La aparición del género Homo marca el advenimiento de cerebros más grandes, el surgimiento de la cultura material (al menos la cultura material que sobrevive en el registro arqueológico) y la eventual colonización del mundo fuera de África. Las primeras especies de Homo son contemporáneas con varios australopitos: Au. africanus, Au. garhi, Au. sediba y todas las especies de Paranthropus. África estaba al ras de homínidos (Figura\(\PageIndex{0}\)). Hay varias tendencias que vemos en la evolución de las primeras especies de Homo hacia nosotros mismos, Homo sapiens:
- Redondeo del cráneo
- Agrandamiento y recableado del cerebro
- Caras y dientes más pequeños
- Prognatismo decreciente
- Tallness
- Diversidad de rasgos culturales
Al igual que con la visión general sobre los primeros homínidos, seguiremos tomando el enfoque de los lumper. Varias especies de Homo no serán discutidas en detalle, pero pueden mencionarse de pasada, por ejemplo, Homo rudolfensis, Homo ergaster, Homo gautengensis, Homo antecessor, Homo cepranensis, Homo rhodesiensis, Homo tsaichangensis. Primero examinaremos las características morfológicas de diversas especies de Homo. Los rasgos culturales se abordarán por separado.

Figura\(\PageIndex{0}\): Modelo de la evolución del género Homo en los últimos 2 millones de años (eje vertical). La rápida expansión “Fuera de África” de H. sapiens se indica en la parte superior del diagrama, con mezcla indicada con neandertales, denisovanos y homínidos africanos arcaicos no especificados. La supervivencia tardía de australopitecinos robustos (Paranthropus) junto al Homo hasta 1.2 Mya se indica en púrpura.
Homo habilis
Homo habilis fue descubierto por primera vez por Louis y Mary Leakey en Olduvai Gorge, Tanzania, en 1960. Asociados con herramientas de piedra (Oldowan), los Leakeys nombraron a su descubrimiento “hombre práctico”. Se han encontrado fósiles de H. habilis en Tanzania, Kenia, Etiopía y Sudáfrica, aunque existe cierto debate sobre si los especímenes de Sudáfrica deben incluirse en la especie. Algunos paleoantropólogos sostienen que hubo otra especie de Homo primitiva, Homo rudolfensis, que se remontan a 2.4mya. Los fósiles de H. rudolfensis son ligeramente más grandes que los de H. habilis, lo que lleva a algunos investigadores a sugerir que H. habilis exhibió dimorfismo sexual y lo que estamos viendo son especímenes masculinos y femeninos de H. habilis. Otros afirman que las diferencias de tamaño son lo suficientemente significativas como para asegurar las dos designaciones de especies (O'Neil c1999-2012). En 2013 se descubrió una mandíbula Homo en el área de investigación Ledi-Geraru, Afar, Etiopía. Con una fecha de 2.8 a 2.75mya, la mandíbula presenta una barbilla similar a Australopithecus y dientes similares a Homo (Villmoare et al. 2015). Si bien aún es temprano en el proceso de investigación, este descubrimiento y otras investigaciones pueden retrasar la fecha del origen del Homo y ayudar a resolver el debate entre los fósiles de H. rudolfensis y H. habilis. Para nuestro propósito, los consideraremos todos H. habilis, haciendo el rango de fechas para este homínido 2.4-1.4mya.
