
1.6: La ley del surtido independiente
- Page ID
- 57554
Introducción
La ley de la segregación nos permite predecir cómo se hereda una sola característica asociada a un solo gen. En algunos casos, sin embargo, podríamos querer predecir la herencia de dos características asociadas a dos genes diferentes. ¿Cómo podemos hacer esto?
- [Actualización sobre la ley de segregación]
-
La ley de segregación establece que cada gameto (espermatozoide u óvulo) hecho por un organismo obtendrá solo una de las dos copias génicas presentes en un organismo progenitor, y que las copias génicas se asignan aleatoriamente a los gametos. Por ejemplo, si un organismo tiene un genotipo de Aa, la mitad de sus gametos contendrá un alelo A, y la otra mitad contendrá un alelo a.
Puedes usar el enlace al inicio del párrafo para conocer más sobre la ley de segregación.
Para hacer una predicción precisa, necesitamos saber si los dos genes se heredan de forma independiente o no. Es decir, necesitamos saber si se “ignoran” entre sí cuando se clasifican en gametos, o si “se mantienen unidos” y se heredan como una unidad.
Cuando Gregor Mendel hizo esta pregunta, encontró que diferentes genes se heredaban independientemente unos de otros, siguiendo lo que se llama la ley del surtido independiente. En este artículo, analizaremos más de cerca la ley del surtido independiente y cómo se utiliza para hacer predicciones. También veremos cuándo y por qué la ley del surtido independiente lo hace (¡o no!) mantener la verdad.
Nota: Si aún no estás familiarizado con cómo se heredan los genes individuales, es posible que quieras consultar el artículo sobre la ley de segregación o el video de introducción a la herencia antes de sumergirte en este artículo.
¿Cuál es la ley del surtido independiente?
La ley de surtido independiente de Mendel establece que los alelos de dos (o más) genes diferentes se clasifican en gametos independientemente uno del otro. Es decir, el alelo que recibe un gameto por un gen no influye en el alelo recibido por otro gen.
Ejemplo: Color de guisante y genes de forma de guisante
Veamos un ejemplo concreto de la ley del surtido independiente. Imagina que cruzamos dos plantas de guisante de raza pura: una con semillas amarillas, redondas (YYRR) y otra con semillas verdes arrugadas (yyrr). Debido a que cada padre es homocigótico, la ley de la segregación nos dice que los gametos hechos por la planta arrugada y verde son todos ry, y los gametos hechos por la planta redonda y amarilla son todos RY. Eso nos da F 1 descendencia que son todas rRy.
El alelo que especifica el color amarillo de la semilla es dominante al alelo que especifica el color verde de la semilla, y el alelo que especifica la forma redonda es dominante al alelo que especifica la forma arrugada, como lo muestran las letras mayúsculas y minúsculas. Esto significa que las plantas F 1 son todas amarillas y redondas. Debido a que son heterocigóticas para dos genes, las plantas F 1 se denominan dihíbridos (di- = dos, -híbrido = heterocigótico).
Un cruce entre dos dihíbridos (o, equivalentemente, autofertilización de un dihíbrido) se conoce como cruce dihíbrido. Cuando Mendel hizo este cruce y miró a la descendencia, encontró que había cuatro categorías diferentes de semillas de guisante: amarillas y redondas, amarillas y arrugadas, verdes y redondas, y verdes y arrugadas. Estas categorías fenotípicas (categorías definidas por rasgos observables) aparecieron en una proporción de aproximadamente 9:3:3:1.
