
16.3: Pelea o huida
- Page ID
- 148198
- Compara diferentes teorías sobre la generación de emociones.
- Describir las bases anatómicas y químicas de la ira y el miedo.
- Analizar cómo la investigación epigenética ha mejorado nuestra comprensión del debate sobre la crianza de la naturaleza en torno a la ira y el miedo.
Visión general
Una función importante de las emociones en los seres humanos, y otros animales que viven en grupos de cualquier tipo, es comunicarse con otros miembros de la propia especie. Las emociones que son particularmente importantes para comunicar a otros miembros son las relacionadas con el miedo, y las relacionadas con la ira. Cuando pensamos en la excitación fisiológica asociada a las emociones, a menudo hablamos de la respuesta de “lucha o huida”. Estas se corresponden muy bien con las dos emociones negativas principales: la ira y el miedo. Múltiples estudios de paradigmas animales e investigaciones neurocientíficas humanas, incluyendo estudios con condiciones psicopatológicas, han servido para examinar la naturaleza de estas emociones. Se discutirán algunas de estas teorías y hallazgos.
Teorías de las Distinciones Emocionales
¿Pueden describirse mejor las emociones como cualitativamente distintas, por ejemplo, como “emociones básicas” discretas o “clases naturales” (Ekman et al., 1983; Izard, 1992; Panksepp, 2005) o como cuantitativamente distintas, por ejemplo, como puntos a lo largo de una circunplexión definido por dimensiones como excitación y valencia (Russell y Barrett, 1999; Barrett y Wager, 2006)? En los últimos años se ha observado un prolongado debate en la literatura sobre cómo capturar con mayor precisión la naturaleza de la emoción (Barrett et al., 2007; Izard, 2007; Panksepp, 2007; Tracy y Randles, 2011), con modelos propuestos de emoción que incluyen no solo básicos modelos emocionales y dimensionales, pero también aquellos que se enfocan en evaluaciones relevantes para objetivos de estímulos emocionales (Moors et al., 2013), emociones como respuestas de afrontamiento (Roseman, 2013) y emociones como circuitos de supervivencia (LeDoux, 2012). Una conversación extendida sobre las fortalezas y debilidades de estos diversos puntos de vista no se revisará en su totalidad aquí, más bien, la atención se centrará en la consideración básica de si las diferentes emociones (por ejemplo, miedo, ira) son mejor vistas como cualitativa o cuantitativamente distintas.
Ideas Cualitativas

Los modelos que postulan las emociones como cualitativamente distintas, como los modelos de “emoción básica”, sostienen que un número limitado de emociones como el miedo, la ira y la excitación positiva emergen de procesos neurofisiológicos disociables (Ekman et al., 1983; Izard, 1992; Panksepp, 2005 ; Lench et al., 2011). En Figura\(\PageIndex{1}\) vemos una representación de esta idea de que inicialmente tenemos emociones básicas entre ellas el miedo, la ira, el asco, el desprecio, la alegría, la tristeza y el interés, que luego crecen para incluir emociones autoconscientes (culpa, orgullo, vergüenza, triunfo) y finalmente emociones cognitivamente complejas (envidia, gratitud, decepción, arrepentimiento, esperanza, schadenfreude, empatía, compasión). Estos procesos neurofisiológicos generalmente están vinculados a la actividad en las estructuras subcorticales evolutivamente antiguas del mesencéfalo, el cuerpo estriado y el sistema límbico más comúnmente vinculados a la emoción (Panksepp, 2005; Vytal y Hamann, 2010). Entonces, por ejemplo, la generación de excitación positiva está ligada a la activación en un circuito estriatal centrado en las neuronas dopaminérgicas en el núcleo accumbens (Ikemoto y Panksepp, 1999), mientras que la generación de miedo se asocia con la actividad en un circuito que involucra el gris periacueductal, hipotálamo anterior y medial, y amígdala (LeDoux, 2000). En esta visión, las gradaciones más finas de la experiencia resultan cuando las emociones básicas son moduladas o elaboradas por procesos cognitivos de nivel superior controlados por la corteza cerebral, pero la aparición de emociones cualitativamente distintas no depende de estos procesos controlados corticalmente (Panksepp, 2005).
