
1.13: Sexo y reproducción en plantas no semilleras
- Page ID
- 56425

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Casi todas las plantas son capaces de reproducirse sin sexo. Más comúnmente esto sucede como consecuencia de que la planta se corta en pedazos y estas piezas son capaces de regenerar las partes que se perdieron. Adicionalmente, algunas especies han desarrollado piezas que son particularmente propensas a romperse, por ejemplo, las gemmas (singular de gemma) de algunos musgos y hepáticas. Es relativamente común que las plantas con flores produzcan propágulos asexuales en los lugares donde normalmente se desarrollan las semillas, produciendo estructuras similares a semillas sin el proceso normal del sexo. Si bien la reproducción sin sexo es común en las plantas y para algunas especies representa su único medio de reproducción, la mayoría de las plantas dependen de la reproducción sexual, y dos innovaciones, la semilla y la flor, han sido particularmente significativas para la evolución de las plantas.
Con respecto al sexo, el reino vegetal ilustra maravillosos patrones de unidad y diversificación. Por un lado, toda la vida vegetal muestra un patrón de sexo fundamentalmente similar, por otro lado, las plantas ilustran una notable diversificación de este tema común.
TEMAS
- Alternancia de generaciones
- Musgos, hepáticas y hornworts
- Helechos
- Colas de caballo
- Clubmusses
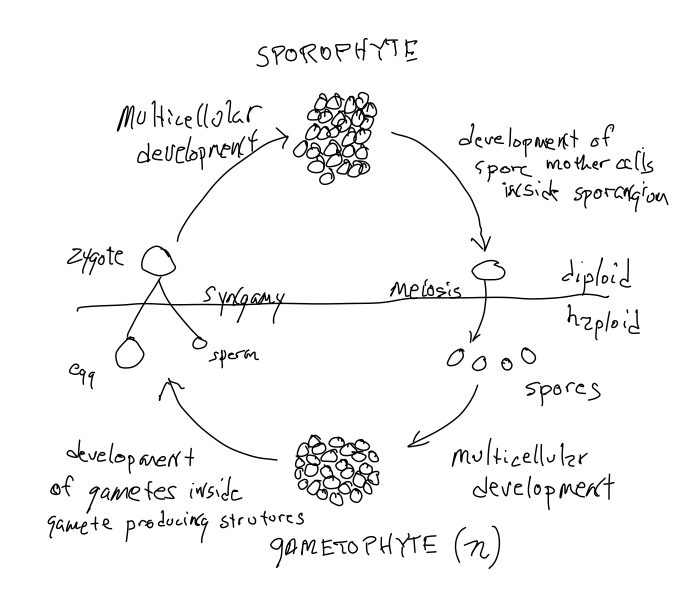
Todas las plantas presentan alternancia de generación s, producen dos tipos de organismos multicelulares: uno diploide y derivado del desarrollo de un cigoto, y otro haploide y derivado del desarrollo de una célula haploide llamada espora. El 'desarrollo' implica la proliferación de células, la diferenciación de células y la formación de estructuras específicas con roles específicos, es decir, la morfogénesis.
Todas las plantas presentan alternancia de generaciones y alternan entre organismos haploides y diploides. Para completar el ciclo sexual el organismo haploide debe producir gametos que se unan para formar el cigoto. Es decir, entre todas las células haploides que se producen como resultado de las divisiones celulares de una espora haploide, algunas de las células están dotadas de capacidades especiales que les permiten encontrarse e interactuar con otro gameto para formar un cigoto. De igual manera, de todas las células diploides derivadas del cigoto, algunas, llamadas 'células madre de esporas', están dotadas de la capacidad de sufrir meiosis para producir células haploides, esporas, que crecen en individuos haploides. Tenga en cuenta que la meiosis NO produce gametos, sino que produce esporas que se dispersan y se desarrollan en plantas haploides. La planta diploide que produce células madre de esporas y esporas se llama 'esporofita'. El organismo productor de gametos, que es haploide, se llama gametofito. Así, existe una 'alternancia de generaciones' con una forma haploide alternando con una forma diploide (Figura 1). A menudo, una de las dos formas es difícil de apreciar, en gran parte porque a menudo son difíciles de ver, a menudo son pequeñas, efímeras y pueden no vivir una existencia separada de la generación anterior. Es decir, las formas haploides y diploides pueden no ser espacialmente distintas entre sí; ¡pueden ser temporalmente, o permanentemente, una parte de su padre!