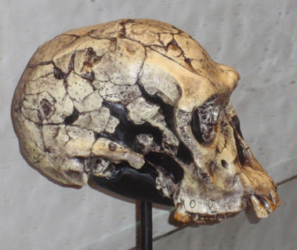
Figura\(\PageIndex{1}\) - Homo habilis
Morfológicamente, H. habilis tiene un cerebro más grande que los australopitos, aproximadamente 35% más grande (O'Neil c1999-2012). Recordarás de la sección sobre tendencias en la evolución humana que el cerebro también comenzó a recablear en este punto. H. erectus presenta menos prognatismo y platicefalia que los homínidos tempranos. La cresta de la ceja también es más pequeña. Todos estos rasgos juntos hacen que el rostro sea más pequeño que los australopitos. Postcranealmente, H. habilis exhibe una mezcla de rasgos primitivos y derivados. Los rasgos primitivos que lo conectan con un antepasado australopital son los antebrazos más largos y el tamaño de los huesos de los dedos junto con la forma en que los tendones se unen a los huesos de la muñeca. Las puntas de los huesos de los dedos son anchas como los humanos. Los dientes más pequeños, una arcada dental en forma de arco parabólico, la morfología del pie y un cráneo más redondeado completan los rasgos humanos. El microanálisis del desgaste dental indica que H. habilis fue omnívoro.
Homo erectus
Con base en datos fósiles actuales, el Homo erectus existió entre 1.9mya a 25 kya (hace mil años) (Jurmain 2013). Eugene Dubois encontró el primer fósil de H. erectus, literalmente “humano recto”, en 1891 en Trinil, Java. También se han encontrado fósiles de H. erectus en África, China, Europa e Israel. Con base en diferencias morfológicas en el cráneo, algunos científicos identifican dos especies, H. erectus en Asia y H. ergaster en África, siendo los especímenes africanos más pequeños que los asiáticos; sin embargo, utilizaremos la designación H. erectus.

Figura\(\PageIndex{2}\) - Homo erectus: Chico Turkana
H. erectus es el primer homínido en alcanzar la altura humana moderna. En 1984, se encontró un esqueleto de H. erectus casi completo a lo largo del río Nariokotome en el norte de Kenia. La datación por potasio-argón coloca a “Turkana Boy” entre 1.64 y 1.33mya. El envejecimiento y el sexado de los restos fósiles indican que el individuo era un macho de aproximadamente ocho años de edad. Él medía alrededor de 5'3” de altura. Si H. erectus siguiera el patrón de crecimiento de los humanos modernos entonces Turkana Boy podría haber alcanzado los 6' como adulto; sin embargo estudios recientes indican que Turkana Boy siguió el patrón de crecimiento de los simios, por lo que habría estado cerca de su estatura adulta en el momento de su muerte (Jurmain et al. 2013).
El patrón de aumento del tamaño cerebral continuó con H. erectus; su cerebro es hasta 50% más grande que su predecesor (O'Neil c1999-2012). Este gran cerebro estaba apoyado por una dieta pesada en carne y otras proteínas. Sus características distintivas incluyen su quilla sagital (un engrosamiento del hueso que va de adelante hacia atrás en la parte superior del cráneo) crestas masivas de cejas (tori supraorbital) y prominencias óseas en la parte posterior del cráneo. Estas prominencias le dan al dorso del cráneo de H. erectus 'una forma de pentágono por detrás. Por un lado, su cráneo tiene forma de fútbol. Postcranealmente, los huesos de H. erectus son más gruesos que H. habilis, al igual que sus mandíbulas y huesos de la cara, y la proporción de brazos a piernas es la de los humanos modernos, lo que hace que algunos sugieran que su andar bípedo era como el nuestro. La longitud de los huesos de sus piernas indica que H. erectus habría sido un eficiente corredor de larga distancia, permitiéndoles “atropellar animales de caza pequeños e incluso medianos en las sabanas tropicales de África Oriental” (O'Neil c1999-2012). Si es así, es probable que H. erectus tuviera mucho menos vello corporal que sus predecesores, ya que habrían necesitado para poder sudar de manera eficiente. Con base en estudios de piojos del cuerpo humano, sabemos que los piojos y los piojos del vello púbico, o piojos de cangrejo, son especies separadas. Los piojos de cangrejo están en realidad más estrechamente relacionados con los piojos del pelo de gorila que los piojos La pérdida de vello corporal creó una barrera entre las dos especies, lo que les permitió a ambas prosperar en los humanos. Los estudios de ADN de las dos especies indican que dividieron 3.0mya, lo que sugiere que los humanos comenzaron a perder el vello corporal en ese momento. Es posible que H. erectus tuviera poco vello corporal (NOVA 2011).
