
11.7: Regulación Génica - Teoría de Operones
- Page ID
- 54583

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Comparar operones inducibles y operones reprimibles
- Describir por qué es importante la regulación de operones
Cada célula nucleada en un organismo multicelular contiene copias del mismo ADN. De igual manera, todas las células en dos cultivos bacterianos puros inoculados de la misma colonia inicial contienen el mismo ADN, con la excepción de los cambios que surgen de mutaciones espontáneas. Si cada célula en un organismo multicelular tiene el mismo ADN, entonces ¿cómo es que las células en diferentes partes del cuerpo del organismo exhiben características diferentes? Del mismo modo, ¿cómo es que las mismas células bacterianas dentro de dos cultivos puros expuestos a diferentes condiciones ambientales pueden exhibir diferentes fenotipos? En ambos casos, cada célula genéticamente idéntica no enciende, ni expresa, el mismo conjunto de genes. Solo se expresa un subconjunto de proteínas en una célula en un momento dado.
El ADN genómico contiene tanto genes estructurales, que codifican productos que sirven como estructuras celulares o enzimas, como genes reguladores, que codifican productos que regulan la expresión génica. La expresión de un gen es un proceso altamente regulado. Mientras que la regulación de la expresión génica en organismos multicelulares permite la diferenciación celular, en organismos unicelulares como los procariotas, principalmente asegura que los recursos de una célula no se desperdicien produciendo proteínas que la célula no necesita en ese momento.
El esclarecimiento de los mecanismos que controlan la expresión génica es importante para la comprensión de la salud humana. Las disfunciones en este proceso en humanos conducen al desarrollo de cáncer y otras enfermedades. Comprender la interacción entre la expresión génica de un patógeno y la de su hospedador humano es importante para la comprensión de una enfermedad infecciosa particular. La regulación génica implica una compleja red de interacciones dentro de una célula dada entre señales del entorno celular, moléculas de señalización dentro de la célula y el ADN de la célula. Estas interacciones conducen a la expresión de algunos genes y a la supresión de otros, dependiendo de las circunstancias.
Procariotas y eucariotas comparten algunas similitudes en sus mecanismos para regular la expresión génica; sin embargo, la expresión génica en eucariotas es más complicada debido a la separación temporal y espacial entre los procesos de transcripción y traducción. Así, aunque la mayor parte de la regulación de la expresión génica ocurre a través del control transcripcional en procariotas, la regulación de la expresión génica en eucariotas ocurre a nivel transcripcional y postranscripcional (después de que se haya realizado el transcrito primario).
Regulación de genes procariotas
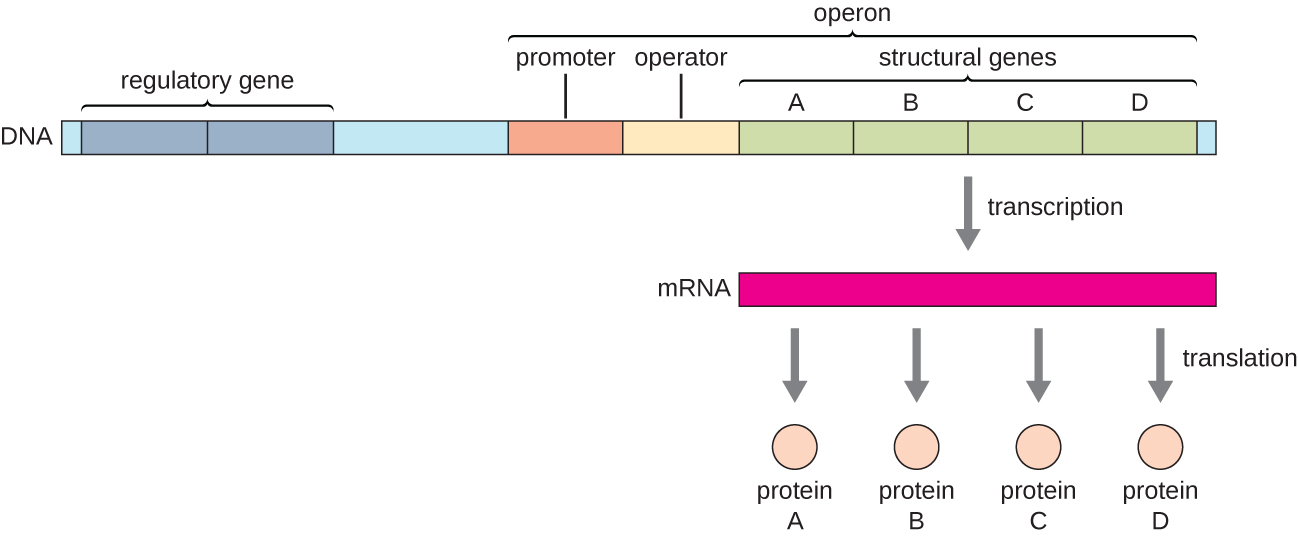
En bacterias y arqueas, las proteínas estructurales con funciones relacionadas generalmente se codifican juntas dentro del genoma en un bloque llamado operón y se transcriben juntas bajo el control de un solo promotor, dando como resultado la formación de un transcrito policistrónico (Figura\(\PageIndex{1}\)). De esta manera, la regulación de la transcripción de todos los genes estructurales que codifican las enzimas que catalizan los muchos pasos en una sola vía bioquímica puede controlarse simultáneamente, ya que o bien todos serán necesarios al mismo tiempo, o ninguno será necesario. Por ejemplo, en E. coli, todos los genes estructurales que codifican enzimas necesarias para usar lactosa como fuente de energía se encuentran uno al lado del otro en el operón de lactosa (o lac) bajo el control de un solo promotor, el promotor lac. Los científicos franceses François Jacob (1920—2013) y Jacques Monod del Instituto Pasteur fueron los primeros en mostrar la organización de genes bacterianos en operones, a través de sus estudios sobre el operón lac de E. coli. Por esta obra, ganaron el Premio Nobel de Fisiología o Medicina en 1965. Aunque los genes eucariotas no están organizados en operones, los operones procariotas son excelentes modelos para aprender sobre la regulación génica en general. Hay algunos grupos de genes en eucariotas que funcionan de manera similar a los operones. Muchos de los principios se pueden aplicar a los sistemas eucariotas y contribuir a nuestra comprensión de los cambios en la expresión génica en eucariotas que pueden resultar en cambios patológicos como el cáncer.