Al igual que los de animales familiares, los gametos de todas las plantas son de dos tipos, un óvulo más grande e inmóvil y un espermatozoide más pequeño que de alguna manera debe ser móvil para encontrarse con el óvulo. Los gametofitos de las plantas pueden ser bisexuales (hermafroditas), produciendo tanto óvulo como esperma, o unisexuales, es decir, hay gametofitos masculinos y femeninos separados. En algunos grupos de plantas, incluyendo todos los discutidos en este capítulo, los espermatozoides son móviles porque poseen flagelos. Los espermatozoides nadan lejos del gametofito masculino que lo produjo y son atraídos químicamente por la estructura que produce óvulos. Debido a que las plantas son organismos terrestres, el agua requerida para que un espermatozoide flagelado nade hasta el óvulo no siempre está presente. Veremos en el próximo capítulo que algunas plantas han eludido este problema al hacer que los gametofitos masculinos se vuelvan móviles, no los espermatozoides que producen.
Al igual que en los animales, el gameto femenino, el huevo, no se mueve. Se retiene en el organismo que lo produce. En el caso de las plantas se trata de un gametofito haploide (no un individualas diploides en mamíferos). Y en algunas de las plantas discutidas en este capítulo el cigoto formado por la fertilización de un óvulo crece a partir del archegonio (la estructura en la que se produce un óvulo) y después de la fertilización produce un esporófito adherido al gametofito. Alternativamente, todo el gametofito (femenino) de algunos grupos de plantas está incrustado en la planta diploide (el esporófito) que produjo la espora que se desarrolló en el gametofito femenino. Es decir, en algunos grupos de plantas, las esporas producidas por los esporófitos no se dispersan, en cambio la espora germina donde se produce y crece para producir una planta de gametofitos sobre o dentro de una planta de esporófitos. Los gametofitos masculinos móviles y los gametofitos femeninos sedentarios son características de algunas plantas discutidas en este capítulo pero se desarrollan de manera mucho más significativa en plantas semilleras, discutidas en el siguiente capítulo. En este capítulo consideramos la alternancia de generaciones más fácilmente observable y comprensible que se encuentra en plantas sin semillas.
Musgos Hornworts y Hornworts

En musgos, hepáticas y hornworts (los tres grupos de plantas que carecen de tejido vascular) es la planta del gametofito la que vive más tiempo y es la más visible. Produce una forma capaz de recolectar materiales para el crecimiento, principalmente a través de la fotosíntesis (Figura 2). La forma más común, que se encuentra en todos los musgos y muchas hepáticas, es un eje alargado que lleva colgajos de tejido que aumentan el área fotosintética. En algún momento, muchas veces una vez al año, el gametofito produce estructuras productoras de gametos que se llaman archegonia (singular = archegonio) si producen óvulos y anteridios (singular = antheridium) si producen esperma. Estas estructuras se producen típicamente en medio de 'hojas' modificadas en los extremos de los tallos/ramas en musgos o, en algunas hepáticas, ocurren en la parte inferior de estructuras en forma de paraguas/hongos que se extienden hacia arriba desde el cuerpo principal de las hepáticas taloides. Los anteridios (Figura 3) son recipientes aproximadamente esféricos en los que las células se diferencian en espermatozoides flagelados y se liberan cuando los anteridios se abren. Las archegonia son estructuras en forma de matraz con un solo óvulo cerca de la base y un cuello alargado que desarrolla un canal a través del cual los espermatozoides pueden nadar para fertilizar el óvulo. El cigoto resultante se desarrolla dentro del archegonio, produciendo un esporófito que finalmente se extiende típicamente de uno a seis cm más allá del archegonio y los dexistas como apéndice del gametofito, nunca viviendo una existencia independiente de él. Aunque el esporofito a veces es verde y capaz de fotosíntesis durante parte de su vida, su estructura, un simple tallo sin 'hojas' para aumentar la superficie, no es particularmente adecuada para la fotosíntesis. Más bien la estructura es adecuada para la dispersión de esporas; el tallo suele elevar la cápsula, que es el sitio donde se desarrollan las células madre de esporas, experimentan meiosis y producen esporas.


Si la 'separación espacial' se emplea rigurosamente para definir un organismo entonces estos esporofitos no serían considerados un organismo separado, simplemente son una parte del gametofito (Figura 5). Pero, a la luz del ciclo sexual y el patrón de alternancia de generaciones, es útil considerarlos como organismos separados. Esto se sustenta en el hecho de que tienen un número diferente de cromosomas que el gametofito del que crecen.