Los científicos generalmente coinciden en que H. erectus fue el primer homínido en abandonar África. Como se mencionó anteriormente, se han encontrado fósiles en África, China, Europa e Israel. En Dmanisi, República de Georgia, se encontraron fósiles en estratos fechados en 1.7mya, lo que sugiere que H. erectus abandonó África poco después de su evolución. Un reporte reciente (Dembo et al. 2015) postula que H. habilis fue el primer homínido en salir de África, no H. erectus. En caso de que esta contención sea apoyada con más datos, aún se puede argumentar que H. erectus tuvo bastante éxito en colonizar el Viejo Mundo, ayudado, sin duda, por sus comportamientos culturales avanzados.
Existe un consenso general de que H. erectus evolucionó a partir de H. habilis y Homo heidelbergensis evolucionó de H. erectus en África, suplantando eventualmente a las poblaciones de H. erectus en el Viejo Mundo (Figura\(\PageIndex{0}\)).
Homo heidelbergensis
Algunas publicaciones, por ejemplo Larsen 2014, se refieren a H. heidelbergensis como el arcaico Homo sapiens, pero para nuestros fines, usaremos la designación H. heidelbergensis. Otto Schoetensack encontró los primeros fósiles de Homo heidelbergensis en 1907 en Mauer, Alemania. Desde entonces se han encontrado fósiles de H. heidelbergensis en África, Europa y Asia. El rango de fechas para la especie es de 800 kya a 350 kya. Los rasgos primitivos incluyen su gran tori supraorbital, hueso frontal bajo, quilla sagital y hueso frontal bajo. Los rasgos derivados incluyen el tori supraorbital separado sobre cada órbita ocular, una bóveda craneal posterior más vertical, huesos parietales anchos en relación con la base craneal y una capacidad craneal mayor que H. erectus (Becoming Human c2008). Adicionalmente, exhiben un dimorfismo sexual similar al de los humanos modernos.

Figura\(\PageIndex{3}\) - Homo heidelbergensis
Existe variación regional en la morfología de H. heidelbergensis. Especímenes europeos encontrados en Atapuerca, (España), Petralona (Grecia), Seinheim (Alemania) y Swancombe (Inglaterra) muestran que tenían cuerpos compactos, lo que podría haber sido una respuesta a vivir en los climas fríos del norte ya que ayudaría a conservar el calor. Adicionalmente, el cráneo es un mosaico de rasgos de H. erectus y rasgos derivados. En Asia, los datos de sitios como Zhoukoudian, Jinniushan y Dalí (China) muestran una mezcla de rasgos de H. erectus y H. sapiens; este último incluye gran capacidad craneal y paredes delgadas del cerebro. Los especímenes africanos de Kabwe (Zambia), Florisbad (Sudáfrica), Laetoli (Tanzania) y Bodo (Etiopía) también muestran una combinación de rasgos de H. erectus y H. sapiens. Comparte el toro supraorbital masivo y el toro occipital prominente con H. erectus y bóveda craneal delgada, occipital menos angulada y base craneal con H. sapiens.
H. heidelbergensis es el ancestro común del Homo neanderthalensis en Europa y del Homo sapiens en África.
Homo neanderthalensis
Numerosos fósiles de neandertales han sido recuperados desde su descubrimiento en 1856 en el Valle de Neander, Alemania. Si bien se desconoce el nombre de la persona que encontró el primer fósil, fue descrito y nombrado por William King. Los neandertales han sido la especulación de los científicos y del público en general desde entonces. Algunos antropólogos clasifican a los neandertales como una subespecie de Homo sapiens, Homo sapiens neanderthalensis, mientras que otros interpretan las diferencias morfológicas como lo suficientemente significativas como para justificar su clasificación como una especie diferente, Homo neanderthalensis. En este ensayo, utilizaremos esta última designación.