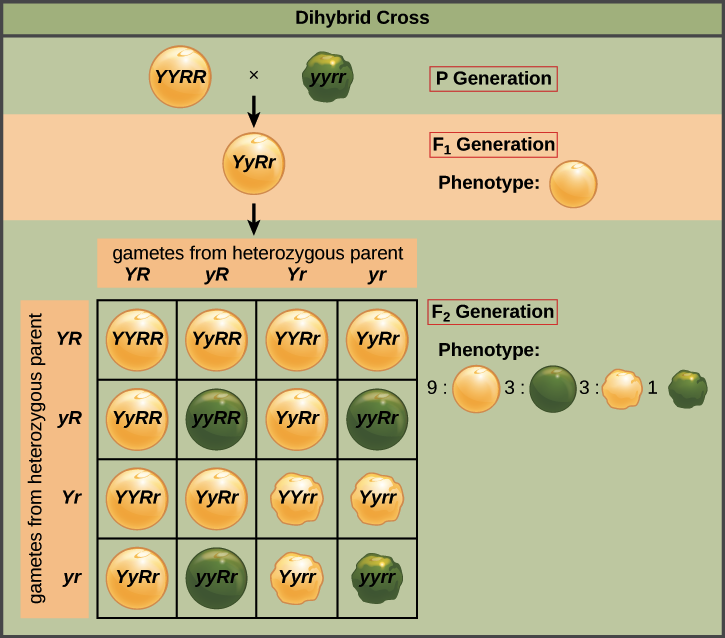
Esta relación fue la clave que llevó a Mendel a la ley del surtido independiente. Eso es porque una proporción 9:3:3:1 es exactamente lo que esperaríamos ver si la planta F 1 fabricara cuatro tipos de gametos (espermatozoides y óvulos) con igual frecuencia: YR, Yr, Yr y yr. En otras palabras, este es el resultado que predeciríamos si cada gameto obtuvo aleatoriamente un alelo Y o y, en un proceso separado, también obtuvo aleatoriamente un alelo R o r (haciendo cuatro combinaciones igualmente probables).
Podemos confirmar el vínculo entre los cuatro tipos de gametos y la relación 9:3:3:1 utilizando el cuadrado Punnett anterior. Para hacer el cuadrado, primero colocamos los cuatro tipos de gametos igualmente probables a lo largo de cada eje. Después, unimos gametos en los ejes en las casillas de la tabla, representando eventos de fertilización. Los 16 eventos de fertilización de igual probabilidad que pueden ocurrir entre los gametos se muestran en las 16 casillas. Los genotipos de descendencia en las cajas corresponden a una relación 9:3:3:1 de fenotipos, tal como observó Mendel.
- [Más sobre cuadrados Punnett de dos genes]
-
Podemos dibujar un cuadrado Punnett para un escenario de dos genes siguiendo las mismas reglas básicas que para un cruce monohíbrido, colocando los gametos a lo largo de los ejes y combinándolos en los cuadrados para representar eventos de fertilización. No obstante, dado que ahora hay más tipos de gametos, también debe haber más cuadrados en la tabla: 4 posibles tipos de gametos maternos x 4 posibles tipos de gametos paternos = 16 cuadrados totales.
Al igual que con un cuadrado Punnett de un solo gen, colocamos todos los tipos posibles de gametos a lo largo de los ejes, luego los combinamos en los cuadrados donde se cruzan las columnas y filas para representar eventos de fertilización (la formación de cigotos, o descendencia).
Para aprender cómo se pueden utilizar las reglas de probabilidad para predecir el resultado de un cruce dihíbrido, consulte el artículo sobre probabilidades en genética.
Surtido independiente vs. varillaje
La sección anterior nos da en pocas palabras la ley del surtido independiente de Mendel, y nos permite ver cómo la ley del surtido independiente conduce a una proporción 9:3:3:1. Pero, ¿cuál era la posibilidad alternativa? Es decir, ¿qué pasaría si dos genes no siguieran surtido independiente?
En el caso extremo, los genes para el color de la semilla y la forma de la semilla podrían haberse heredado siempre como pareja. Es decir, los alelos amarillos y redondos podrían haber permanecido siempre juntos, y también los alelos verdes y arrugados.
Para ver cómo esto podría funcionar, imagine que los genes de color y forma están físicamente pegados entre sí y no se pueden separar, como lo representan las cajas alrededor de los alelos en el diagrama a continuación. Por ejemplo, esto podría suceder si los dos genes se ubicaran muy, muy cerca entre sí en un cromosoma (idea que exploraremos más a fondo al final del artículo).
