
13.2A: Opsonización
- Page ID
- 55469

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Discutir cómo los anticuerpos defienden el cuerpo a través de la opsonización. (Incluya qué clases o isotipos de inmunoglobulinas están involucradas, el papel de la porción Fab del anticuerpo, el papel, si lo hay, de la porción Fc del anticuerpo, y el papel de cualquier proteína del complemento, si la hubiera, involucrada).
- Describa brevemente dos formas diferentes en que las bacterias pueden resistir la opsonización.
La opsonización, o unión potenciada, se refiere a las moléculas de anticuerpo IgG e IgE, las proteínas del complemento C3b y C4b, y otras opsoninas que unen antígenos a fagocitos. Esto da como resultado una fagocitosis mucho más eficiente.
Opsonización con IgG, IgA, C3b y C4b
El proceso comienza con anticuerpos del isotipo IgG, IgA o IgM que se hacen contra un antígeno de superficie del organismo o célula que se va a fagocitar. Las porciones Fab del anticuerpo reaccionan con epítopos del antígeno. La porción Fc de IgG (pero no IgM) puede entonces unirse a receptores en neutrófilos y macrófagos pegando así el antígeno al fagocito (Figura\(\PageIndex{1}\)). La porción Fc de IgA secretora también puede unirse a macrófagos y neutrófilos para opsonización.
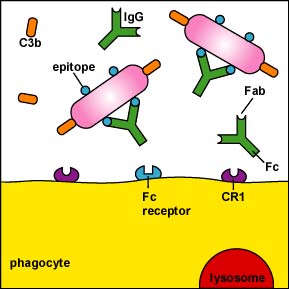
La porción Fc de IgA secretora también puede unirse a macrófagos y neutrófilos para opsonización.
Alternativamente, IgG, IgA e IgM pueden activar las vías del complemento (Figura\(\PageIndex{2}\)) y C3b o C4b pueden pegar el antígeno a los fagocitos (Figura\(\PageIndex{1}\)). Al igual que IgG, C3b, y en menor medida C4b, pueden funcionar como opsoninas, es decir, pueden unir antígenos a fagocitos.Una porción del C3b se une a proteínas y polisacáridos en superficies microbianas; otra porción se une a receptores CR1 en fagocitos, linfocitos B y células dendríticas para mejorar fagocitosis (Figura\(\PageIndex{3}\)). (Recuerde que C3b y C4b también se producen durante la vía alternativa del complemento y la vía lectina como se discutió en la Unidad 5.) La activación de la vía del complemento también promueve la inflamación para llevar los fagocitos y los químicos de defensa del torrente sanguíneo al sitio de la infección, como se discutirá más adelante en este tema.
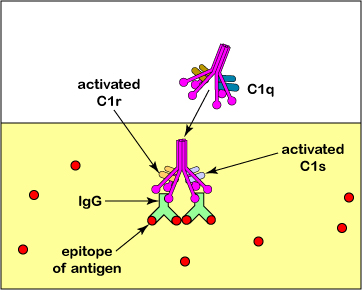
En realidad, la molécula C3b puede unirse a prácticamente cualquier proteína o polisacárido. Las células humanas, sin embargo, producen Factor H que se une a C3b y permite que el Factor I inactive el C3b. Por otro lado, sustancias como LPS en células bacterianas facilitan la unión del Factor B a C3b y esto protege al C3b de la inactivación por el Factor I. De esta manera, C3b no interactúa con nuestras propias células sino que es capaz de interactuar con células microbianas.
La unión promueve entonces la destrucción del antígeno. Los microorganismos se colocan en fagosomas (Figura\(\PageIndex{4}\)) donde finalmente son digeridos por los lisosomas (Figura\(\PageIndex{5}\)). Si el antígeno es una célula demasiado grande para ser ingerida -como las células hospedadoras infectadas por virus, las células trasplantadas y las células cancerosas-, el fagocito vacía el contenido de sus lisosomas directamente sobre la célula para la destrucción extracelular (Figura\(\PageIndex{6}\) y Figura\(\PageIndex{7}\)).
La opsonización es especialmente importante frente a microorganismos con estructuras antifagocíticas como las cápsulas ya que los anticuerpos opsonizantes hechos contra la cápsula son capaces de pegar cápsulas a los fagocitos (Figura\(\PageIndex{8}\)). En las vacunas contra la neumonía neumococal y Haemophilus influenzae tipo b, es el polisacárido capsular el que se administra como antígeno con el fin de estimular al organismo a elaborar anticuerpos opsonizantes contra la bacteria encapsulada.
Opsonización con IgE y promoción de la inflamación
El isotipo de anticuerpo IgE se elabora contra gusanos parásitos (helmintos) y artrópodos. Las porciones Fab de IgE se unen a epítopos en el helminto o artrópodo mientras que la porción Fc se une a receptores en eosinófilos permitiendo la opsonización. Es decir, la IgE pega eosinófilos fagocíticos a helmintos y artrópodos para la matanza extracelular de ese organismo (Figura\(\PageIndex{9}\)).

La porción Fc de IgE también se une a receptores en mastocitos y basófilos para desencadenar la liberación de mediadores inflamatorios (Figura\(\PageIndex{10}\)). La respuesta inflamatoria permite entonces que los fagocitos y los químicos de defensa salgan del torrente sanguíneo y vayan al sitio infectado como se discutirá más adelante bajo este tema.