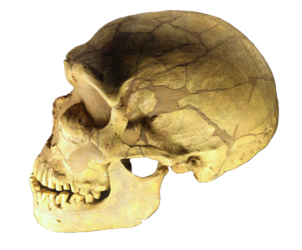
Figura\(\PageIndex{4}\) - Homo neanderthalensis
Los verdaderos neandertales aparecen por primera vez en el registro fósil alrededor de 200 kya, con fósiles que exhiben características similares a los neandertales que aparecen tan pronto como 400 kya. Investigaciones recientes indican que los neandertales se extinguieron entre 41 kya y 39 kya (Higham et al. 2014). La investigación molecular denota que parte del ADN neandertal vive en los humanos modernos, aproximadamente el 2% del ADN de “las personas que descienden de europeos, asiáticos y otros no africanos es neandertal” (Callaway 2014). Los genes neandertales están involucrados en combatir infecciones, lidiar con la radiación ultravioleta (Callaway 2014), y vivir a gran altura (Callaway 2015). El ADN neandertal se ha relacionado recientemente con depresión, obesidad y ciertos trastornos de la piel, por ejemplo, lesiones causadas por la exposición al sol (Callaway 2015).
Los neandertales son el único homínido que originalmente evolucionó en un ambiente glacial, dando lugar a algunas características adaptadas al clima frío. Muchos científicos sostienen que el prognatismo de la mitad de la cara permitió agrandamiento de los senos paranasales que funcionaban para calentar y agregar humedad al aire frío y seco antes de ingresar a los pulmones. Pequeños agujeros debajo de las órbitas oculares, llamados foramina infraorbitaria, son más grandes en los neandertales europeos que los humanos modernos, lo que sugiere que los vasos sanguíneos eran más grandes, lo que permitió un mayor flujo sanguíneo a la cara. Esto habría ayudado a mantener la cara más caliente. Son relativamente cortos y robustos, los machos promedian 5' 5” y las hembras 5'1”, con apéndices más cortos, los cuales ayudarían a conservar el calor al proporcionar menos superficie desde la que irradiar calor. Sus huesos de las piernas son gruesos y densos, lo que sugiere que frecuentemente caminaban y corrían, muy probablemente en actividades de adquisición de alimentos. Algunos postulan que algunos neandertales tenían piel pálida que habría ayudado a aumentar la síntesis de vitamina D al aumentar la cantidad de radiación UV para ser absorbida por el cuerpo (O'Neil c1999-2014). El ancho de sus troncos corporales y las tibias cortas se ajustan a las predicciones para la adaptación al clima frío como propone Christopher Ruff (Larsen 2013). El tamaño cerebral de los neandertales también puede estar relacionado con el clima frío. Con un promedio de una capacidad craneal entre 1300 y 1400 cm3, tienen los cerebros más grandes de todos los homínidos, incluyendo H. sapiens. El tamaño puede estar asociado con una mayor eficiencia metabólica en clima frío, lo cual es similar a los pueblos inuit modernos de hoy que tienen un tamaño cerebral mayor que otras poblaciones humanas (Jurmain et al. 2013).
La presencia de un moño occipital es una de las características utilizadas para identificar especímenes de H. neanderthalensis, aunque cabe señalar que estas características persisten en un pequeño porcentaje de las poblaciones humanas modernas. El moño occipital puede haber evolucionado para contrarrestar la cara pesada de Neanderthal al correr; evita que la cabeza haga enormes aceleraciones horizontales (NOVA 2002). El moño occipital también alarga el cráneo en comparación con H. sapiens. Tenían fuertes crestas de cejas como las de H. heidelbergensis. Los cráneos neandertales recuperados de Amud y Tabún en Israel presentan más características craneales similares a H. sapiens, incluyendo falta de moño occipital, órbitas oculares más pequeñas, aberturas nasales altas y anchas y dientes más pequeños (Larsen 2014).