En todas las demás plantas además de musgos, hepáticas y hornworts es la planta productora de esporas la que vive más tiempo y es la más visible; es la forma que vemos y reconocemos como planta. El esporófito aún comienza su desarrollo creciendo a partir del archegonio donde se produjo el óvulo y fue fertilizado, pero el crecimiento del esporófito es tal que se vuelve completamente independiente del gametofito del que emerge y finalmente tiene una existencia completamente autónoma. Así, cuando vemos helechos, colas de caballo, musgos y semillas, lo que estamos observando es una planta diploide que produce esporas. En todos estos grupos el gametofito es pequeño y esquivo pero el ciclo vital básico es el mismo que en todas las plantas: una alternancia de generaciones entre un gametofito y un esporófito. Las semillas y las plantas semilleras se discuten en el siguiente capítulo, a continuación se consideran aspectos del ciclo sexual, en particular características de los gametofitos menos comúnmente vistos que son producidos por plantas vasculares sin semillas.
Helechos
La mayoría de los helechos tienen un gametofito pequeño y fotosintético que suele tener menos de 1 cm de ancho y una célula de espesor, es decir, una lámina. Carece de un eje de tallo y a menudo tiene 'forma de corazón' (Figura 6-8 y 11). Generalmente se une a un sustrato a través de rizoides (filamentos de células no fotosintéticas).

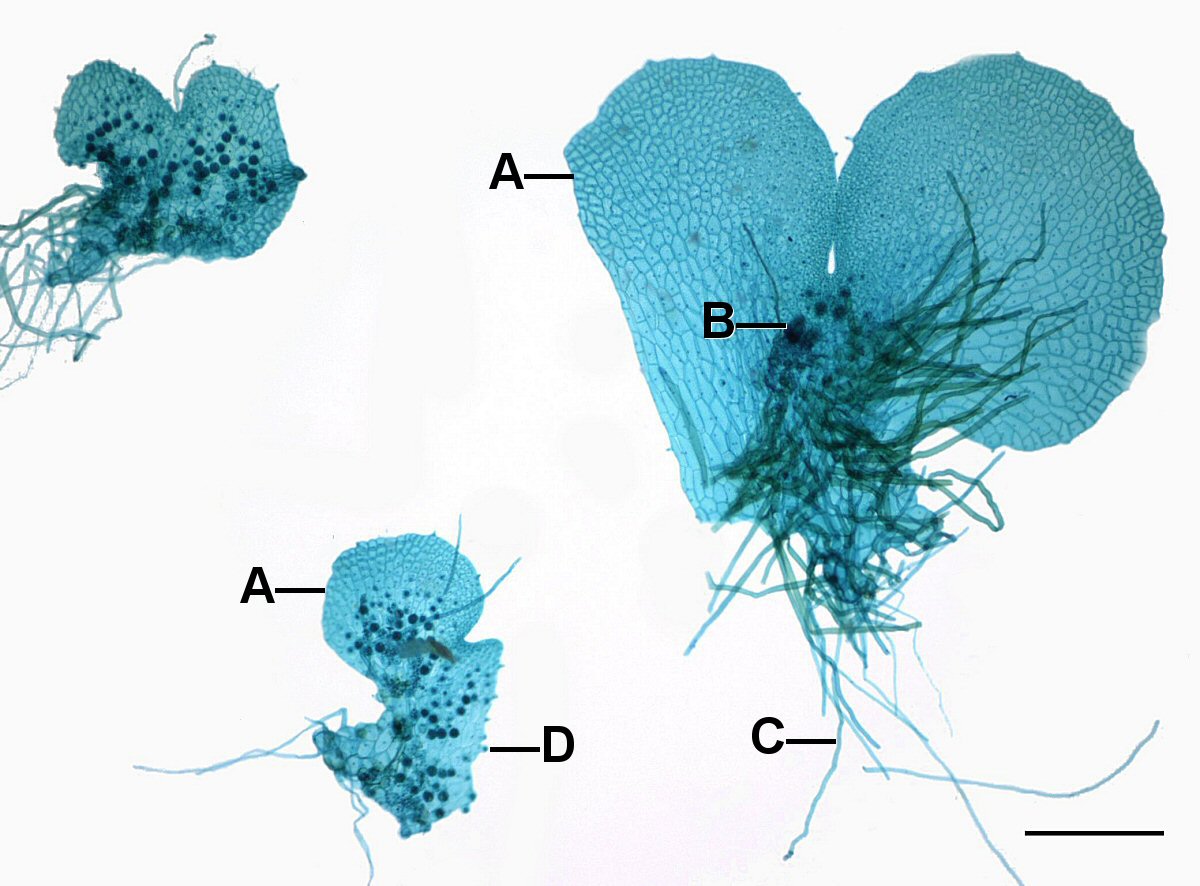
Como fue el caso en los musgos, los gametofitos de helecho producen estructuras donde el óvulo y el esperma se producen como resultado de que las células se dividen en un patrón particular para producir archegonia (Figura 7-9) y anteridios. Es importante señalar que los gametos no son producidos por la meiosis porque todas las células del gametofito ya son haploides.



