
19.7B: Traducción
- Page ID
- 55495

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Definir traducción.
- Describa brevemente la función de lo siguiente en términos de traducción:
- Subunidad ribosómica 30S
- sitio de unión al ribosoma
- codón de inicio
- complejo de iniciación
- Subunidad ribosómica 50S
- ARNt
- Aminoacil-ARNt
- anticodón
- Sitio P del ribosoma
- Sitio A del ribosoma
- Sitio E del ribosoma
- peptidil-transferasa
- codón sin sentido (parada)
- factores de liberación

Una subunidad ribosómica 50S luego se une al complejo de iniciación y los factores de iniciación se van. Esto forma el ribosoma de los 70. (ver Figura\(\PageIndex{4}\)).
La unión de aminoácidos individuales para formar una proteína o polipéptido se conoce como la fase de elongación de la traducción. Hay tres sitios en el ribosoma de los años 70. El sitio A o aceptor o aminoacilo es donde se une primero un aminoacil-ARNt. El sitio P o péptido es donde un ARNt sostiene temporalmente la cadena de aminoácidos en crecimiento a medida que se lee el siguiente codón en el ARNm. El sitio E o salida es donde el ARNt no cargado que ha liberado su aminoácido sale del ribosoma. Durante la formación del enlace peptídico, la cadena de aminoácidos o péptido se mueve desde el ARNt en el sitio P y forma un enlace peptídico con el nuevo aminoácido unido al ARNt en el sitio A. El enlace peptídico está formado por una ribozima, una enzima compuesta por el propio ARNr 23S, llamada peptidil-transferasa. El ARNt ahora no cargado en el sitio P deja el ribosoma a través del sitio E para finalmente recoger un nuevo aminoácido y ser reciclado. En tanto, el ribosoma 70S mueve una distancia de un codón hacia abajo del ARNm a través de un proceso llamado translocación para permitir la decodificación del siguiente codón en el mensaje (ver Figura\(\PageIndex{5}\) A - 5F). La cadena polipeptídica en crecimiento pasa a través de un túnel en la subunidad ribosómica 50S.
Este proceso continúa una y otra vez en la dirección 5' a 3' hasta que el ribosoma alcanza un codón de parada. Un codón de parada es una serie de tres bases de ARNm que no codifican ningún aminoácido y así termina la cadena proteica. UAA, UAG, UGA son los tres codones de parada en el código genético. Los codones de parada no codifican un aminoácido porque un ARNt no puede reconocerlos.
Las proteínas llamadas factores de liberación liberan la proteína del ARNt y las dos subunidades ribosómicas se separan para ser recicladas (ver Figura\(\PageIndex{5}\) F). Durante este proceso de elongación, la proteína ha asumido su forma funcional tridimensional. Las proteínas llamadas chaperoninas ayudan en el plegamiento de las proteínas.
| Animación 3D ilustrando la traducción. De Drew Berry, wehi.edu.au. Esta animación tarda algún tiempo en cargarse. |
Una vez que el ribosoma está libre del sitio de unión al ribosoma y el codón de inicio AUG, otra subunidad ribosómica 30S se une al sitio de unión al ribosoma del ARNm para iniciar otra ronda de traducción. De esta manera, se pueden producir múltiples copias de una proteína a partir de una sola molécula de ARNm. Un ARNm con múltiples ribosomas unidos se conoce como polirribosoma o polisoma.
Resumen
- Durante la traducción, los ARNt específicos recogen aminoácidos específicos, los transfieren a los ribosomas y los insertan en su lugar apropiado de acuerdo con el “mensaje” genético del ARNm.
- Esto se hace mediante la porción anticodón de las moléculas de ARNt que se empareja de bases complementarias con los codones a lo largo del ARNm.
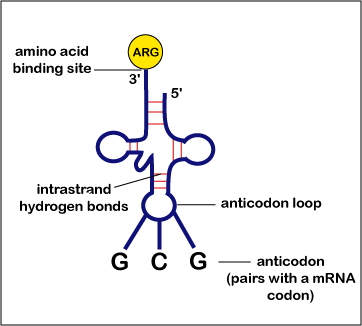
- El ARN de transferencia (ARNt) es una molécula de ARN tridimensional con forma de hoja de trébol invertida.
- En la parte superior, o extremo 3', se puede unir un aminoácido específico a un ARNt específico por medio de enzimas específicas llamadas aminoacil-ARNt sintetasas.
- En el bucle inferior de la hoja de trébol hay una serie de tres bases de ARNt desapareadas llamadas anticodón. Un anticodón es una serie de tres bases de ARNt complementarias a un codón de ARNm.
- Los anticodones de algunos ARNt son capaces de reconocer más de un codón debido a que el reconocimiento del ARNt del tercer nucleótido del codón no siempre es preciso, sin embargo, el aminoácido correcto aún se inserta porque hay 61 codones que codifican para los 22 aminoácidos diferentes.
- Para iniciar la traducción en células procariotas, una subunidad ribosómica 30S se une a una secuencia de nucleótidos corta en el ARNm llamada sitio de unión al ribosoma.
- AUG es conocido como el codón de inicio. En este punto, se forma un complejo de iniciación compuesto por la subunidad 30S, un ARNt que tiene el anticodón UAC y que porta una forma alterada del aminoácido metionina (N-formilmetionina o F-met), y proteínas llamadas factores de iniciación. Una subunidad ribosómica 50S luego se une al complejo de iniciación y los factores de iniciación se van. Esto forma el ribosoma de los 70.
- El sitio A o aceptor o aminoacilo del ribosoma es donde se une primero un aminoacil-ARNt.
- El sitio P o péptido del ribosoma es donde un ARNt sostiene temporalmente la cadena de aminoácidos en crecimiento a medida que se lee el siguiente codón en el ARNm.
- El E o sitio de salida del ribosoma es donde el ARNt no cargado que ha liberado su aminoácido sale del ribosoma.
- Durante la formación del enlace peptídico, la cadena de aminoácidos o péptido se mueve desde el ARNt en el sitio P y forma un enlace peptídico con el nuevo aminoácido unido al ARNt en el sitio A.
- Un codón de parada es una serie de tres bases de ARNm que no codifican ningún aminoácido y así termina la cadena proteica. UAA, UAG, UGA son los tres codones de parada en el código genético. (Los codones de parada no codifican un aminoácido porque un ARNt no puede reconocerlos).
- Un ARNm con múltiples ribosomas unidos se conoce como polirribosoma o polisoma.