16.7: Espacio de fase de múltiples recursos
- Page ID
- 53309

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Como siguiente paso, considere el caso de dos recursos esenciales. Esto se puede hacer matemáticamente siguiendo el enfoque que usamos anteriormente para un solo recurso.
Llame a los dos recursos\(R_A\) y\(R_B\). Supongamos que del total de estos dos recursos utilizados por la Especie 1, una proporción\(p_1\) es el Recurso A y por lo tanto una proporción\(q_1\,=\,1−p_1\) es el Recurso B. Asimismo, para la Especie 2 una proporción\(p_2\) es el Recurso A y\(q_2\,=\,1−p_2\) es el Recurso B.
Esto es confuso, así que para un ejemplo clarificante, supongamos que\(R_A\) es fosfato, PO 4, y\(R_B\) es silicato, SiO 2, ambos esenciales para dos especies de algas en una vía fluvial. Tomar la Especie 1 para ser una especie de Asterionella y la Especie 2 para ser una especie de Cyclotella, como en un estudio pionero de David Tilman (1977). En este caso, la Especie 1 necesita recursos de fosfato y silicato en una proporción de aproximadamente 1:99, mientras que la Especie 2 los necesita en una proporción de aproximadamente 6:94. Si el silicato es bajo, la Especie 1 sufrirá así primero, ya que necesita una mayor proporción del mismo, mientras que la Especie 2 sufrirá primero si el fosfato es bajo, por la razón relacionada. Aquí sería\(p_1\,=\,0.01,\,q_1\,=\,1−p_1\,=\,0.99\) para el uso de los Recursos A y B por Especie 1, y\(p_2\,=\,0.06,\,q_1\,=\,1−p_1\,=\,0.94\) para uso por Especie 2.
\(u_1\)Al ser la cantidad total de recurso amarrada por cada individuo de la Especie 1,\(p_1u_1\) será la cantidad de Recurso A atada por la Especie 1 y\(q_1u_1\) la cantidad de Recurso B atada de la misma manera. De igual manera,\(p_2u_2\) será la cantidad de Recurso A amarrada por cada individuo de\(q_2u_2\) la Especie 2 y la cantidad de Recurso B. Asimismo, supongamos como antes que los recursos en consideración desaparezcan del ambiente cuando sean tomados por individuos recién nacidos, sean liberados inmediatamente cuando los individuos mueren.
Con esto en mente, los recursos que queden en cualquier momento, como funciones del recurso máximo y la abundancia de cada especie, serán
\[R_A\,=\,R_{Amax}\,-\,p_1u_1N_1\,-\,p_2u_2N_2\]
\[R_A\,=\,R_{Bmax}\,-\,q_1u_1N_1\,-\,q_2u_2N_2\]
Supongamos que las poblaciones de las Especies 1 y 2 crecen en base a qué recurso está más cercano al que\(R^{\ast}\) experimenta esa especie para el recurso. Esto puede ser representado por la función “min”, min (\(a,b\)), que selecciona el menor de dos valores. Por ejemplo, min (200,10) = 10, min (−200,10) = −200. Ahora las ecuaciones de crecimiento de dos especies y dos recursos, que generalizan el crecimiento de una sola especie y un solo recurso en la Ecuación 16.1.1, son
\(\frac{1}{N_1}\frac{dN_1}{dt}\,=\,m_1\,min(R_A\,-\,R_{1A}^{\ast},\,R_B\,-\,R_{1B}^{\ast})\)
\(\frac{1}{N_2}\frac{dN_2}{dt}\,=\,m_2\,min(R_A\,-\,R_{2A}^{\ast},\,R_B\,-\,R_{2B}^{\ast})\)
Esto podría refinarse, para que las tasas de crecimiento\(m_1\) y\(m_2\) dependieran de qué recurso estuviera limitando, pero esto no importa en el presente análisis. Si las especies son lo suficientemente similares y el nivel de recursos es tal que están limitadas por el mismo recurso, una tenderá a ser excluida competitivamente, como en el apartado anterior. Pero si las dos especies son bastante diferentes, pueden estar limitadas por diferentes recursos y las ecuaciones pueden simplificarse.