Cada operón incluye secuencias de ADN que influyen en su propia transcripción; estas se localizan en una región llamada región reguladora. La región reguladora incluye el promotor y la región que rodea al promotor, a la que pueden unirse factores de transcripción, proteínas codificadas por genes reguladores. Los factores de transcripción influyen en la unión de la ARN polimerasa al promotor y permiten su progresión para transcribir genes estructurales. Un represor es un factor de transcripción que suprime la transcripción de un gen en respuesta a un estímulo externo al unirse a una secuencia de ADN dentro de la región reguladora llamada operador, que se encuentra entre el sitio de unión a la ARN polimerasa del promotor y el sitio de inicio de la transcripción del primer gen estructural. La unión del represor bloquea físicamente la ARN polimerasa para que no transcriba genes estructurales. Por el contrario, un activador es un factor de transcripción que aumenta la transcripción de un gen en respuesta a un estímulo externo al facilitar la unión de la ARN polimerasa al promotor. Un inductor, un tercer tipo de molécula reguladora, es una molécula pequeña que activa o reprime la transcripción al interactuar con un represor o un activador.
En procariotas, hay ejemplos de operones cuyos productos génicos se requieren de manera bastante consistente y cuya expresión, por lo tanto, no está regulada. Dichos operones se expresan constitutivamente, es decir, se transcriben y traducen continuamente para proporcionar a la célula niveles intermedios constantes de los productos proteicos. Dichos genes codifican enzimas involucradas en las funciones de mantenimiento necesarias para el mantenimiento celular, incluida la replicación, reparación y expresión del ADN, así como enzimas involucradas en el metabolismo central. En contraste, hay otros operones procariotas que se expresan solo cuando son necesarios y están regulados por represores, activadores e inductores.
Ejercicio\(\PageIndex{1}\)
- ¿Cuáles son las partes en la secuencia de ADN de un operón?
- ¿Qué tipos de moléculas reguladoras hay?
Regulación por Represión
Los operones procariotas son comúnmente controlados por la unión de represores a regiones operadoras, impidiendo así la transcripción de los genes estructurales. Dichos operones se clasifican como operones reprimibles o como operones inducibles. Los operones reprimibles, como el operón triptófano (trp), contienen típicamente genes que codifican enzimas necesarias para una ruta biosintética. Mientras el producto de la vía, como el triptófano, siga siendo requerido por la célula, se seguirá expresando un operón reprimible. Sin embargo, cuando el producto de la vía biosintética comienza a acumularse en la célula, eliminando la necesidad de que la célula continúe haciendo más, se reprime la expresión del operón. Por el contrario, los operones inducibles, como el operón lac de E. coli, a menudo contienen genes que codifican enzimas en una vía involucrada en el metabolismo de un sustrato específico como la lactosa. Estas enzimas solo se requieren cuando ese sustrato está disponible, por lo que la expresión de los operones se induce típicamente solo en presencia del sustrato.
El operón trp: un operón reprimible
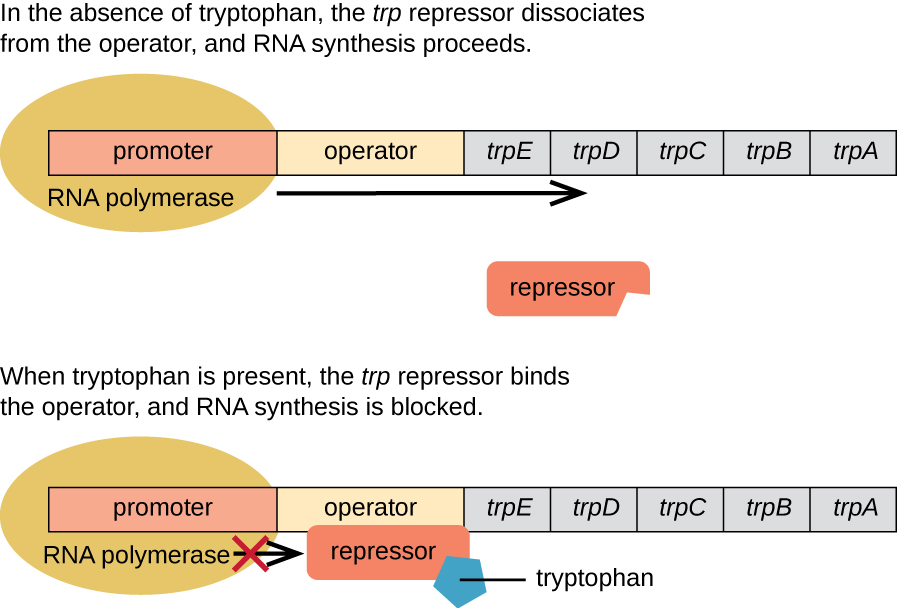
E. coli puede sintetizar triptófano usando enzimas codificadas por cinco genes estructurales ubicados uno al lado del otro en el operón trp (Figura\(\PageIndex{2}\)). Cuando el triptófano ambiental es bajo, el operón se enciende. Esto significa que se inicia la transcripción, se expresan los genes y se sintetiza triptófano. Sin embargo, si el triptófano está presente en el ambiente, el operón trp se apaga. La transcripción no ocurre y el triptófano no se sintetiza.
