de todas las células es el agua y su adquisición es generalmente lo que impulsa el crecimiento celular (es decir, la expansión en tamaño). Para los organismos con paredes celulares, la entrada de agua en la célula se ve afectada de dos maneras: al debilitar la fuerza de la pared celular o al aumentar la concentración de solutos, lo que lleva a un aumento de la presión interna. Esto se discute más en el capítulo 22, pero por ahora se debe apreciar que ambos procesos pueden ser controlados por la actividad celular. En consecuencia, la absorción w ater y el crecimiento celular pueden ocurrir como una fase distinta que ocurre antes o después de que se obtengan otros materiales, o puede ocurrir gradualmente. Es decir, la célula puede: (1) crecer de manera constante a medida que se obtienen tanto el agua como otros materiales; (2) dividir, adquirir materiales distintos del agua mientras permanecen aproximadamente del mismo tamaño, y una vez que se han obtenido suficientes materiales distintos del agua, adquieren rápidamente agua y crecen o (3) dividir, crecer rápidamente por absorbiendo agua y mostrando poco crecimiento ya que adquiere materiales distintos al agua, para luego dividirlo nuevamente. En el primer caso hay un rango de tamaños celulares y la edad de una célula se correlaciona con su tamaño.. En el segundo caso, las células que están a punto de dividirse son reconocibles por su gran tamaño. En el tercer caso, una población de células sería todas del mismo tamaño a excepción de algunas células pequeñas que acaban de producirse por división celular.
El patrón mostrado por ciertas diatomeas unicelulares ilustra posibilidades adicionales de buzos e. Todas las d iatomeas están encerradas en una cubierta de sílice con dos mitades; algunas están estructuradas como una placa de Petri con una mitad superior circular que tiene un diámetro ligeramente mayor que el de la mitad inferior, permitiendo que sea una cubierta de la parte inferior. Cuando el citosol se divide (citocinesis), la mitad superior produce una célula hija al crecer una nueva mitad inferior y la célula recién producida está 'completamente crecida' en su inicio, es decir, no muestra crecimiento. Esta célula, como todas las células, pasa por las etapas G1 y G2 y durante estas etapas 'se adquieren materiales y se juntan en biomoléculas que posteriormente se pueden dividir entre dos células hijas, pero estas fases, aunque a menudo se describen como 'crecimiento 1' y 'crecimiento 2' no implican un aumento en tamaño de celda. Consideremos ahora la segunda celda hija, la que está asociada con el plato de fondo viejo, el platillo más pequeño. Este plato inferior se convierte en un 'plato superior' ya que la celda genera una nueva mitad inferior. Al igual que su célula hermana no crece; en realidad termina un poco más pequeña que la celda padre porque la mitad inferior de la celda original se ha convertido en la mitad superior de la celda hija; en consecuencia, esta celda es un poco más pequeña que su padre (recuerde que la cubierta inferior tiene que caber dentro de la cubierta superior y por lo tanto, debe ser más pequeño). Así, después de la división celular, se producen dos células; una es del mismo tamaño que la célula original y la otra es ligeramente más pequeña. El crecimiento de células individuales no está ocurriendo. A través del tiempo, el tamaño celular medio de la población de diatomeas se hace cada vez más pequeño hasta que se alcanza algún tamaño mínimo crítico cuando se desencadena la reproducción sexual y resulta, entre otras cosas, en células que son del mismo tamaño que la original.
Exactamente lo que provoca que una célula se divida es un proceso estrechamente controlado que ha sido ampliamente estudiado debido a su conexión con el cáncer. Pero el fenómeno también es importante en otras situaciones. Por ejemplo, en las 'floraciones algales', cuando una población de alga comienza a reproducirse rápidamente, produciendo poblaciones muy grandes cuya existencia a menudo tiene consecuencias muy significativas. Tanto en las floraciones de cáncer como de algas, la importancia de los mecanismos de control, distintos de los nutrientes, está indicada por el hecho de que los nutrientes por sí solos pueden no desencadenar el crecimiento población/cáncer: los nutrientes son necesarios pero no suficientes para el crecimiento. L al control del proceso no es simplemente que las células se dividan cuando adquieren suficientes materiales para formar una segunda celda.