Denisovanos
En 2010, los científicos anunciaron el descubrimiento y análisis de ADN de un hueso de dedo y dos dientes encontrados en la cueva Denisova, Siberia (Reich et al. 2010). Los artefactos fueron recuperados de un depósito fechado de 50 kya a 30 kya. Los datos sugieren que los restos eran de un individuo que compartía un origen común con los neandertales, pero no era un neandertal, ni era un humano moderno. Los individuos denisovanos comparten 4-6% de su material genético con pueblos modernos que viven en Nueva Guinea, Islas Bougainville y China. Otros estudios (Cooper y Stringer 2013) indican que los denisovanos cruzaron Eurasia y se cruzaron con humanos modernos, pero genéticamente los denisovanos estaban más estrechamente relacionados con los neandertales que con los humanos modernos (Meyer et al. 2012). Si bien otros hallazgos arrojarán más luz sobre los denisovanos, es claro que hubo más variabilidad genética durante el Pleistoceno de lo que se pensaba anteriormente (Larsen 2014), pero que el flujo de genes entre las diversas poblaciones tuvo implicaciones para la emergencia de los humanos modernos (Pääbo 2015).
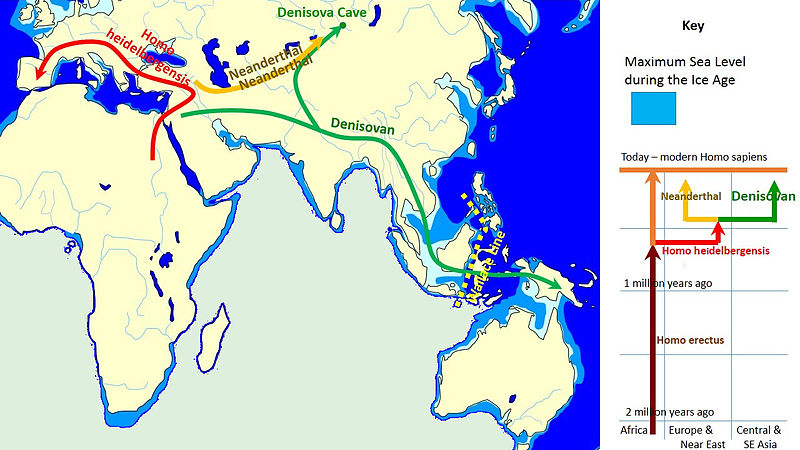
Figura\(\PageIndex{5}\) - Difusión y evolución de los denisovanos
Homo floresiensis
En 2003, Peter Brown y Michael Morwood lideraron un equipo que descubrió fósiles inusuales en la cueva de Liang Bua, Isla Flores, Indonesia. La pequeña estatura del individuo condujo al nombramiento de una nueva especie, Homo floresiensis. Desde entonces se han encontrado doce individuos que van en el tiempo de 74 kya a 17 kya. H. floresiensis tenía un cerebro pequeño y solo tenía cerca de 3.5' de altura cuando era adulto. Tenían frentes retrocedidas, sin barbas, hombros encogidos hacia adelante y pies grandes en relación con sus piernas cortas. Comparten algunas características con H. sapiens, incluyendo dentición más pequeña, crestas de cejas separadas y una cara no prognática.