Cuando el triptófano no está presente en la célula, el represor por sí mismo no se une al operador; por lo tanto, el operón está activo y se sintetiza triptófano. Sin embargo, cuando el triptófano se acumula en la célula, dos moléculas de triptófano se unen a la molécula represora trp, lo que cambia de forma, permitiendo que se una al operador trp. Esta unión de la forma activa del represor trp al operador impide que la ARN polimerasa transcriba los genes estructurales, deteniendo la expresión del operón. Así, el producto real de la vía biosintética controlada por el operón regula la expresión del operón.
Mira este video para conocer más sobre el operón trp.
El operón lac: un operón inducible
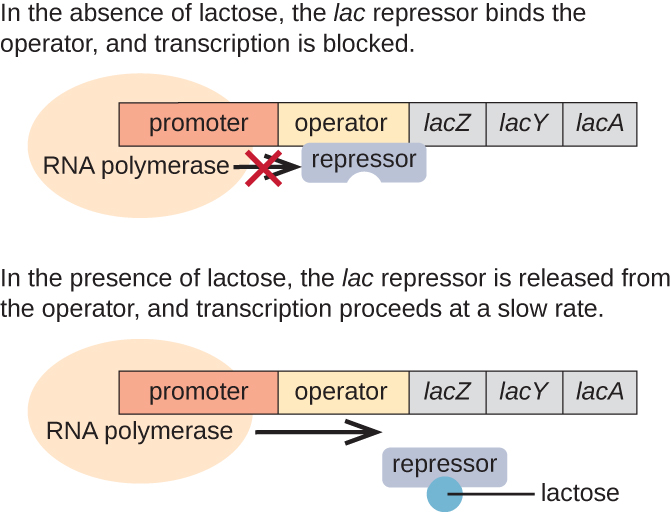
El operón lac es un ejemplo de un operón inducible que también está sujeto a activación en ausencia de glucosa (Figura\(\PageIndex{3}\)). El operón lac codifica tres genes estructurales necesarios para adquirir y procesar el disacárido lactosa del ambiente, descomponiéndolo en los azúcares simples glucosa y galactosa. Para que se exprese el operón lac, debe estar presente lactosa. Esto tiene sentido para la célula porque sería energéticamente derrochador crear las enzimas para procesar la lactosa si no estuviera disponible la lactosa.
En ausencia de lactosa, el represor lac se une a la región operadora del operón lac, impidiendo físicamente que la ARN polimerasa transcriba los genes estructurales. Sin embargo, cuando la lactosa está presente, la lactosa dentro de la célula se convierte en alolactosa. La alolactosa sirve como molécula inductora, uniéndose al represor y cambiando su forma para que ya no sea capaz de unirse al ADN del operador. La eliminación del represor en presencia de lactosa permite que la ARN polimerasa se mueva a través de la región operadora e inicie la transcripción de los genes estructurales lac.
El Operón lac: Activación por Proteína Activadora de Catabolitos
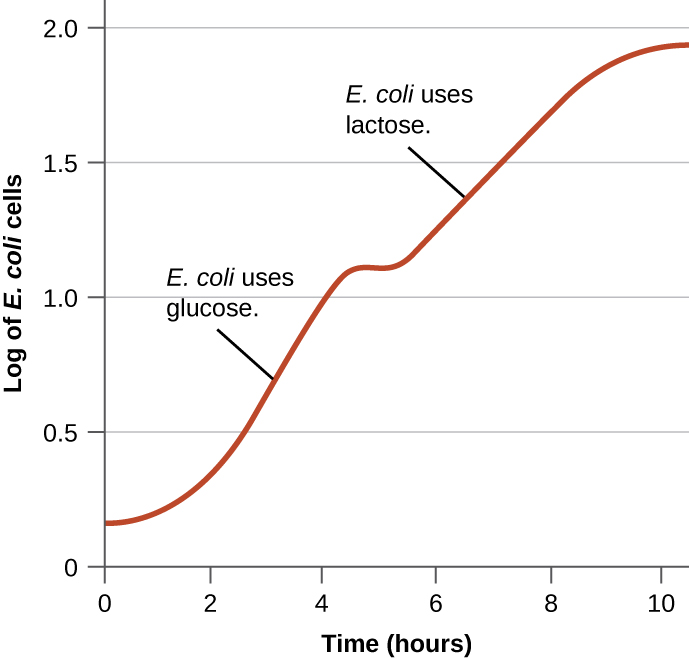
Las bacterias suelen tener la capacidad de usar una variedad de sustratos como fuentes de carbono. Sin embargo, debido a que la glucosa suele ser preferible a otros sustratos, las bacterias tienen mecanismos para asegurar que los sustratos alternativos solo se utilicen cuando la glucosa se haya agotado. Adicionalmente, las bacterias tienen mecanismos para asegurar que los genes que codifican enzimas para el uso de sustratos alternativos se expresen solo cuando el sustrato alternativo está disponible. En la década de 1940, Jacques Monod fue el primero en demostrar la preferencia por ciertos sustratos sobre otros a través de sus estudios del crecimiento de E. coli cuando se cultivó en presencia de dos sustratos diferentes simultáneamente. Dichos estudios generaron curvas de crecimiento diáuxico, como la que se muestra en la Figura\(\PageIndex{4}\). Aunque el sustrato preferido de glucosa se usa primero, E. coli crece rápidamente y las enzimas para el metabolismo de la lactosa están ausentes. Sin embargo, una vez agotados los niveles de glucosa, las tasas de crecimiento se ralentizan, induciendo la expresión de las enzimas necesarias para el metabolismo del segundo sustrato, la lactosa. Observe cómo la tasa de crecimiento en lactosa es más lenta, como lo indica la menor pendiente de la curva de crecimiento.