Ejercicio: Preguntas de Pensar Par-Compartir
Comparar y contrastar cómo IgG, IgM e IgE promueven la opsonización. .
Debido a un trastorno de inmunodeficiencia particular, una persona es incapaz de producir C3 convertasa. ¿Cuál de los isotipos de anticuerpos anteriores aún podría participar en la opsonización? Explique brevemente por qué.
Cómo las bacterias resisten la unión a los fagocitos
Como aprendimos anteriormente, algunas bacterias por medio de las actividades que se describen a continuación son capaces de resistir la unión fagocítica:
- Una molécula de membrana externa de Neisseria gonorrhoeae llamada Proteína II y la proteína M de Streptococcus pyogenes permiten que estas bacterias sean más resistentes al engullido fagocítico. La proteína M de S. pyogenes, por ejemplo, se une al factor H de la vía del complemento y esto conduce a la degradación de la opsonina C3b por el factor I y la formación de C3 convertasa.
- Algunas cápsulas simplemente cubren el C3b que sí se une a la superficie bacteriana y evitan que el receptor C3b en los fagocitos entre en contacto con el C3b (Figura\(\PageIndex{11}\)). Esto se observa con la cápsula de Streptococcus pneumoniae.
- Las cápsulas también pueden resistir la unión no mejorada al evitar que los receptores de glicoproteínas en los fagocitos reconozcan los componentes de la pared celular bacteriana y los carbohidratos que contienen manosa.
- S. pneumoniae activa la vía clásica del complemento, pero resiste la opsonización de C3b y el complemento causa más inflamación en los pulmones.
- Neisseria meningitidis tiene una cápsula compuesta por ácido siálico mientras que Streptococcus pyogenes (estreptococos beta del grupo A) tiene una cápsula hecha de ácido hialurónico. Ambos polisacáridos se asemejan mucho a los carbohidratos que se encuentran en los polisacáridos de tejido humano y debido a que no son reconocidos como extraños por los linfocitos que llevan a cabo las respuestas inmunitarias, no se elaboran anticuerpos contra estas cápsulas.
- Algunas bacterias son capaces de cubrirse con proteínas hospedadoras como la fibronectina, la lactoferrina o la transferrina. Esto evita que las moléculas de anticuerpo se unan a epítopos en la superficie bacteriana.
- Staphylococcus aureus produce proteína A mientras que Streptococcus pyogenes produce proteína G. Ambas proteínas se unen a la porción Fc de los anticuerpos, la porción que normalmente se une a los receptores en los fagocitos (Figura\(\PageIndex{12}\)). De esta manera las bacterias se recubren con anticuerpos de una manera que no resulta en opsonización (Figura\(\PageIndex{13}\)).
- Streptococcus pyogenes produce proteínas Mac que son capaces de unirse a los receptores en los fagocitos a los que normalmente se unen IgG y C3b (Figura\(\PageIndex{14}\) y Figura\(\PageIndex{15}\)). Esto bloquea la opsonización.
- Yersinia patógena, como la que causa la plaga, entra en contacto con los fagocitos y, por medio de un sistema de secreción tipo III, libera proteínas que despolimerizan los microfilamentos de actina necesarios para el engullido fagocítico en los fagocitos. Otra proteína Yersinia degrada C3b y C5a.
Resumen
La opsonización, o unión potenciada, se refiere a las moléculas de anticuerpo IgG e IgE, las proteínas del complemento C3b y C4b, y otras opsoninas que unen antígenos a fagocitos. Las porciones Fab del anticuerpo IgG reaccionan con epítopos del antígeno. La porción Fc de IgG puede entonces unirse a neutrófilos y macrófagos pegando así el antígeno al fagocito. La porción Fc de IgA secretora también puede unirse a macrófagos y neutrófilos para opsonización. IgG e IgM pueden activar la ruta clásica del complemento y C3b o C4b pueden pegar el antígeno a los fagocitos. La IgE se elabora contra gusanos parásitos (helmintos) y artrópodos. Las porciones Fab de IgE se unen a epítopos en el helminto o artrópodo mientras que la porción Fc se une a receptores en eosinófilos permitiendo la opsonización.
Preguntas
Estudie el material en esta sección y luego escriba las respuestas a estas preguntas. No solo haga clic en las respuestas y escríbelas. Esto no pondrá a prueba tu comprensión de este tutorial.
- Discutir cómo los anticuerpos defienden el cuerpo a través de la opsonización. (Incluya qué clases o isotipos de inmunoglobulinas están involucradas, el papel de la porción Fab del anticuerpo, el papel, si lo hay, de la porción Fc del anticuerpo, y el papel de cualquier proteína del complemento, si la hubiera, involucrada). (ans)
- Sabemos que Streptococcus pneumoniae está encapsulado y las cápsulas resisten la fagocitosis. Sin embargo, el cuerpo finalmente es capaz de fagocitar a este organismo. Describir cómo. (ans)
- Staphylococcus aureus produce una exotoxina llamada Proteína A. La proteína A es capaz de reaccionar con la porción Fc de IgG. En términos de inmunidad humoral, discuta cómo la Proteína A puede ayudar al Staphylococcus a resistir la fagocitosis. (ans)
- La proteína M de Streptococcus pyogenes se une al factor H de la vía del complemento y permite que estas bacterias sean más resistentes al engullido fagocítico. Explique cómo. (ans)
- Opción múltiple (ans)