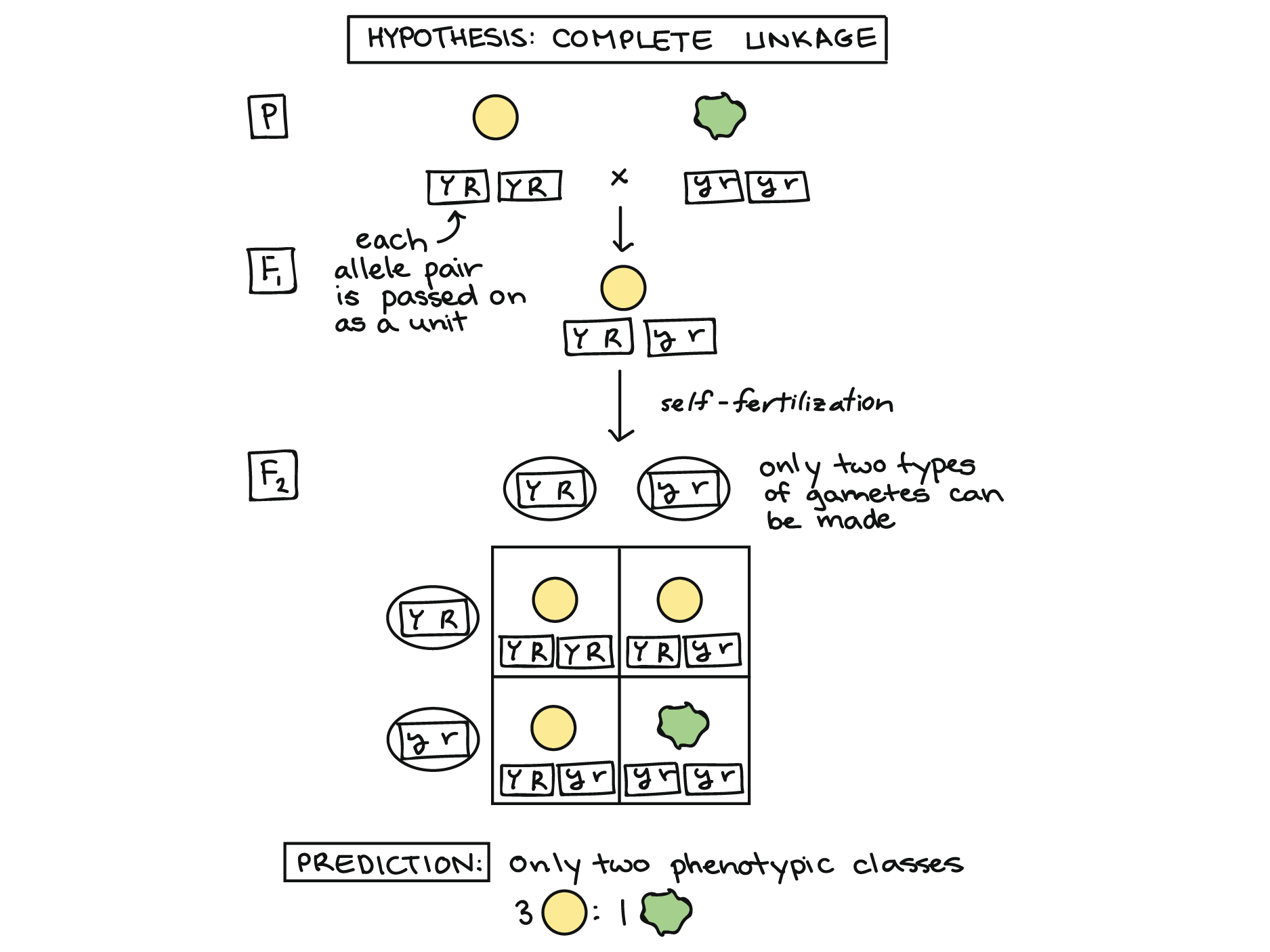
En lugar de dar un alelo de color y, por separado, darle un alelo de forma a cada gameto, la planta dihíbrida F 1 simplemente daría una “unidad combo” a cada gameto: un par de alelos YR o un par de alelos yr.