Los gametofitos de helecho generalmente tienen estructuras femeninas en forma de matraz (archegonia) ubicadas en la muesca entre los lóbulos y estructuras masculinas globulares (anteridios) ubicadas en los lóbulos. Si bien la mayoría de los gametofitos de helechos son hermafroditas, algunos son unisexuales y para algunos su expresión sexual depende de las condiciones ambientales. Todas las células del gametofito son haploides pero produce una célula, el óvulo, con habilidades especiales de desarrollo. Los anteridios liberan espermatozoides que tienen flagelos que les permiten nadar hasta la archegonia, donde se encuentran los óvulos, nadan por un canal estrecho y se fusionan con el óvulo en la base. El cigoto se convierte en esporófito, produciendo tallos y raíces. Los tallos producen hojas que dan sombra al gametofito y pronto muere (Figura 9)

El esporófito sigue creciendo para producir el helecho que reconocemos. Tiene la misma estructura que la mayoría de las plantas: un eje raiz-brote con hojas producidas por el brote. La mayoría de los helechos en esta zona tienen tallos (rizomas) que están bajo tierra y relativamente cortos. Lo que vemos son las hojas que emergen sobre el suelo de este rizoma. En algún momento este organismo diploide produce estructuras denominadas esporangios, dentro de las cuales se encuentran células madre de esporas que se someten a meiosis para producir un grupo de cuatro esporas haploides (una tétrada) que son liberadas al ambiente. Cuando estos germinan, se convierten en gametofitos haploides y el proceso se repite. Generalmente, los esporangios se producen en racimos llamados 'puntos de fruto' que se localizan en la parte inferior de las hojas (ver helecho maderero). Otros helechos tienen porciones enteras de sus hojas que obviamente son diferentes y donde se producen esporas. Algunos helechos en esta zona son dimórficos (ver helecho sensible), produciendo dos tipos de hojas, algunas que son verdes y fotosintéticas y que nunca producen esporas y otras hojas que no son fotosintéticas y producen abundantes esporas, al tiempo que se nutren de la parte fotosintética de la planta.
Si bien este es el patrón general para los helechos, existe alguna variación, un ejemplo de lo cual es el helecho acuático Marsilea, que tiene varias características interesantes (ver la ficha informativa sobre Marsilea). Como es el caso en una serie de helechos, las esporas se producen en una hoja especializada que es muy diferente al aspecto de las hojas fotosintéticas normales. Mientras que las hojas normales son verdes y tienen forma de hojas de trébol, las hojas portadoras de esporas se empaquetan inicialmente en una estructura similar a una semilla, duras por fuera y capaces de ser secadas y revitalizadas (germinadas) cuando se vuelven a humedecer. En este momento la hoja 'fructífera' (es decir, portadora de esporas) emerge al agua, pareciéndose muy poco a una hoja: es sin clorofila, muy pequeña y gelatinosa. Produce esporas en racimos y hay dos tipos de esporas, esporas masculinas llamadas microsporas y esporas femeninas llamadas megasporas, cada una en esporangios separados. El nombre técnico de las plantas que producen dos tipos de esporas es heterosporosas. Por el contrario, la mayoría de los helechos son homosporosos, produciendo solo un tipo de espora que generalmente produce gametofitos hermafroditos (bisexuales) (descritos anteriormente); algunas formas homosporosasproducen gametofitos unisexuales, tanto masculinos como femeninos, pero ambos provenientes de esporas de aspecto idéntico.
Los dos tipos de esporas de Marsilea se distinguen fácilmente por su tamaño. Las megasporas tienen alrededor de 1 mm de longitud y germinan para producir gametofitos femeninos productores de huevo. Las microsporas tienen solo ~ 70 um de longitud y producen gametofitos masculinos productores de esperma. Sin ningún aumento de tamaño y sin emerger de la microsporas, las esporas masculinas germinan para producir una 'planta' con ~35 células, 32 de ellas son espermatazoides, espermatozoides en forma de sacacorchos multiflagelados, de aproximadamente 10 um de tamaño, que se liberan del gametofito masculino cuando se rompe la pared de la microespora. Los espermatazoides son atraídos químicamente hacia el gametofito femenino. El gametofito femenino es sustancialmente más grande que el gametofito masculino pero aún es pequeño y, al igual que el gametofito masculino, exhibe desarrollo endosporico, su desarrollo ocurre dentro del caso de esporas de la megaspora, con solo el cuello muy corto extendiéndose desde ella. Produce un solo archegonio con un solo óvulo en el que el esperma nada y fertiliza formando un cigoto. Si bien la nueva planta esporofita aparentemente brota de la espora hembra, en realidad proviene de un gametofito femenino que está creciendo dentro del caso de esporas.