Desarrollo de la forma celular
Parte del proceso de desarrollo, tanto para organismos unicelulares como para las células de organismos multicelulares, implica la adquisición de una forma característica. Para algunas células, el crecimiento avanza por igual en todas las dimensiones y las células pequeñas tienen una forma muy similar a las grandes, pero para muchas células el crecimiento es decididamente diferente en diferentes direcciones y esto produce células 'adultas' con formas características, distintas de la forma típicamente esférica/cuboidal del células recién producidas. El logro de esta forma es una característica importante de su desarrollo. Para los organismos con paredes celulares, la forma de las células está determinada por la fuerza relativa de la pared celular en diferentes direcciones. Las células crecen al tener una presión interna que excede la fuerza cohesiva de la pared celular. Los rendimientos de la pared y se produce el crecimiento celular. Si la fuerza de la pared celular es uniforme entonces la celda se expande uniformemente (piense en un globo típico). Pero si la pared tiene menos resistencia en una dirección particular, entonces la expansión se producirá en esa dirección, piense en los globos especiales que se vuelven largos y flacos, esto es resultado de que el globo es mucho más resistente (mayor resistencia cohesiva) a la expansión radial que a la extensión. La fuerza de las paredes celulares viene determinada por la orientación de las microfibrillas de celulosa y esta orientación se determina a medida que se depositan.
Desarrollo en organismos coloniales
Un aspecto esencial para el desarrollo unicelular es que las células hijas se separan, resultado de que la unión entre la célula 'nueva' y la 'vieja' es débil y puede romperse por fuerzas en el ambiente o por fuerzas que acompañan la expansión de una o ambas de las células hijas. Si esto no sucede, la división celular provoca una acumulación de células que pueden afectar drásticamente el funcionamiento de cada una cambiando el entorno al que están expuestas y cambiando de forma. Los racimos de células se consideran colonias que presentan 'multicelularidad simple'. Son estructuras biológicas que no encajan muy bien en la categoría de organismo, estando en algún lugar entre un organismo y una población. A menudo las colonias tienen formas características que se desarrollan como consecuencia de patrones de división celular. Anteriormente se describieron tres patrones básicos que resultan del control del plano de división celular: filamentos, láminas y colonias esféricas . Cada uno de estos patrones tiene consecuencias para las células individuales y su interacción con el entorno. Las colonias esféricas dan como resultado algunas células (las del interior) que tienen muy poco contacto con su entorno, las láminas y filamentos representan situaciones donde la colonia de células tiene más interacción con su entorno aunque nunca tanto como sería el caso si las células se hubieran separado de sus padre. Si bien la mayoría de los organismos coloniales son indeterminados, sin un punto final específico, creciendo a tamaños que son dictados por condiciones bióticas y abióticas, existen algunas colonias que están determinadas, produciendo una colonia con un número determinado de células y generalmente con una forma específica.
Desarrollo en organismos que son coenocíticos/sifonáceos
La gran mayoría de los organismos que son grandes (un mm o más de longitud) están hechos de células que están organizadas de una manera particular y repetible para producir forma, es decir, son multicelulares y su desarrollo a partir de una sola célula implica la creación de células en lugares particulares para crear una forma. Pero hay algunos organismos 'grandes' cuya forma no es consecuencia de una acumulación de células sino que son una sola célula grande que se ha desarrollado a un tamaño considerable. Un ejemplo notable de este tipo de crecimiento es la Acetabularia, una alga verde que varía hasta varios cm de tamaño y tiene la forma de una sombrilla. La acetabularia, y la mayoría de los organismos que son coenocíticos/sifonáceos, poseen una pared celular y tienen presiones internas que se desarrollan como resultado de fuerzas osmóticas. La forma que producen tales células es el resultado del patrón de expansión celular, que, como se discutió anteriormente, es consecuencia de la resistencia relativa de la pared en diferentes direcciones. El moho del pan produce partes (estolones, rizoides, esporangióforos) que son todas excrecencias de una sola célula y deben existir controles de desarrollo que dirijan dónde y cuándo se producen las excrecencias y qué estructura se va a formar. Del mismo modo, algunas algas verdes sifonáceas (por ejemplo, Caulerpa)

son capaces de producir formas complejas y grandes mediante el control de la forma y dirección de las extensiones fuera de una sola celda (Figura 8). Algunas formas coloniales sifonáceas son parcialmente celulares en su construcción: en Hydrodictyon las células individuales son grandes (hasta un centímetro) y multinucleadas pero se unen en anillos, comúnmente de cinco o seis miembros, para formar una red poligonal.

El desarrollo en los mohos coenocíticos de limo plasmodial es particularmente dramático. En un periodo de 20 horas una célula multinucleada gigante que fluye en una red de canales y se mueve a velocidades de hasta 1 cm por hora puede transformarse en una estructura rígida delimitada por una pared celular y en forma de bosque en miniatura (~ 1-2 mm de altura) de estructuras acechadas que llevan esporangios en su cumbre (Figura 10).