Podemos usar un cuadrado Punnett para predecir los resultados de la autofecundación en este caso, como se muestra arriba. Si los genes de color y forma de semilla de hecho siempre fueron heredados como una unidad, o completamente unidos, un cruce dihíbrido debería producir solo dos tipos de descendencia, amarilla/redonda y verde/arrugada, en una proporción 3:1. Los resultados reales de Mendel fueron bastante diferentes de esto (la relación 9:3:3:1 que vimos anteriormente), diciéndole que los genes surgieron de forma independiente.
La razón del surtido independiente
Para ver por qué ocurre el surtido independiente, necesitamos avanzar rápidamente medio siglo y descubrir que los genes se encuentran físicamente en los cromosomas. Para ser exactos, las dos copias de un gen portado por un organismo (como un alelo Y y) se localizan en el mismo punto en los dos cromosomas de un par homólogo. Los cromosomas homólogos son similares pero no idénticos, y un organismo obtiene un miembro del par de cada uno de sus dos progenitores.
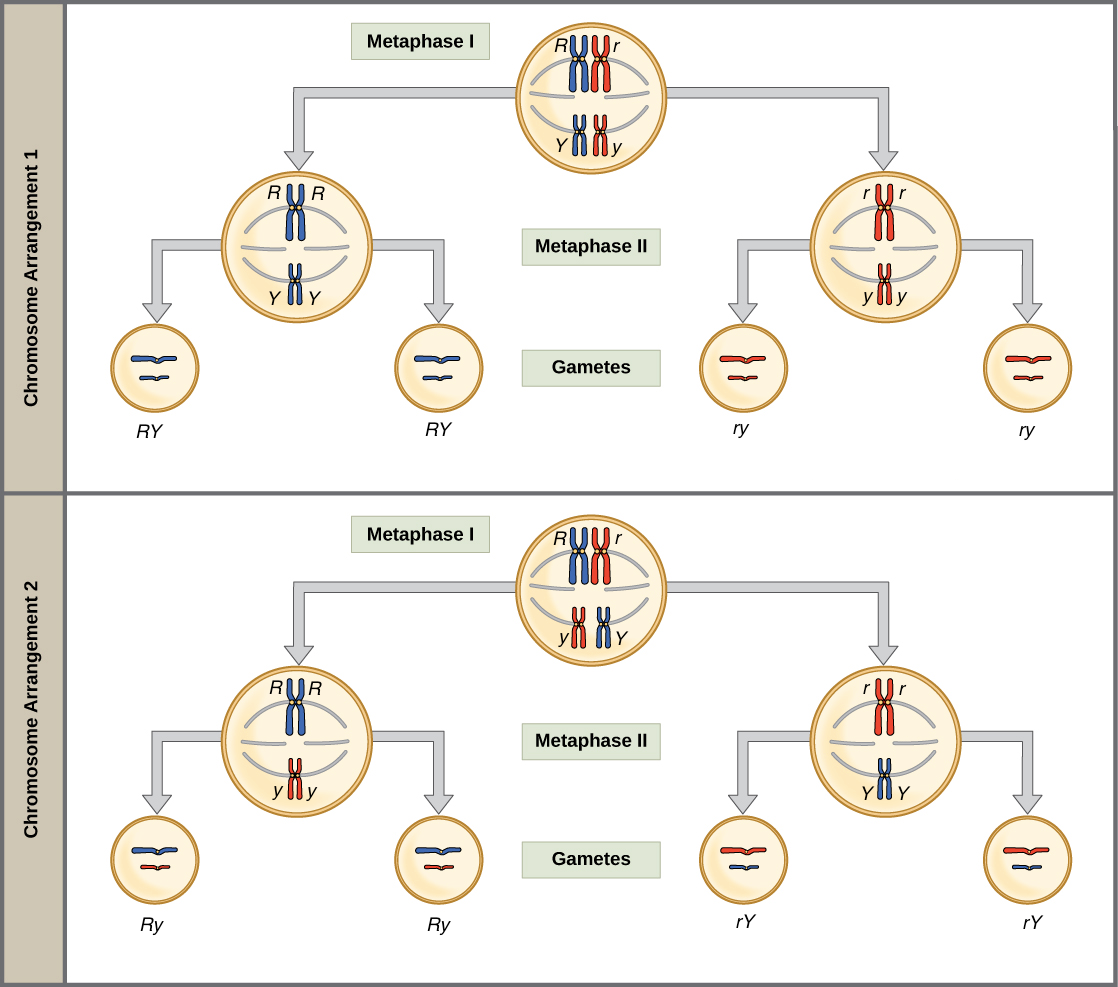
La base física de la ley del surtido independiente radica en la meiosis I de formación de gametos, cuando las parejas homólogas se alinean en orientaciones aleatorias en la mitad de la célula mientras se preparan para separarse. Podemos conseguir gametos con diferentes combos de homólogos de “mamá” y “papá” (y así, los alelos en esos homólogos) porque la orientación de cada par es aleatoria.