Lectura adicional
“Marsilea: hábitat, características externas y reproducción” en Biología Discussion.com
Otro helecho interesante es el helecho cerda de los Apalaches, que sólo se conoce por la forma del gametofito. Al parecer, ¡se ha estado reproduciendo asexualmente durante millones de años! Hay varias otras especies de helechos conocidos sólo como gametofitos.

Colas de caballo
El patrón básico encontrado en helechos, con una generación dominante de esporófitos y una generación diminuta de gametofitos, se encuentra en las colas de caballo, un grupo de plantas vasculares que se originaron en el Paleozoico y produjeron una serie de formas arbóreas que fueron significativas en la producción de depósitos extensos que se convirtieron en carbón y aceite. Solo queda un género de colas de caballo y hay menos de 20 especies en todo el mundo. Todas son herbáceas con rizomas perennes que envían ramas verticales que tienen un patrón de crecimiento muy distintivo con tallos fotosintéticos, hojas muy pequeñas en forma de escamas y ramas verticiladas o sin ramas. Las esporas se producen en una estructura terminal en forma de cono, que es un grupo de esporangios. Las esporas son dispersadas por el viento pero su movimiento y liberación del esporangio pueden ser ayudados por estructuras llamadas elators, apéndices en forma de correa en la espora que se mueven en respuesta a la absorción y pérdida de agua. La germinación de la espora produce un gametofito fotosintético pequeño (~ 1-3 cm) que parece un cojín de alfiler. Los 'tallos' muy cortos están presentes con apéndices que aumentan el área fotosintética. La anteridia y la archegonia suelen producirse ambas a partir del mismo gametofito, aunque puede ser unisexual por un periodo de tiempo. Los espermatozoides son multi-flagelados y necesitan nadar para llegar al óvulo. La fertilización da como resultado un cigoto que se convierte en un esporófito diploide que pronto sobrecrece el gametofito del que emerge, produciendo raíces y tallos tanto horizontales (rizomas) como verticales.
Clubmusses, Spikemosses y Quillworts
Se cree que los tres grupos están estrechamente relacionados y se agrupan como 'Licópodos' o 'Licófitos', un grupo que también incluye una serie de formas fósiles, incluyendo formas arbóreas que fueron muy importantes al final de la era Paleozoica. El grupo tiene 1200 especies y es considerablemente más diverso que las colas de caballo pero mucho menos diverso que los helechos (12, 000 especies). Los esporofitos de musgos clubmusses existentes, musgos espigas y quillworts son todos herbáceas perennes. Generalmente se propagan extensamente con tallos sobre el suelo y debajo del suelo (los miembros tropicales suelen ser epífitas). Aunque los antiguos miembros del grupo exhibieron crecimiento leñoso, ninguna de las especies vivas hoy en día sí lo hace.
Los musgos (Figura 13) son únicos en tener gametofitos subterráneos y no fotosintéticos, sobreviviendo como parásitos al alimentarse de hongos. Estos gametofitos viven mucho más tiempo que la mayoría de los gametofitos de plantas vasculares, algunos más de 15 años. Las espikemosas y quillworts son heterosporosas y, al igual que el helecho acuático Marsilea, los gametofitos se desarrollan endoesporicamente; viviendo del material que fue abastecido en la esporita por la planta esporofita. El gametofito masculino es de muy corta duración y tiene poco material almacenado (la microsporas es pequeña), pero el gametofito femenino es considerablemente más grande y vive meses sobre el material presente en la espora. El crecimiento temprano del esporófito, a partir del gametofito femenino que está presente en una megaspora, parece una semilla en germinación (Figura 14) y veremos en el próximo capítulo que estas son analogías —no se piensa que las plantas semilleras se desarrollaron a partir de los licópodos. Otra característica de algunas espikemósesis es que las esporas 'femeninas', las que se convierten en gametofitos femeninos, a veces se desarrollan EN la planta de esporófitos, otra característica que se repite en las plantas semilleras.