Desarrollo en organismos multicelulares
El desarrollo en organismos multicelulares es un proceso mucho más complicado. La multicelularidad requiere que un organismo produzca más de un tipo de célula. Excepto en el caso inusual de los mohos celulares de limo (Dictyostelium), todas las células de organismos multicelulares tienen la misma composición genética. T su significa que hay que controlar procesos que dicten que algunas celdas sigan un conjunto de instrucciones mientras que otras celdas siguen diferentes instrucciones. Los organismos unicelulares y coloniales pueden tener la capacidad de hacerlo de manera limitada. A pesar de que la mayoría de las células de estos organismos son todas iguales, pueden producir células especializadas para afectar el proceso sexual, para llevar a cabo funciones metabólicas especializadas (por ejemplo, los akinetes de cianobacterias), o para lograr la dispersión. La multicelularidad compleja requiere una característica significativa adicional además de la capacidad de producir diferentes tipos de células: la capacidad de organizar múltiples tipos de células en un patrón tridimensional para formar tejidos y con tejidos organizados para formar órganos y con los órganos organizados para formar organismos. Esta organización requiere de un proceso de desarrollo que dicte qué tipos de células se producen, dónde se producen, y en qué medida y en qué dirección se expande cada célula.
Una distinción significativa en el desarrollo entre los animales y la mayoría de los organismos cubiertos aquí resulta de la presencia de la pared celular. El desarrollo animal incluye la posibilidad de migración celular, con el movimiento de las células permitiendo la producción de arreglos y formas particulares. Fuera del reino animal la migración celular ocurre ocasionalmente pero sólo en organismos que carecen de paredes celulares. Las células de moho de limo celular deben migrar para agregarse y posteriormente moverse entre sí, y el desarrollo de moho de limo celular implica un reordenamiento físico de las células. Cuando se produce una forma 'final' de un moho celular de limo, las celdas individuales ya no son capaces de moverse porque han desarrollado paredes celulares y están pegadas a las celdas vecinas. Casi todos los organismos multicelulares tratados en este libro poseen paredes celulares y una célula recién producida está 'pegada' a la célula que la produjo. Además, para todas las plantas, la división celular ocurre en una región llamada meristemo, una región que produce nuevas células con una organización tridimensional. Las nuevas células que se producen no solo están unidas a su célula parental, sino que también se unen a múltiples células que se están produciendo simultáneamente a su alrededor. En contraste, como se discutió anteriormente, los hongos y algunas algas rojas, verdes y pardas producen estructuras multicelulares tridimensionales de formas que no involucran un meristemo, generalmente al tener filamentos individuales (estructuras unidimensionales) pegados entre sí de manera que forman una forma tridimensional.
Desarrollo en plantas
Las plantas (aquí solían incluir musgos, hepáticas, hornworts y plantas vasculares) tienen varias características de desarrollo que no se encuentran en animales familiares:
- El crecimiento de las plantas es generalmente indeterminado, es decir, sin un criterio de valoración definido
- Las plantas poseen meristemos, regiones embrionarias, a lo largo de su vida.
- Las plantas exhiben crecimiento modular, un patrón que también se encuentra en algunos animales menos familiares (por ejemplo, hidroides, corales, esponjas), pero no en animales familiares.
- Las plantas muestran alternancia de generaciones, es decir, que existen dos formas del organismo, una haploide y una diploide. La alternancia de generaciones (Capítulo 11) también está presente en algunas algas verdes, algas pardas, algas rojas y algunos quítridos (hongos).
Los tres primeros puntos se consideran a continuación con el punto final considerado en una sección posterior del libro.