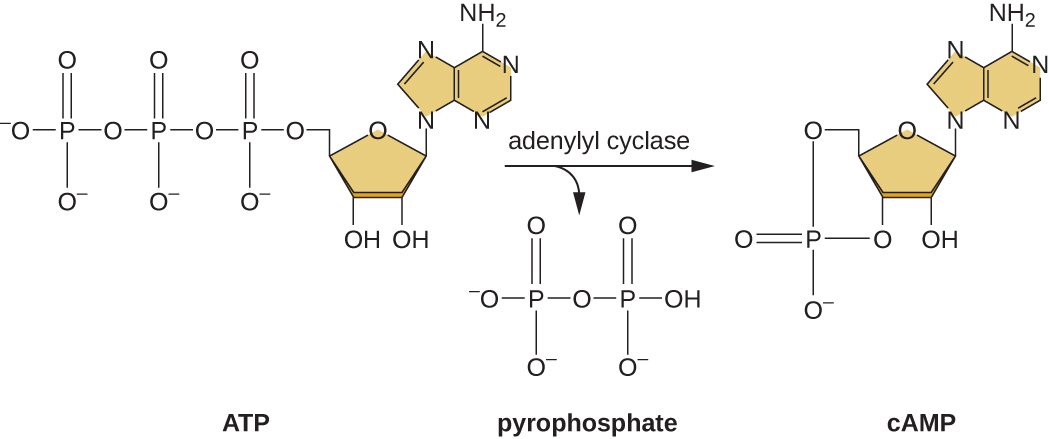
La capacidad de cambiar del uso de glucosa a otro sustrato como la lactosa es consecuencia de la actividad de una enzima llamada Enzima IIA (EIIA). Cuando los niveles de glucosa bajan, las células producen menos ATP a partir del catabolismo (ver Catabolismo de Carbohidratos), y EIIA se fosforila. La EIIA fosforilada activa la adenilil ciclasa, una enzima que convierte parte del ATP restante en AMP cíclico (AMPc), un derivado cíclico del AMP e importante molécula de señalización involucrada en el metabolismo de la glucosa y la energía en E. coli. Como resultado, los niveles de AMPc comienzan a subir en la celda (Figura\(\PageIndex{5}\)).
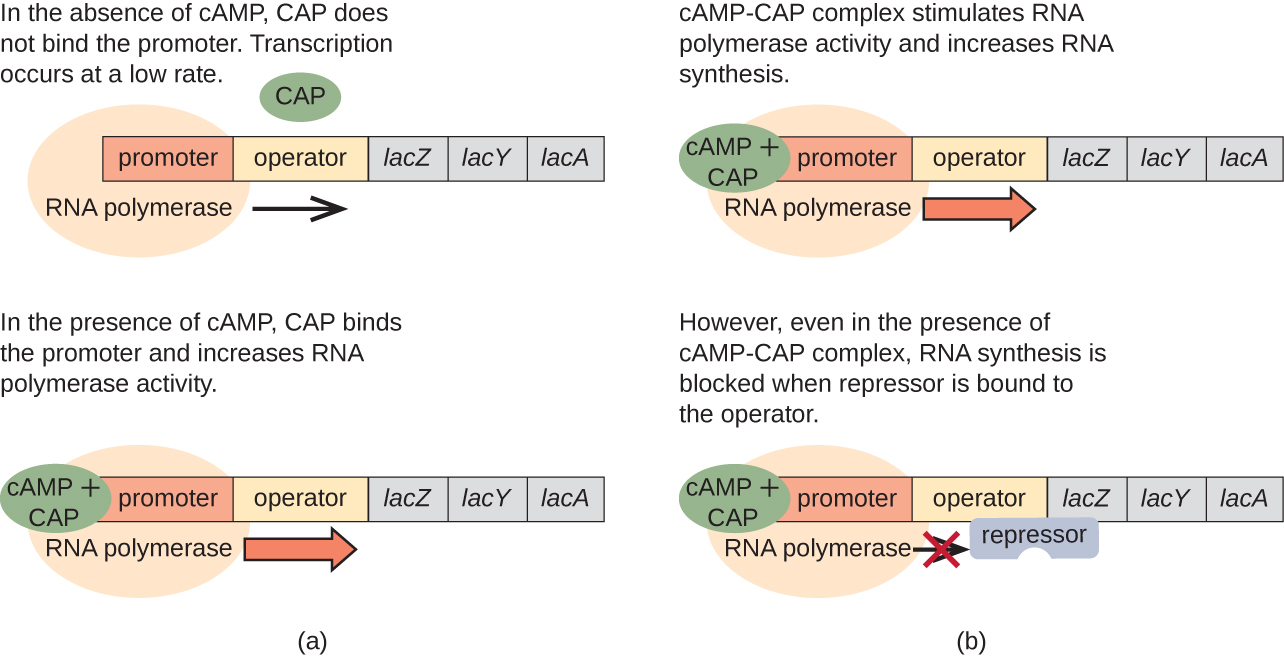
El operón lac también juega un papel en este cambio de usar glucosa a usar lactosa. Cuando la glucosa es escasa, el AMPc acumulado causado por el aumento de la actividad de la adenilil ciclasa se une a la proteína activadora de catabolitos (CAP), también conocida como proteína receptora de AMPc (CRP). El complejo se une a la región promotora del operón lac (Figura\(\PageIndex{6}\)). En las regiones reguladoras de estos operones, un sitio de unión a CAP se localiza aguas arriba del sitio de unión a la ARN polimerasa en el promotor. La unión del complejo CAP-cAMP a este sitio aumenta la capacidad de unión de la ARN polimerasa a la región promotora para iniciar la transcripción de los genes estructurales. Así, en el caso del operón lac, para que se produzca la transcripción, debe estar presente lactosa (eliminando la proteína represora lac) y los niveles de glucosa deben agotarse (permitiendo la unión de una proteína activadora). Cuando los niveles de glucosa son altos, hay represión catabolita de operones que codifican enzimas para el metabolismo de sustratos alternativos. Debido a los bajos niveles de AMPc bajo estas condiciones, existe una cantidad insuficiente del complejo CAP-cAMP para activar la transcripción de estos operones. Consulte\(\PageIndex{1}\) el Cuadro para un resumen de la regulación del operón lac.