Figura\(\PageIndex{6}\) - Homo floresiensis
Se han propuesto varias hipótesis para explicar la aparición de H. floresiensis. Una hipótesis sugiere que estamos viendo enanismo insular, que es un proceso evolutivo resultante del aislamiento a largo plazo en una isla pequeña. Recursos limitados y falta de depredadores seleccionados para individuos de cuerpo más pequeño que necesitan menos recursos que los individuos de cuerpo grande. Otra hipótesis afirma que H. floresiensis no es una especie separada, sino que H. sapiens exhibe microcefalia o alguna otra deficiencia del desarrollo como el cretinismo hipertiroideo (Oxnard et al. 2010) porque los “... rasgos craneales... están dentro del rango moderno de variación observada en poblaciones vivas de la región más grande [de Indonesia]” (Larsen 2014:409), dando apoyo a la segunda hipótesis. La forma en que H. floresiensis encaja en el panorama evolutivo no está claro con base en los datos actuales. Una visión general de las diversas hipótesis sobre sus relaciones evolutivas en el artículo Homo floresiensis: Making Sense of the Smalled Hominin Fossils from Flores.
Homo sapiens
En la actualidad, solo una especie de homínidos habita en el planeta Tierra, el Homo sapiens. Morfológicamente, las características de H. sapiens incluyen la presencia de un mentón (o prominencia mental), un cerebro grande, cara plana, cráneo redondeado o globular, y una cresta ceja continua y reducida. Sus huesos son graciles en comparación con los homínidos anteriores, aunque los primeros H. sapiens son más robustos que las poblaciones modernas y no muestran ninguna de las adaptaciones de clima frío de los neandertales. La naturaleza más gracile da crédito a la hipótesis de que los humanos modernos evolucionaron primero en África. Las proporciones corporales más delgadas son más adaptativas a los ambientes tropicales africanos ya que habría más superficie corporal para irradiar calor. La evidencia de ADN también respalda un origen africano. Las poblaciones africanas modernas tienen más diversidad genética que cualquier otra población humana moderna, inferyendo que han ido evolucionando más tiempo (Becoming Human c2008).
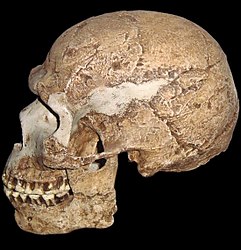
Figura\(\PageIndex{7}\) - Homo sapiens
Los fósiles más antiguos de H. sapiens se encontraron en África en Omo Kibish, Etiopía, fechados en 195 kya. Los fósiles de H. sapiens encontrados en Herto, Etiopía (160 kya—154 kya), la boca del río Klasies y la cueva fronteriza, Sudáfrica (120 kya — 89 kya), Skhūl, Israel (130 kya—100 kya), y Qafzeh, Isreal (120 kya—92 kya), agregan más apoyo. Sitios en Asia, e.g., Zhoukoudian y Tianyuan en China y Niah Cave, Borneo, Indonesia datan de 40 kya o menos y en Europa que datan no antes de 31 kya (Mladeč, República Checa). Los humanos modernos llegaron a Australia por 55 kya; sin embargo, no se han encontrado restos humanos en el continente que datan antes de 30 kya en el lago Mungo. Estos datos dan poco apoyo a la Hipótesis Multirregional, un modelo desarrollado por Milford Wolpoff que propuso que el H. sapiens moderno evolucionó en lugares separados del Viejo Mundo a partir de poblaciones locales arcaicas de H. sapiens. Este modelo sugirió que el flujo de genes entre las poblaciones condujo a la similitud genética de los humanos modernos. La mayoría de los antropólogos apoyan el modelo de origen africano. Hay dos variantes de este modelo. El modelo Out of Africa, propuesto por Chris Stringer, afirma que los humanos modernos surgieron en África y se extendieron a Europa después de 50 kya, reemplazando a la población no H. sapiens sin flujo génico. Fred Smith y Erik Trinkhaus propusieron el modelo de Asimilación que también afirmó que el humano moderno surgió por primera vez en África y se extendió a Europa y Asia. La principal diferencia entre los modelos Out of Africa y Asimilation es que este último afirma flujo génico entre H. sapiens y H. neanderthalensis. Teniendo en cuenta los recientes análisis de ADN de neandertales discutidos anteriormente, múltiples líneas de evidencia apoyan el modelo de Asimilación.