Crecimiento determinado e indeterminado y crecimiento de plantas
Con mucho, el patrón más familiar de desarrollo del organismo es el que muestran los mamíferos que crecen por un corto período de tiempo (relativo a toda la vida del organismo) y luego pasan la mayor parte de su vida como forma adulta. Este tipo de desarrollo puede denominarse 'determinado' porque la forma final está 'determinada', hay un punto final (un adulto) para el proceso de desarrollo. Si bien algunos de los organismos cubiertos en este curso, en particular muchas de las formas unicelulares, muestran este tipo de desarrollo, la mayoría de los 'no animales', y en particular las plantas y hongos, muestran patrones de desarrollo muy diferentes, uno que se describe como indeterminado, donde no hay punto final y el organismo es, en esencia, eterno. La clave de su naturaleza eterna viene del hecho de que las plantas retienen porciones de su cuerpo que son permanentemente embrionarias. En el patrón familiar de desarrollo de los mamíferos, un organismo es un embrión durante una parte de su vida y luego se transforma en un juvenil y eventualmente en un adulto. Un embrión a menudo se define como un 'organismo joven' pero lo que lo hace especial no es su edad sino el hecho de que las células son capaces de división celular, diferenciación celular y crecimiento celular. En los animales estas habilidades celulares solo se encuentran por un corto periodo de tiempo. Todo el animal tiene básicamente la misma edad y a través del tiempo se transforma de embrión a adulto. Para observar el desarrollo en animales se observa un embrión a través del tiempo. Si bien el adulto sí reemplaza a las células, en términos de desarrollo no está cambiando de forma. En contraste, las plantas siempre tienen regiones que son embrionarias y capaces de dividirse, diferenciarse y crecer; y en cualquier momento se puede ver el proceso de desarrollo al pasar de las partes embrionarias de la planta a las partes más viejas de la planta, es decir, toda la planta NO tiene la misma edad. Si plantaste una bellota hace diez años podrías considerar que la estructura que ahora ves es de 'diez años', pero la mayor parte es considerablemente más joven e incluso se pueden encontrar partes en las regiones embrionarias que nacieron 'ayer'. En cualquier momento se puede observar todo el desarrollo al proceso, desde el embrión hasta el 'adulto', moviéndose distalmente, desde las regiones embrionarias, ubicadas en las puntas de las raíces y brotes, hasta tejidos progresivamente más viejos más alejados de las puntas.
Las regiones embrionarias de las plantas se llaman meristemas. Hay dos características básicas que distinguen a las regiones embrionarias: (1) las células se están dividiendo y produciendo con ello más células, (2) las células son pequeñas, indiferenciadas y aún no comprometidas con un tipo celular final. La actividad de los meristemos puede variar estacionalmente, a menudo tiene períodos de quiescencia o latencia, pero su capacidad para producir más células que se desarrollan en una variedad de células es intrínseca a la región del meristema. Muy temprano en la vida de la planta toda la planta es embrionaria pero pronto algunas de las células se programan en desarrollo para madurar en tipos particulares de células y, junto con esto, pierden su capacidad de dividirse. En las plantas semilleras esta transformación tiene lugar dentro de la semilla y una semilla madura tiene dentro de ella una planta pequeña con dos regiones embrionarias en extremos opuestos de un eje raíz/brote muy corto. A lo largo de la vida de la planta este eje se extenderá debido a la expansión de las células producidas en los dos extremos. Los meristemas producen cada vez más células y la expansión de estas células empuja a los dos meristemas más lejos el uno del otro y alarga la raíz y el brote. Las regiones embrionarias se denominan meristemas primarios (son los primeros formados) y también meristemos apicales (se localizan en las puntas, ápices, de raíces y brotes). Los meristemos apicales radiculares se dividen para producir células que extienden el eje radicular. Los meristemos apicales de los brotes cumplen una función similar para los brotes, pero mientras producen nuevo material de brotes, también están produciendo hojas. El crecimiento a partir de meristemos apicales se llama crecimiento primario. En general, la parte más antigua de la planta se encuentra en la superficie del suelo y uno encuentra tejidos cada vez más jóvenes a medida que uno se mueve hacia las puntas de los brotes o puntas de las raíces. La región real de crecimiento generalmente se restringe a una pequeña porción, típicamente un centímetro o menos, justo debajo de las regiones meristemáticas.
El crecimiento de las plantas es modular
Además de ser indeterminado, el crecimiento de las plantas es modular, produciendo estructuras fundamentalmente autosimilares y recursivas. Hay dos módulos básicos para una planta vascular: raíces y brotes, y ambos pueden producir nuevos módulos, raíces de rama y brotes de rama, que son réplicas del eje del brote original y del eje radicular. Con el tiempo, se produce una entidad aparentemente compleja, sin embargo las reglas que rigen su construcción son muy simples: el brote puede generar ramas y éstas pueden generar más ramas y éstas pueden generar aún más ramas. Lo mismo es cierto para las raíces. El crecimiento de todos los módulos resulta de las actividades de los meristemos apicales y todo ese crecimiento se considera crecimiento primario.