| Glucosa | CAP se une | Lactosa | Represor se une | Transcripción |
|---|---|---|---|---|
| + | — | — | + | No |
| + | — | + | — | Algunos |
| — | + | — | + | No |
| — | + | + | — | Sí |
Mira un tutorial animado sobre el funcionamiento del operón lac aquí.
Ejercicio\(\PageIndex{2}\)
- ¿Qué afecta la unión del represor del operón trp al operador?
- ¿Cómo y cuándo se altera el comportamiento de la proteína represora lac?
- Además de ser reprimible, ¿de qué otra manera se regula el operón lac?
Respuestas Globales de Procariotas
En procariotas, también hay varios niveles más altos de regulación génica que tienen la capacidad de controlar la transcripción de muchos operones relacionados simultáneamente en respuesta a una señal ambiental. Un grupo de operones todos controlados simultáneamente se llama regulón.
Alarmones
Al detectar estrés inminente, los procariotas alteran la expresión de una amplia variedad de operones para responder en coordinación. Esto lo hacen a través de la producción de alarmonas, que son pequeños derivados de nucleótidos intracelulares. Las alarmonas cambian qué genes se expresan y estimulan la expresión de genes específicos de respuesta al estrés. El uso de alarmonas para alterar la expresión génica en respuesta al estrés parece ser importante en bacterias patógenas. Al encontrar mecanismos de defensa del huésped y otras condiciones duras durante la infección, muchos operones que codifican genes de virulencia están regulados positivamente en respuesta a la señalización de alarmonas. El conocimiento de estas respuestas es clave para poder comprender completamente el proceso de infección de muchos patógenos y para el desarrollo de terapias para contrarrestar este proceso.
Factores σ alternos
Dado que la subunidad σ de la ARN polimerasa bacteriana confiere especificidad en cuanto a qué promotores deben transcribirse, alterar el factor σ utilizado es otra forma para que las bacterias cambien rápida y globalmente qué regulones se transcriben en un momento dado. El factor σ reconoce secuencias dentro de un promotor bacteriano, por lo que diferentes factores σ reconocerán cada uno secuencias promotoras ligeramente diferentes. De esta manera, cuando la célula detecta condiciones ambientales específicas, puede responder cambiando qué factor σ expresa, degradando el antiguo y produciendo uno nuevo para transcribir los operones que codifican genes cuyos productos serán útiles bajo la nueva condición ambiental. Por ejemplo, en bacterias esporulantes de los géneros Bacillus y Clostridium (que incluyen muchos patógenos), un grupo de factores σ controla la expresión de los muchos genes necesarios para la esporulación en respuesta a señales estimulantes de esporulación.
Ejercicio\(\PageIndex{3}\)
- ¿Cuál es el nombre que se le da a una colección de operones que se pueden regular como grupo?
- ¿Qué tipo de estímulo desencadenaría la transcripción de un factor σ diferente?
Métodos Adicionales de Regulación en Bacterias: Atenuación y Ribointerruptores
Aunque la mayor parte de la expresión génica está regulada a nivel de iniciación de la transcripción en procariotas, también existen mecanismos para controlar tanto la finalización de la transcripción como la traducción simultáneamente. Desde su descubrimiento, se ha demostrado que estos mecanismos controlan la finalización de la transcripción y traducción de muchos operones procariotas. Debido a que estos mecanismos vinculan directamente la regulación de la transcripción y la traducción, son específicos de los procariotas, porque estos procesos están físicamente separados en eucariotas.