El caso de los neandertales en desaparición
Sabemos que algunos genes neandertales están en el genoma de los humanos modernos, por lo que en esencia, alguna parte de los neandertales sobrevive hoy. Sin embargo, no hay neandertales caminando en el mundo moderno. Entonces, ¿qué pasó con los neandertales? Los científicos sugieren que H. sapiens superó a los neandertales con su dieta más diversa y “habilidades sofisticadas y cognitivas” (Becoming Human c2008). Estos rasgos permitieron a H. sapiens adaptarse fácilmente a las condiciones climáticas rápidamente cambiantes durante el Paleolítico Superior.
Una hipótesis reciente sugiere que los neandertales se extinguieron durante un período de poderosa actividad volcánica en Eurasia occidental. Las excavaciones en la cueva Mezmaiskaya en las montañas del Cáucaso en el sur de Rusia han recuperado una gran cantidad de huesos neandertales, herramientas de piedra y restos de animales presa. Los restos y artefactos neandertales aparecen en estratos por encima de una capa de ceniza volcánica y debajo de una segunda capa de ceniza volcánica. No se han encontrado huesos ni artefactos neandertales por encima del segundo nivel, lo que sugiere que los neandertales ya no ocuparon la zona. Ambas capas de ceniza contienen niveles de polen asociados con climas más fríos y secos. “Las capas de ceniza corresponden cronológicamente a lo que se conoce como la súper erupción campaniana de Ignimbrita, que ocurrió hace alrededor de 40 mil años en la Italia moderna, y una erupción más pequeña que se cree que ocurrió alrededor de la misma época en las montañas del Cáucaso” (University of Chicago Press Journals 2010). El invierno volcánico subsiguiente provocó un cambio climático dramático que llevó a la desaparición de los neandertales. Estos datos se correlacionan con la investigación de Higham et al. (2014) que afirma que los neandertales se extinguieron entre 41 kya y 39 kya.
Referencias
- Volviéndose Humano. c2008. El linaje humano a través del tiempo. Instituto de Orígenes Humanos [Internet] [citado 2015 Ago 3]. Disponible en: http://www.becominghuman.org/node/human-lineage-through-time
- Callaway E. 2014. Los genomas humanos modernos revelan nuestro Neandertal interior. Naturaleza [Internet] [citado 2015 Ago 21]. Disponible en: http://www.nature.com/news/modern-hu...erthal-1.14615
- Callaway E. 2015. Los neandertales tuvieron un efecto enorme en la biología humana. Naturaleza [Internet] [citado el 21 de ago de 2015]; 523 (7562): 512-513. Disponible en: http://www.nature.com.offcampus.lib....iology-1.18086. doi: 10.1038/523512a
- Cooper A, Stringer CB. 2013. ¿Los denisovanos cruzaron la Línea Wallace? Ciencia [Internet] [citado 2015 Ago 21}; 342 (6156): 321-323. Disponible en: http://www.sciencemag.org.offcampus....d-963a99b700ea. doi: 10.1126/ciencia.1244869.
- Dembo M, Matzke NJ, Mooers AØ, Collard M. 2015. El análisis bayesiano de una supermatriz morfológica arroja luz sobre las controvertidas relaciones con los homínidos. Proc R Soc B [Internet] [citado el 20 de ago de 2015]; 282 (1812). Disponible en: http://rspb.royalsocietypublishing.o...0943.e-letters. doi: 10.1098/rspb.2015.0943.
- eFósiles [Internet] [citado 2015 ago 10]. Departamento de Antropología, La Universidad de Texas en Austin. Disponible en: http://efossils.org/
- Higham T, Douka K, Wood R et al. 2014. El momento y el patrón espaciotemporal de la desaparición neandertal. Naturaleza [Internet] [citado el 21 de ago de 2015]; 512 (7514): 306-309. Disponible en: http://www.nature.com.offcampus.lib....ture13621.html. doi:10.1038/naturaleza13621
- Jurmain R, Kilgore L. Trevathan W. 2013 Esenciales en antropología física. Belmont (CA): Wadsworth Cengage Learning.