Para ver lo que esto significa, compare el arreglo cromosómico 1 (arriba) y el arreglo cromosómico 2 (abajo) en la etapa de metafase I en el diagrama a continuación. En un caso, los cromosomas rojos “mamá” van de la mano, mientras que en el otro, se separan y se mezclan con los cromosomas azules “papá”. Si la meiosis ocurre muchas veces, como ocurre en una planta de guisante, obtendremos ambos arreglos, y por lo tanto clases de gametos RY, Ry, Ry y ry, con igual frecuencia.
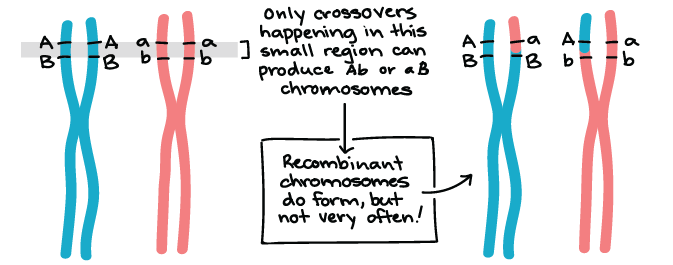
Los genes que se encuentran en diferentes cromosomas (como los genes Y y R) se asumen de forma independiente. Los genes de color y forma de semilla se encuentran en los cromosomas 1 y 7 del genoma del guisante, respectivamente, en la vida real 1. Los genes que están muy separados en el mismo cromosoma también se asumen independientemente gracias al cruce, o intercambio de bits de cromosomas homólogos, que ocurre temprano en la meiosis I.
- [Ver una foto]
-
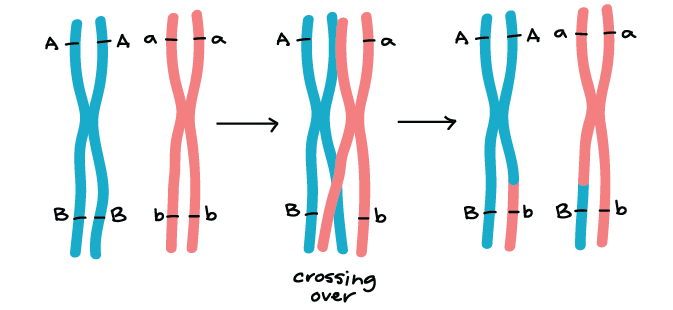
Existen, sin embargo, pares de genes que no se asumen de forma independiente. Cuando los genes están muy juntos en un cromosoma, los alelos del mismo cromosoma tienden a heredarse como una unidad con mayor frecuencia que no. Dichos genes no muestran surtido independiente y se dice que están vinculados. Analizaremos más de cerca la vinculación genética en otros artículos y videos.
- [Ver una foto]
-
Verifica tu comprensión
Consulta\(\PageIndex{1}\)
- [Pista]
-
Dado que todos los perros F 1 son negros y de pelo recto, sabemos que el color del pelaje negro y la textura del pelaje recto son dominantes sobre el color amarillo y la textura del pelaje rizado. Si llamamos al gen de color b/b y al gen de textura c/C, y usamos letras mayúsculas para la forma dominante de cada gen y letras minúsculas para la forma recesiva, podemos asignar los dos genotipos de perros parentales de bbCC (negro y rizado) y bbCC (amarillo y de pelaje recto). Cuando se cruzan los perros parentales, producen perros F 1 negros de pelo recto que son dihíbridos: BBcc.