\[\frac{1}{N_1}\frac{dN_1}{dt}\,=\,m_1(R_A\,-\,R_{1A}^{\ast})\]
\[\frac{1}{N_2}\frac{dN_2}{dt}\,=\,m_2(R_B\,-\,R_{2B}^{\ast})\]
Algún álgebra revelará las propiedades básicas. Si sustituyes las expresiones para\(R_1\) y\(R_2\) de las Ecuaciones 16.6.1 y 16.6.2 en las Ecuaciones 16.6.3 y 16.6.4 obtendrás
\[\frac{1}{N_1}\frac{dN_1}{dt}\,=\,m_1(R_{Amax}\,-\,p_1u_1N_1\,-\,p_2U_2N_2\,-\,R_{1A}^{\ast})\]
\[\frac{1}{N_2}\frac{dN_2}{dt}\,=\,m_2(R_{Bmax}\,-\,q_1u_1N_1\,-\,q_2u_2N_2\,-\,R_{2B}^{\ast})\]
Ahora bien, si expandes, recolectas y reordenas términos, obtienes este formulario equivalente:
\[\frac{1}{N_1}\frac{dN_1}{dt}\,=\,m_1(R_{Amax}\,-\,R_{1A}^{\ast})\,-\,m_1p_1u_1N_1\,-\,m_1p_2u_2N_2\]
\[\frac{1}{N_2}\frac{dN_2}{dt}\,=\,m_2(R_{Bmax}\,-\,R_{2B}^{\ast})\,-\,m_2q_2u_2N_2\,-\,m_2q_1u_1N_1\]
Observe que, nuevamente, un modelo mecanicista con parámetros medibles es solo el modelo general de RSN ecológico disfrazado.
La formulación RSN puede exponer las posibilidades de situaciones de dos recursos en el espacio de fases. Las isoclinas son las Ecuaciones 16.5.5 y 16.5.6 de este capítulo, con pendientes −\(s_{1,1}/s_{1,2}\) y −\(s_{2,1}/s_{2,2}\) para las Especies 1 y 2, respectivamente.
Estas dos pendientes se pueden escribir en términos del recurso. Con los valores\(s_{i,j}\) de las Ecuaciones 16.6.7 y 16.6.8\((s_{1,1}\,=\,−m_1p_1u_1,\,s_{1,2}\,=\,−m_1p_2u_2,\,s_{2,1}\,=\,−m_2q_1u_1,\) y\(s_{2,2}\,=\,−m_2q_2u_2\)), las pendientes de las isoclinas se convierten
\(\frac{s_{1,1}}{s_{1,2}}\,=\,\frac{-m_1p_1u_1}{-m_1p_2u_2}\,=\,\frac{u_1}{u_2}\frac{p_1}{p_2}\)
\(\frac{s_{1,1}}{s_{1,2}}\,=\,\frac{-m_1p_1u_1}{-m_1p_2u_2}\,=\,\frac{u_1}{u_2}\frac{1-p_1}{1-p_2}\)
Primero note que si las dos especies utilizan los dos recursos en proporciones iguales (si\(p_1\,=\,p_2\)), ambas pendientes se vuelven\(u_1/u_2\). Las pendientes son paralelas, como en el caso de un solo recurso en las Figuras 16.5.1 a 16.5.4. Por lo tanto, si dos especies utilizan dos recursos diferentes de manera idéntica, es decir, en proporciones iguales, no coexisten. La convivencia requiere cierta diferencia en la forma en que utilizan los recursos.
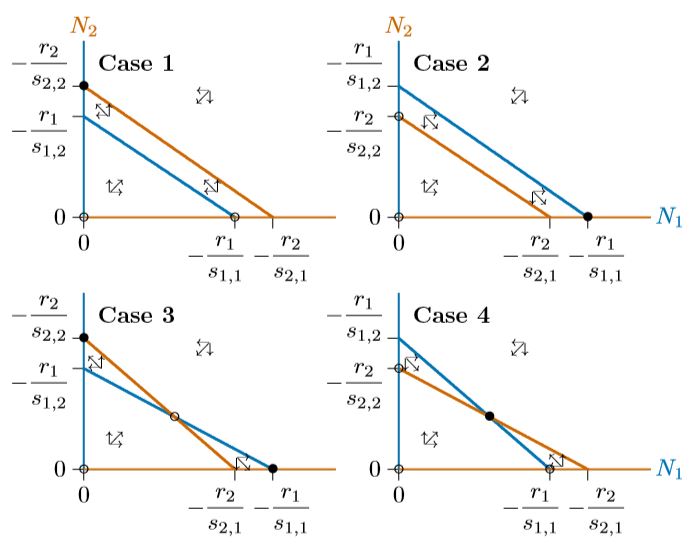
Sin embargo, utilizar los recursos de manera diferente no garantiza la convivencia. Dependiendo\(p\) de sus's y\(q\)'s, una isoclina aún podría encerrar completamente a la otra. La figura\(\PageIndex{1}\) tiene las mismas propiedades que la figura 16.5.4. En todas partes por debajo de la isoclina roja, la Especie 2 aumentará, incluyendo la banda ancha entre las isoclinas rojas y azules donde la Especie 1 disminuirá.