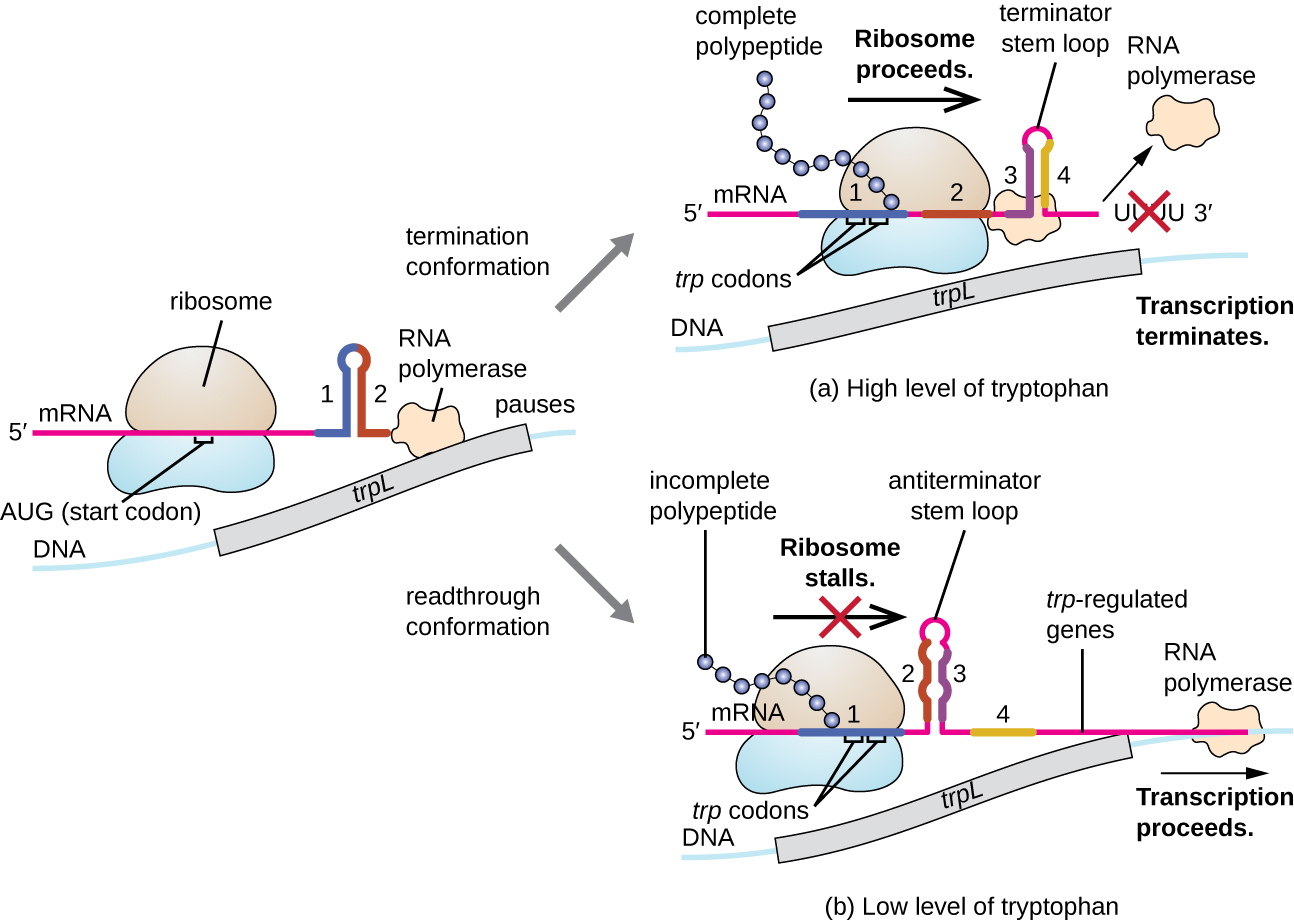
Uno de tales sistemas reguladores es la atenuación, por lo que las estructuras secundarias tallo-bucle formadas dentro del extremo 5' de un ARNm que se transcribe determinan si se producirá la transcripción para completar la síntesis de este ARNm y si este ARNm se utilizará para la traducción. Más allá del mecanismo de represión transcripcional ya discutido, la atenuación también controla la expresión del operón trp en E. coli (Figura\(\PageIndex{7}\)). La región reguladora del operón trp contiene una secuencia líder llamada trPL entre el operador y el primer gen estructural, que tiene cuatro tramos de ARN que pueden emparejarse entre sí en diferentes combinaciones. Cuando se forma un tallo-bucle terminador, la transcripción termina, liberando ARN polimerasa del ARNm. Sin embargo, cuando se forma un tallo-bucle antiterminador, esto evita la formación del tallo-bucle terminador, por lo que la ARN polimerasa puede transcribir los genes estructurales.