Un cruce entre dos perros dihíbridos F 1 da como resultado el cuadrado Punnett que se muestra a continuación. Los perros F 1 pueden hacer cuatro tipos diferentes de gametos, los cuales se representan a lo largo de los dos ejes del cuadrado Punnett. Los cuadrados de la tabla representan eventos de fertilización en los que se combinan los gametos en los ejes. Dado que todos los tipos de gametos son igualmente probables de ser producidos (porque los genes se asumen de forma independiente, es decir, no influyen en la herencia de los demás), todos los cuadrados de la tabla representan eventos de igual probabilidad, los que ocurren 1/16 del tiempo.
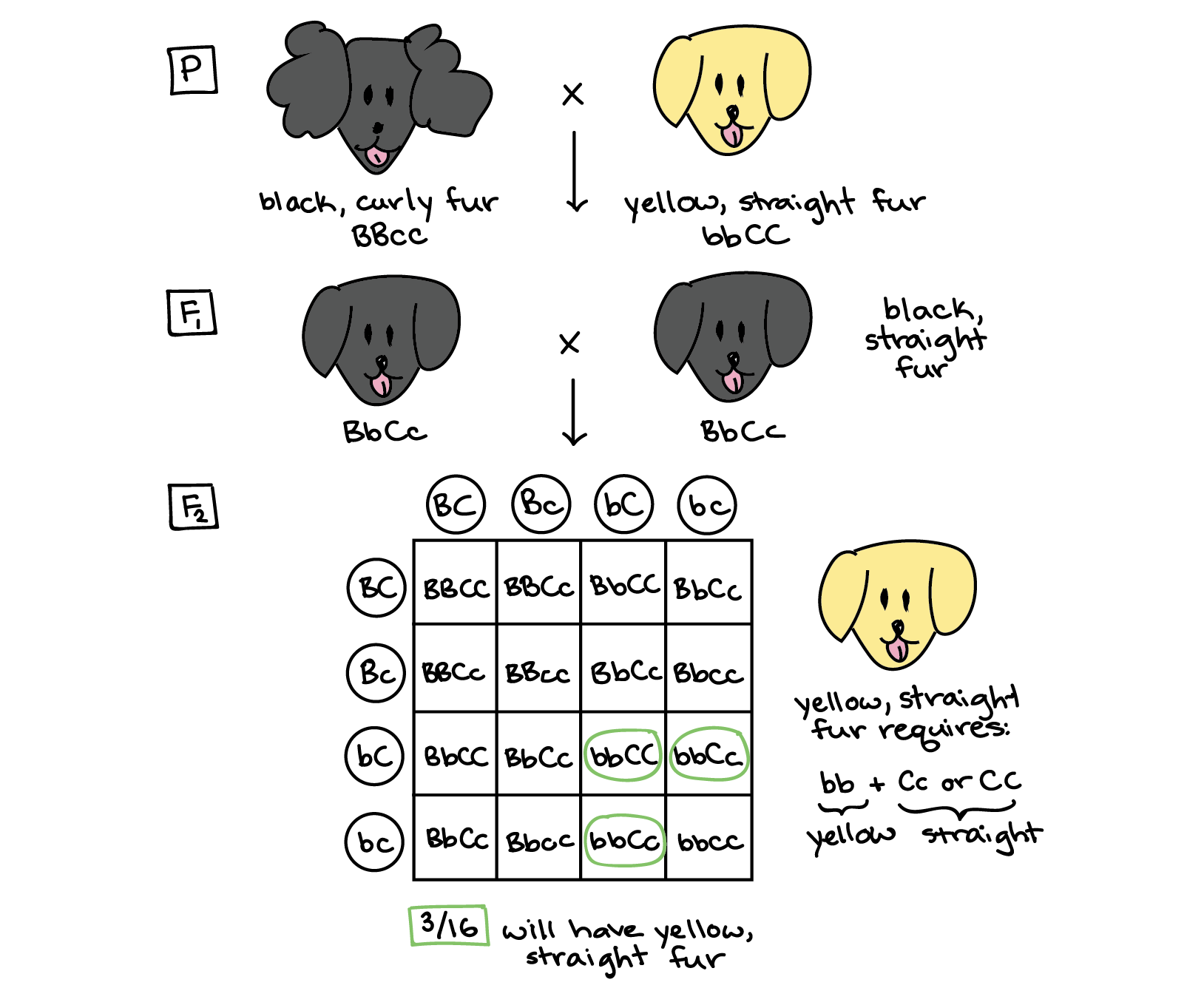
Ahora, necesitamos encontrar los cuadrados que corresponden al resultado que nos interesa: un cachorro de pelaje amarillo, recto. Para tener pelaje amarillo y recto, el cachorro debe obtener dos alelos recesivos para el color del pelaje (genotipo bb) y al menos un alelo dominante para la textura del pelaje (genotipo Cc o CC). Si pasamos por la tabla y rodeamos los genotipos que coinciden con estos requisitos, encontraremos que 3 de las 16 cajas corresponden a cachorros amarillos, de pelo recto. Así, esperaríamos que 3/16 de los cachorros F 2 tuvieran pelaje amarillo, recto.
Respuesta: 3/16.
Colaboradores y Atribuciones
- [Atribución y referencias]
-
Atribución:
Este artículo es un derivado modificado de:
- “Leyes de herencia”, por OpenStax College, Biología (CC BY 3.0). Descarga gratis el artículo original en http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@9.85.
- “Leyes de herencia”, en Principios de Biología, de Robert Bear, David Rintoul, Bruce Snyder, Martha Smith-Caldas, Christopher Herren, y Eva Horne, OpenStax, (CC BY 4.0). Descarga gratis el artículo original en http://cnx.org/contents/db89c8f8-a27c-4685-ad2a-19d11a2a7e2e@24.18.