Si se invierten las isoclinas azul y roja, el resultado es similar, pero con la Especie 1 excluyendo la Especie 2. La figura\(\PageIndex{2}\) muestra esto, con las flechas invertidas a medida que la Especie 1 aumenta en todas partes por debajo de la isoclina azul, incluyendo la banda ancha entre las isoclinas donde la Especie 2 disminuye.
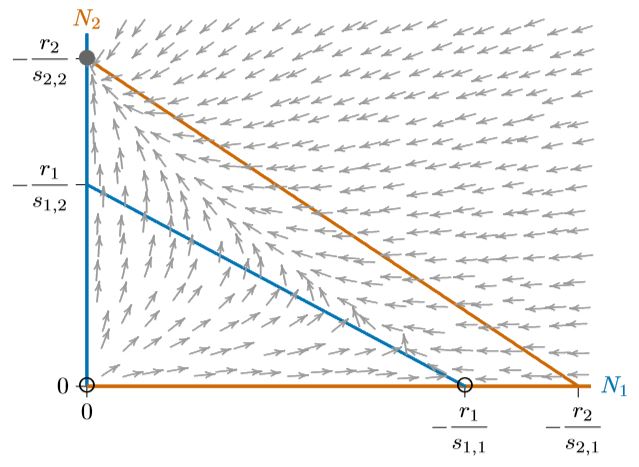
En los tres casos, de las Figuras 16.5.4 a\(\PageIndex{2}\), el sistema tiene tres equilibrios—en el origen (0,0), donde ambas especies están ausentes, en la capacidad de carga\(K_1\) para la Especie 1 sola (−\(r_1/s_{1,1}\) ,0), y en la capacidad de carga\(K_2\) para la Especie 2 sola (0,\(−r_2/s_{2,2}\)). El origen es inestable y sólo uno de los otros dos equilibrios es estable, dependiendo de qué isoclina encierra al otro.
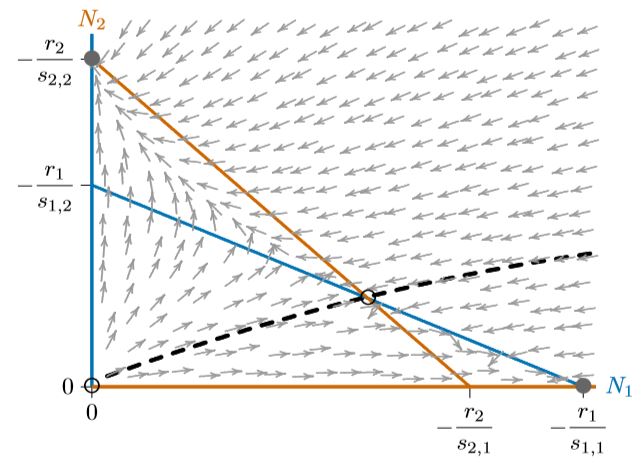
Figuras\(\PageIndex{1}\) y se\(\PageIndex{2}\) pueden combinar para dar a cada especie la oportunidad de excluir a la otra, dependiendo de las circunstancias. Esto significa no permitir que una isoclina encierra completamente a la otra, como en la Figura\(\PageIndex{3}\). Las isoclinas que se cruzan introducen un cuarto equilibrio en el interior del espacio de fase. Este equilibrio es inestable, marcado con un círculo abierto, y los dos equilibrios para las especies individuales, en los ejes, son estables. Sin embargo, ya no son “estables globalmente”, ya que solo las regiones locales del espacio de fase conducen a cualquiera de ellas.