Ideas Cuantitativas
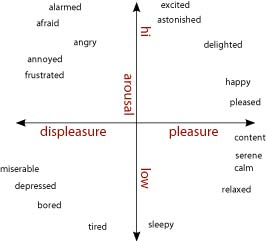
Los modelos que postulan que las emociones son cuantitativamente distintas sostienen que emociones como el miedo, la ira y la felicidad se describen mejor como puntos en una o más dimensiones centrales. Las dimensiones centrales que se proponen típicamente para distinguir entre las emociones son la excitación o activación fisiológica (baja-alta) y la valencia (mala-buena) (Bradley et al., 2001). [Algunos han propuesto una dimensión de enfoque de retiro como sustituto o complemento del eje de valencia (Wager et al., 2003; Christie y Friedman, 2004; van Honk y Schutter, 2006)]. Como se muestra en la figura\(\PageIndex{2}\), dispuestas ortogonalmente, estas dimensiones forman un circumplex sobre el cual las emociones pueden ser trazadas y comparadas cuantitativamente (Barrett y Russell, 1999; Russell y Barrett, 1999; Colibazzi et al., 2010). La excitación positiva se traza como alta en excitación y positiva en valencia, y la tristeza es baja en excitación y negativa en valencia. El miedo se suele trazar como excitación alta y fuertemente negativo, al igual que la ira (Russell y Barrett, 1999). Se piensa que otras distinciones entre las emociones reflejan diferencias en las construcciones cognitivas de los eventos que rodean los cambios básicos en la excitación y la valencia. Así, si un individuo experimenta ira o miedo (que son similares en términos de excitación o valencia) puede ser moldeado por interpretaciones de cambios neurofisiológicos en valencia y excitación a la luz del estímulo provocador y las reservas idiosincrásicas del individuo de conocimiento semántico, recuerdos y respuestas conductuales que dan forma al estado subjetivamente experimentado (Russell, 2003). Bajo esta visión, las distinciones entre los estados emocionales experimentados son altamente dependientes de estos procesos cognitivamente complejos, los cuales son subatendidos por una red distribuida de regiones de la corteza cerebral (Lindquist et al., 2012).
Estos modelos generan distintas predicciones sobre la cuestión de si un trastorno o lesión podría resultar en que una sola emoción se desactive sin afectar la experiencia de otras emociones. La visión de las emociones discretas argumentaría que un trastorno o lesión que resultara en disfunción en las estructuras específicas que sufrían a una emoción particular podría afectar la experiencia de una emoción mientras dejaba intactas a otras. En contraste, la visión dimensional requeriría que otras emociones que son dimensionalmente similares a la emoción afectada también se vean afectadas, o que los déficits en una emoción particular reflejen disfunción en procesos cognitivos de nivel superior impulsados corticalmente.
La psicopatía apoya el modelo cualitativo
El caso de la psicopatía da un claro apoyo a la noción de que el miedo es cualitativamente distinto de otras emociones. En psicopatía, la mayor parte de la evidencia clínica y empírica apunta hacia la conclusión de que el miedo a responder está inhabilitado de manera única, con otras emociones de alta excitación (excitación positiva, ira) y negativamente valenciadas (ira, disgusto) permaneciendo intactas. La visión dimensional no puede explicar fácilmente por qué en los psicópatas se genera fácilmente (quizás con demasiada facilidad) el estado de ira de alta excitación, negativamente valenciado, mientras que el estado de miedo de alta excitación, negativamente valenciado no lo es. El problema no puede estar en una incapacidad para involucrar completamente a los sistemas neurocognitivos subyacentes ya sea en la dimensión de excitación o valencia, porque los psicópatas experimentan otras emociones de alta excitación (excitación positiva) así como otras emociones valenciadas negativamente (disgusto). Tampoco puede resultar de alguna dificultad que surge en la interacción de estos ejes, porque la ira y el miedo son muy similares en términos de ambas dimensiones. Los modelos que sustituyen un eje de retiro-enfoque por un eje negativo-positivo no tienen más éxito; las dos emociones más fuertemente vinculadas a la retirada son el asco y el miedo, y no hay evidencia de deficiencias basadas en asquerosos en la psicopatía. Los individuos con psicopatía tampoco logran reconocer y por lo tanto no tienen una respuesta empática al miedo de los demás.