El artículo modificado está licenciado bajo una licencia CC BY-NC-SA 4.0.
Obras citadas:
- Reid, J. B., y Ross, J. J. (2011). Los genes de Mendel: Hacia una caracterización molecular completa. Genética 189 (1), 3-10. http://dx.doi.org/10.1534/genetics.111.132118. Recuperado de www.ncbi.nlm.nih.gov/pmc/articles/PMC3176118/.
Referencias:
Dihíbrido. (2015). En El diccionario gratuito. Recuperado de http://www.thefreedictionary.com/dihybrid.
Kimball, J. W. (2014, 21 de abril). Vinculación genética y mapas genéticos. En las páginas de biología de Kimball. Recuperado a partir de https://www.biology-pages.info/L/Linkage.html
Purves, W. K., Sadava, D. E., Orians, G. H., y Heller, H.C. (2004). Genética: Mendel y más allá. En la vida: La ciencia de la biología (7ª ed., pp. 187-212). Sunderland, MA: Asociados Sinauer.
Raven, P. H., Johnson, G. B., Mason, K. A., Losos, J. B., y Singer, S. R. (2014). Patrones de herencia. En Biología (10a ed., AP ed., pp. 221-238). Nueva York, NY: McGraw-Hill.
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). Mendel y la idea genética. En Campbell Biology (10a ed., pp. 267-291). San Francisco, CA: Pearson.
Reid, J. B., y Ross, J. J. (2011). Los genes de Mendel: Hacia una caracterización molecular completa. Genética 189 (1), 3-10. http://dx.doi.org/10.1534/genetics.111.132118. Recuperado de www.ncbi.nlm.nih.gov/pmc/articles/PMC3176118/.
El Proyecto Adapa. (2014, 13 de agosto). ¿Cuáles son las leyes de segregación y surtido independiente y por qué son tan importantes? En BioBook. Recuperado de https://adapaproject.org/bbk_temp/tiki-index.php?page=Leaf%3A+What+are+the+laws+of+segregation+and+independent+assortment+and+why+are+they+so+important%3F.