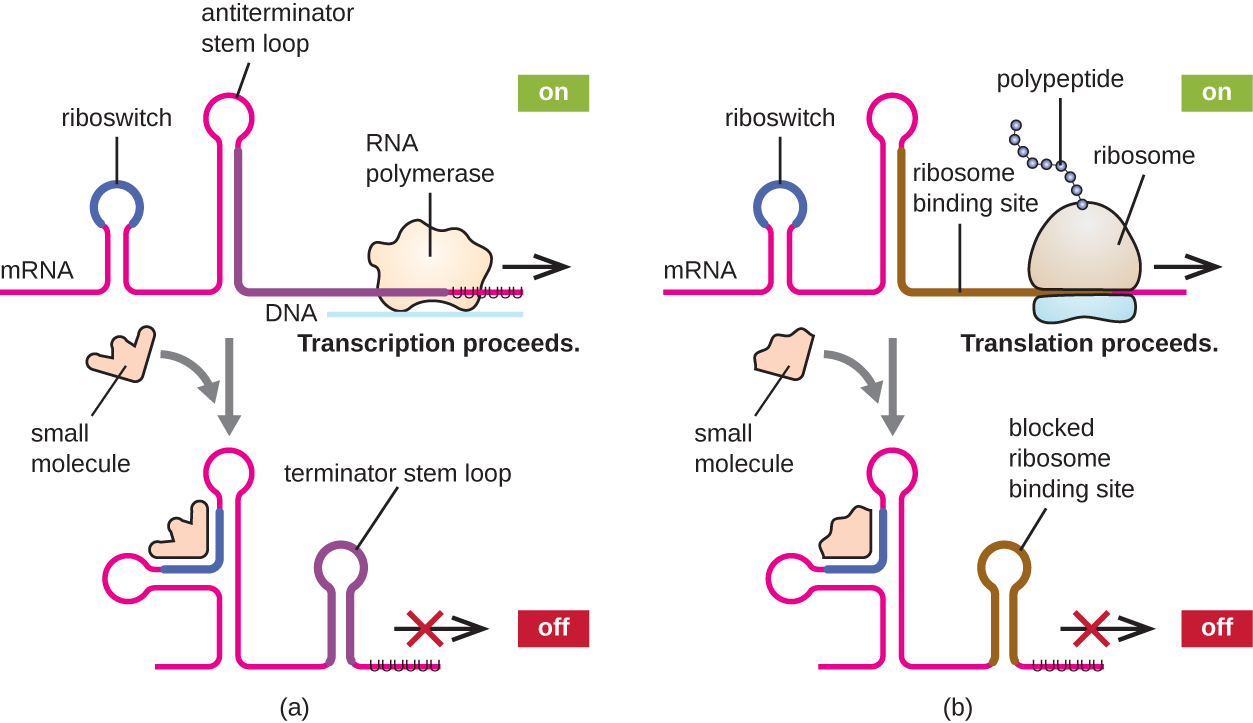
Un mecanismo relacionado de regulación concurrente de la transcripción y traducción en procariotas es el uso de un riboswitch, una pequeña región de ARN no codificante que se encuentra dentro del extremo 5' de algunas moléculas de ARNm procariotas (Figura\(\PageIndex{8}\)). Un riboswitch puede unirse a una pequeña molécula intracelular para estabilizar ciertas estructuras secundarias de la molécula de ARNm. La unión de la molécula pequeña determina qué estructura tallo-bucle se forma, influyendo así en la finalización de la síntesis de ARNm y la síntesis de proteínas.
Otros factores que afectan la expresión génica en procariotas y eucariotas
Aunque el enfoque en nuestra discusión sobre el control transcripcional utilizó operones procariotas como ejemplos, el control transcripcional eucariótico es similar en muchos aspectos. Al igual que en los procariotas, la transcripción eucariota se puede controlar a través de la unión de factores de transcripción incluyendo represores y activadores. Curiosamente, la transcripción eucariota puede verse influenciada por la unión de proteínas a regiones del ADN, llamadas potenciadores, bastante alejadas del gen, a través del bucle de ADN facilitado entre el potenciador y el promotor (Figura\(\PageIndex{9}\)). En general, regular la transcripción es una forma altamente efectiva de controlar la expresión génica tanto en procariotas como en eucariotas. Sin embargo, el control de la expresión génica en eucariotas en respuesta a tensiones ambientales y celulares se puede lograr de maneras adicionales sin la unión de factores de transcripción a regiones reguladoras.
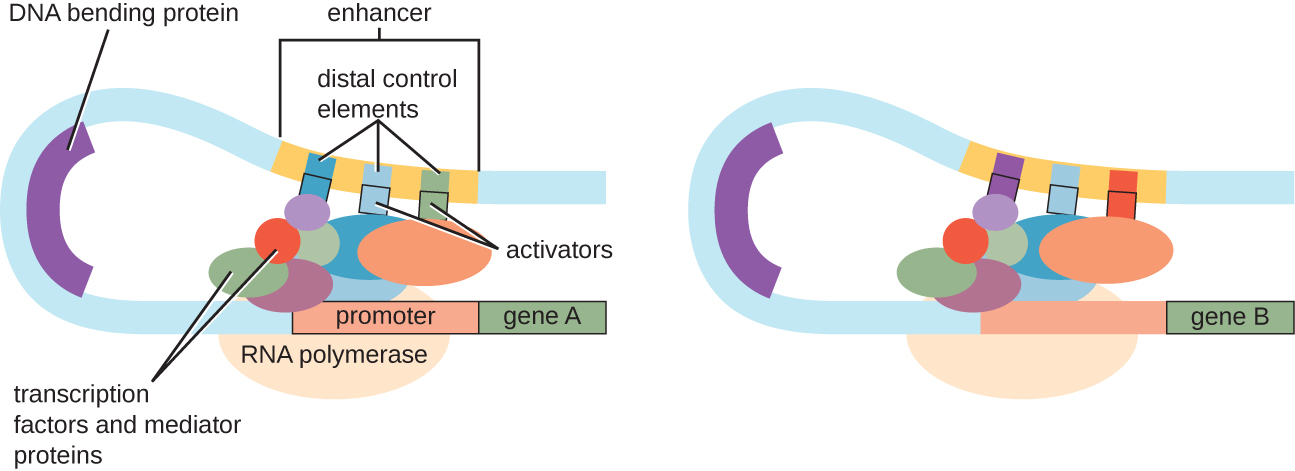
Control a nivel de ADN
En eucariotas, las moléculas de ADN o histonas asociadas pueden modificarse químicamente de tal manera que influyan en la transcripción; esto se llama regulación epigenética. Se ha demostrado que la metilación de ciertos nucleótidos de citosina en el ADN en respuesta a factores ambientales influye en el uso de dicho ADN para la transcripción, y la metilación del ADN se correlaciona comúnmente con niveles bajos de expresión génica. Adicionalmente, en respuesta a factores ambientales, las proteínas histonas para empaquetar ADN también pueden modificarse químicamente de múltiples maneras, incluyendo acetilación y desacetilación, influyendo en el estado de empaquetamiento del ADN y afectando así la disponibilidad de ADN de herida suelta para la transcripción. Estas modificaciones químicas a veces se pueden mantener a través de múltiples rondas de división celular, haciendo que al menos algunos de estos cambios epigenéticos sean heredables.