Figura 11 Crecimiento modular de plantas: Las plantas crecen de manera modular con dos módulos básicos: brotes (que llevan hojas) y raíces. Ambos módulos tienen en su punta una zona meristemática (meristemo apical de brote y meristemo apical radicular). Estos módulos son capaces de producir módulos adicionales, raíces de rama y brotes de ramas y estos a su vez pueden producir más ramas, es decir, las ramas tienen ramas. Se trata de una estructura recursiva. Los meristemos originales de raíz y brote se crean en la semilla. Los brotes de rama se originan en lugares específicos, adyacentes a donde la hoja estaba o está unida al tallo. Un grupo de células meristemáticas, llamadas primordio de brote (rama), es dejado atrás por el meristema apical cada vez que produce una hoja. Estas generalmente no se desarrollan de inmediato y aunque se produce una rama potencial en cada unión foliar, muchas de ellas nunca se desarrollan. La producción de raíces de rama no está tan rígidamente determinada. El crecimiento producido por los meristemos apicales se considera 'crecimiento primario' ya sea que ocurra en la raíz original o en el brote o en las raíces de las ramas o brotes.
Desarrollo foliar: A diferencia de las raíces y brotes, cuyo crecimiento es indeterminado, las hojas son determinadas, produciendo una forma definida, producto de una estructura con un proceso de desarrollo definido (determinado). El meristemo apical del brote produce montículos de tejido embrionario, llamados primordios foliares, que se dividen por un periodo de tiempo, crecen por un periodo de tiempo y luego existen en forma estática hasta que se desencadena el proceso de senescencia que culmina con la hoja desprendiéndose del resto de la planta.
Origen de nuevos módulos. Para crear un nuevo módulo, se necesita crear un nuevo meristemo apical. Estos se originan de manera diferente en raíces y brotes. En las raíces, se forman nuevos meristemas apicales como resultado de la actividad de ciertas células en la capa más externa del núcleo central del tejido vascular radicular (más detalles en el siguiente capítulo). Algunas de estas células son estimuladas para que comiencen a dividirse y se organicen para formar un meristemo apical radicular. El crecimiento de las células producidas por este meristemo empuja la raíz de la rama meristema apical fuera de la raíz original y hacia el suelo, donde continúa creciendo de manera similar a la raíz original.
Los brotes de rama se crean de manera diferente y se originan a partir de un meristema apical de brotes. Además de extender el brote y producir hojas embrionarias, el meristemo apical del brote también produce un nuevo meristema apical de brote en la 'axila' de cada hoja. Estos son nuevos meristemos apicales de brotes y se llaman primordios de cogollos. Se producen en estado latente y necesitan ser estimuladas para comenzar a dividirse y producir nuevas células que permitan alargar la rama. La mayoría de los primordios de cogollos nunca son estimulados para crecer; si todos fueran estimulados, resultaría una estructura muy 'ramificada', produciéndose una rama en cada posición donde hubiera o haya una hoja. A menudo, pero ciertamente no siempre, los brotes de rama solo se estimulan para que crezcan después de que la hoja a la que están adyacentes se haya caído de la planta.
Las plantas también son capaces de producir nuevos módulos de manera no estándar. Las raíces y los brotes así producidos se denominan 'adventicios'. A veces las raíces se producen a partir de los tejidos del tallo y éstas se describirían como raíces adventicias porque todas las raíces “estándar” se forman a partir de raíces existentes. De igual manera, a veces los tejidos radiculares pueden desarrollar brotes; obviamente estos no se originan en la axila de una hoja, por lo que estos serían denominados brotes adventicios. La capacidad de algunas plantas para producir raíces o brotes adventicios permite la propagación vegetativa de las plantas: si los tallos desprendidos pueden ser estimulados para producir raíces entonces se pueden propagar plantas a partir de 'esquejes'; de manera similar, las raíces se pueden utilizar para propagar plantas. Y para algunas plantas se puede estimular el tejido foliar para producir raíces y brotes adventicios; así, a veces es posible la propagación a partir de las hojas. En todas estas situaciones, ciertas células, llamadas células del parénquima, son estimuladas para comenzar a dividirse y organizarse para formar un meristema apical. Una vez que se inicia un módulo (una raíz o un brote), se pueden formar nuevos módulos a partir de él. Una característica única de las células del parénquima es su capacidad para 'desdiferenciarse' y reanudar una condición embrionaria, capaz de dividirse y producir una variedad de tipos celulares, incluso después de que se hayan diferenciado para convertirse en células del parénquima.
Los tipos celulares de las plantas vasculares, junto con la anatomía de las raíces, tallos y hojas serán discutidos con mayor detalle en el próximo capítulo.