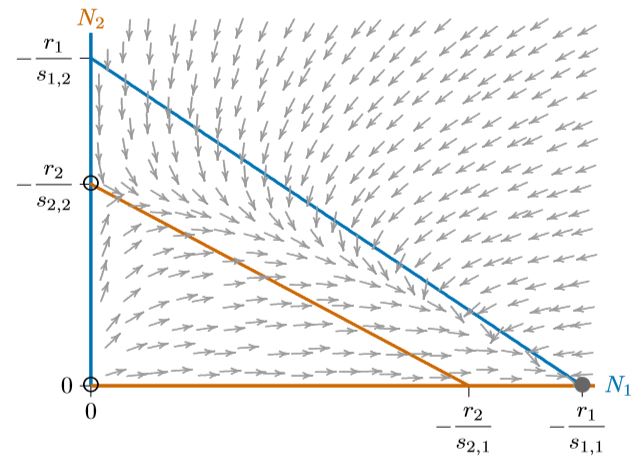
Dependiendo de donde comiencen las poblaciones, una de las dos especies excluirá a la otra. Una curva llamada “separatriz”, que divide estos puntos de partida según qué especies excluirán a la otra, se muestra con la línea discontinua negra en la Figura\(\PageIndex{4}\). Esa separatriz corresponde a una cresta larga y curvada en cualquier superficie por encima del espacio de fase, como se describe en el Capítulo 10. Pasa necesariamente por el equilibrio interior inestable. Aquí se trata de una curva simple, aunque en algunos casos (como el sistema Mandelbrot, que no representa competencia), las curvas relacionadas pueden ser infinitamente complejas.
 Copia y pega la imagen aquí
Copia y pega la imagen aquí
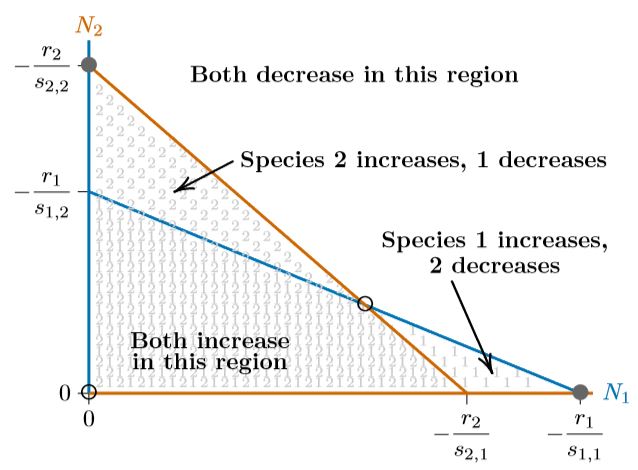
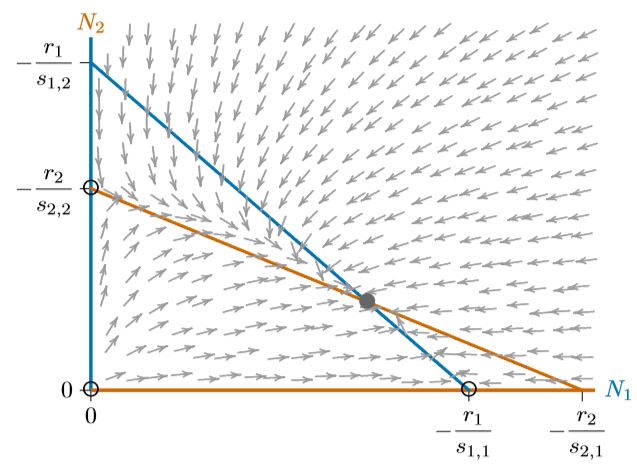
Finalmente, las isoclinas pueden cruzarse de manera opuesta, como en la Figura\(\PageIndex{5}\). En este caso, ninguna de las especies tiene ninguna región donde su isoclina encierra a la otra, como se ve en cada uno de los espacios de fase de las Figuras 16.5.4 a través\(\PageIndex{4}\). ¿Qué pasará cuando ninguna de las especies pueda excluir a la otra en alguna parte del espacio de fases? Se ven obligados a coexistir. Los equilibrios individuales en los ejes se vuelven inestables y el equilibrio interior se vuelve estable, de hecho, globalmente estable.
En resumen, las isoclinas en sistemas competitivos tienen cuatro configuraciones diferentes, como en la Figura\(\PageIndex{6}\). Los casos 1 y 2 pueden representar competencia por un solo recurso o, equivalentemente, competencia por dos recursos diferentes que las dos especies manejan de manera idéntica. Uno de los dos es un competidor superior que excluye al otro. El caso 3 es “biestable”, donde cualquiera de las especies puede excluir a la otra, dependiendo de cómo comience el sistema. Finalmente, el Caso 4 es globalmente estable, donde ninguna de las especies puede excluir a la otra, y prevalece la coexistencia estable. Los casos 3 y 4 pueden representar competencia por dos recursos diferentes.