- Larsen, CS. 2014. Nuestros orígenes: descubrir la antropología física. Nueva York (NY): W. W. Norton & Company, Inc.
- Piojos y evolución humana [Internet]. 2011 Feb 6. NovaCencienow. [citado 2015 Ago 20]. Disponible en: www.pbs.org/wgbh/nova/evolution/lice.html
- Meyer M, Kircher M, Gansauge MT, et al. 2012. Secuencia genómica de alta cobertura de un individuo denisovano arcaico. Ciencia [Internet] [citado el 21 de ago de 2015]; 338 (6104): 222-226. Disponible en: http://www.sciencemag.org.offcampus..../6104/222.full. doi: 10.1126/ciencia.1224344.
- NOVA [Internet]. 2002 Ene 22. Neandertales en juicio. PBS [citado 2015 Ago 21]. Disponible en: http://www.pbs.org/wgbh/nova/transcr...nderthals.html
- O'Neil D. c1999-2012. Evolución humana temprana: un estudio de la evolución biológica y cultural de Homo habilis y Homo erectus. Departamento de Ciencias del Comportamiento, Colegio Palomar [Internet] [citado 2015 Ago 16]. Disponible en: Anthro.palomar.edu/Homo/Default.htm
- O'Neil D. c1999-2014. Evolución de los humanos modernos: un estudio de la evolución biológica y cultural del Homo sapiens arcaico y moderno. Departamento de Ciencias del Comportamiento, Colegio Palomar [Internet] [citado 2015 Ago 16]. Disponible en: anthro.palomar.edu/homo2/default.htm
- Oxnard C, Oberendorf PJ, Kefford BJ. 2010. Los esqueletos postcraneales de cretinas hipertiroideas muestran un mosaico anatómico similar al Homo floesiensis. PLoS One [Internet] [citado el 21 de ago de 2015]; 5 (9). Disponible en: http://www.sciencedaily.com/releases...0928025514.htm. doi: 10.1371/journal.pone.0013018.
- Pääbo S. 2015. Los diversos orígenes del acervo genético humano. Nat Rev Genet [Internet] [citado 2015 Ago 21]; 16:313-314. Disponible en: http://www.nature.com.offcampus.lib....l/nrg3954.html. doi:10.1038/nrg3954
- Reich D, Green RE, Kircher M, et al. 2010. Historia genética de un grupo arcaico de homínidos de la cueva Denisova en Siberia. Naturaleza [Internet] [citado el 21 de ago de 2015]; 468 (7327). Disponible en: http://www.nature.com/nature/journal...ture09710.html
- Institución Smithsonian [Internet]. 2015 ago 4. ¿Qué significa ser humano? [citado 2015 Ago 11]. Disponible en: http://humanorigins.si.edu/evidence/human-fossils/species
- Revistas de prensa de la Universidad de Chicago [Internet]. Los volcanes aniquilaron a los neandertales, sugiere un nuevo estudio. ScienceDaily 2010 Oct 7 [citado 2015 Ago 21]. Disponible en: http://www.sciencedaily.com/releases...1006094057.htm
- Villmoare B, Kimbel WH, Seyoum C, Campisano CJ, DiMaggio EN, Rowan J, Braun DR, Arrowsmith JR, Reed KE. 2015. Homo temprano a 2.8 MA de Ledi-Geraru, Afar, Etiopía. Ciencia [Internet]. [citado 2015 Ago 16]; 347 (6228): 1352-1355. Disponible en: http://www.sciencemag.org.offcampus....6228/1352.full. doi: 10.1126/ciencia.aaa1343