Este video describe cómo la regulación epigenética controla la expresión génica.
Ejercicio\(\PageIndex{}\)
- ¿Qué detiene o permite que la transcripción continúe cuando la atenuación está operando?
- ¿Qué determina el estado de un riboswitch?
- Describir la función de un potenciador.
- Describir dos mecanismos de regulación epigenética en eucariotas.
Foco Clínico: Resolución
Aunque Mark sobrevivió a su pelea con fascitis necrotizante, ahora tendría que someterse a una cirugía de injerto de piel, seguida de fisioterapia a largo plazo. Con base en la cantidad de masa muscular que perdió, es poco probable que su pierna vuelva a su plena fuerza, pero su fisioterapeuta es optimista de que recuperará algún uso de su pierna.
Las pruebas de laboratorio revelaron que el agente causante de la infección de Mark fue una cepa del estreptococo del grupo A (estreptococo del grupo A). Según lo exige la ley, el caso de Mark fue reportado al departamento de salud del estado y finalmente a los Centros para el Control y la Prevención de Enfermedades (CDC). En los CDC, la cepa del estreptococo del grupo A aislada de Mark se analizó más a fondo para determinar la resistencia a la meticilina.
La resistencia a la meticilina está codificada genéticamente y es cada vez más común en el estreptococo del grupo A a través de la transferencia genética horizontal. En la fascitis necrotizante, el flujo sanguíneo al área infectada suele ser limitado debido a la acción de diversas toxinas bacterianas genéticamente codificadas. Es por ello que normalmente hay poco o ningún sangrado como resultado de la prueba de incisión. Desafortunadamente, estas toxinas bacterianas limitan la efectividad de los antibióticos intravenosos para eliminar la infección de la piel y el tejido subyacente, lo que significa que la resistencia a los antibióticos por sí sola no explica la ineficacia del tratamiento de Mark. Sin embargo, la terapia antibiótica intravenosa fue necesaria para ayudar a minimizar el posible resultado de la sepsis, que es un resultado común de la fascitis necrotizante. A través del análisis genómico por los CDC de la cepa aislada de Mark, se demostró que varios de los genes de virulencia importantes estaban codificados en profagos, lo que indica que la transducción es importante en la transferencia horizontal de genes de estos genes de una célula bacteriana a otra.
Conceptos clave y resumen
- La expresión génica es un proceso estrechamente regulado.
- La expresión génica en procariotas está ampliamente regulada en el punto de transcripción. La expresión génica en eucariotas se regula adicionalmente postranscripcionalmente.
- Los genes estructurales procariotas de función relacionada a menudo se organizan en operones, todos controlados por la transcripción de un solo promotor. La región reguladora de un operón incluye el promotor mismo y la región que rodea al promotor a la que los factores de transcripción pueden unirse para influir en la transcripción.
- Aunque algunos operones se expresan constitutivamente, la mayoría están sujetos a regulación mediante el uso de factores de transcripción (represores y activadores). Un represor se une a un operador, una secuencia de ADN dentro de la región reguladora entre el sitio de unión de la ARN polimerasa en el promotor y el primer gen estructural, bloqueando así físicamente la transcripción de estos operones. Un activador se une dentro de la región reguladora de un operón, ayudando a que la ARN polimerasa se una al promotor, potenciando así la transcripción de este operón. Un inductor influye en la transcripción a través de la interacción con un represor o activador.
- El operón trp es un ejemplo clásico de operón reprimible. Cuando el triptófano se acumula, el triptófano se une a un represor, que luego se une al operador, impidiendo una mayor transcripción.
- El operón lac es un ejemplo clásico de un operón inducible. Cuando la lactosa está presente en la célula, se convierte en alolactosa. La alolactosa actúa como inductor, uniéndose al represor y evitando que el represor se una al operador. Esto permite la transcripción de los genes estructurales.
- El operón lac también está sujeto a activación. Cuando se agotan los niveles de glucosa, algo de ATP celular se convierte en AMPc, que se une a la proteína activadora de catabolitos (CAP). El complejo Camp-Cap activa la transcripción del operón lac. Cuando los niveles de glucosa son altos, su presencia impide la transcripción del operón lac y otros operones por represión catabolita.
- Pequeñas moléculas intracelulares llamadas alarmonas se elaboran en respuesta a diversas tensiones ambientales, permitiendo que las bacterias controlen la transcripción de un grupo de operones, llamado regulón.
- Las bacterias tienen la capacidad de cambiar qué factor σ de la ARN polimerasa utilizan en respuesta a las condiciones ambientales para cambiar rápida y globalmente qué regulones se transcriben.
- Los procariotas tienen mecanismos reguladores, incluyendo la atenuación y el uso de ribointerruptores, para controlar simultáneamente la finalización de la transcripción y traducción a partir de ese transcrito. Estos mecanismos funcionan a través de la formación de tallo-bucles en el extremo 5' de una molécula de ARNm que se está sintetizando actualmente.
- Existen puntos adicionales de regulación de la expresión génica en procariotas y eucariotas. En eucariotas, la regulación epigenética por modificación química de ADN o histonas, y la regulación del procesamiento del ARN son dos métodos.