En conjunto, los datos empíricos sustentan la idea de que la amígdala, junto con sus proyecciones eferentes, es una estructura esencial para la generación de respuestas de miedo condicionadas, que dan cuenta de la mayoría del miedo experimentado (Davis, 1992, 1997). Amplia evidencia temprana demostró que la amígdala juega un papel crucial en la creación de miedo condicionado en roedores. Por ejemplo, las lesiones a la amígdala impiden que las ratas desarrollen una respuesta de miedo condicionada, como la congelación en respuesta a un estímulo que predice el choque (Blanchard y Blanchard, 1972). Estudios posteriores aclararon los papeles de los diversos subnúcleos de la amígdala, demostrando que el núcleo lateral está principalmente involucrado en la adquisición de la respuesta al miedo mientras que el núcleo central está involucrado tanto en la adquisición como en la expresión de respuestas condicionadas al miedo (Davis, 1992; Wilensky et al., 2006). Las múltiples proyecciones eferentes de la amígdala coordinan las respuestas autonómicas y conductuales al miedo que provoca estímulos. Las proyecciones desde el núcleo central de la amígdala hasta el hipotálamo lateral están involucradas en la activación de las respuestas del sistema nervioso simpático autónomo, y las proyecciones al gris periacueductal ventrolateral dirigen la expresión de respuestas conductuales, como la congelación defensiva (Davis, 1992; LeDoux, 2012). El papel central de la amígdala en la respuesta coordinada al miedo puede demostrarse mediante estudios de estimulación eléctrica que muestran que patrones complejos de cambios conductuales y autonómicos asociados con las respuestas al miedo son el resultado de la estimulación de las regiones relevantes de la amígdala (Davis, 1992). La fuerte dependencia de modelos animales está justificada en el estudio de la respuesta al miedo y la amígdala dada la gran conservación de los núcleos de amígdala involucrados en la respuesta a amenazas condicionadas en todas las especies que van desde reptiles hasta aves, roedores y primates (LeDoux, 2012).
Investigación que sugiere conexiones entre ira y miedo
Neumann et al. (2010) plantean la hipótesis de que los comportamientos agresivos, que son de dos tipos básicos, reactivos y proactivos, están mediados por bases neurológicas basadas en la ansiedad. Con base en modelos animales, sugieren lo siguiente: “La agresión masculina es necesaria para la adquisición y mantenimiento de la nutrición, el territorio y las parejas de apareamiento. Las reglas específicas de la especie deben ser estrictamente obedecidas para garantizar una comunicación efectiva e inofensiva. Así, la agresión ofensiva adaptativa está compuesta principalmente por comportamientos de amenaza inofensivos que permiten al oponente escapar o cambiar a comportamientos sumisos para evitar la confrontación física directa. En roedores, tales signos de agresión ofensiva incluyen piloerección (intimidación del oponente por mayor apariencia) y amenaza lateral (espalda arqueada y exposición del flanco). En caso de un ataque ofensivo, partes del cuerpo menos vulnerables del oponente, como las cubiertas de músculos y una gruesa capa de piel, son apuntadas para evitar lesiones graves (Blanchard y Blanchard, 1977; Blanchard et al., 2003). Si bien la agresión ofensiva generalmente se expresa durante una lucha por territorio o apareamiento exclusivo, la agresión defensiva se muestra principalmente en situaciones potencialmente mortales y está vinculada a un aumento del miedo (Blanchard y Blanchard, 1981). A diferencia de la agresión ofensiva, la agresión defensiva es menor o no señalada de antemano, y los objetivos de ataque incluyen partes del cuerpo más vulnerables (como la cabeza, el vientre y los genitales) (Blanchard y Blanchard, 1977; Blanchard et al., 2003).” Los paradigmas animales residente-intruso han sido utilizados para medir estas ideas (ver Figura\(\PageIndex{3}\)).

También afirman: “La ansiedad puede interpretarse como una anticipación emocional de una situación aversiva y se refleja en las respuestas de miedo conductual específicas de la especie a estímulos estresantes y amenazantes característicos de la ansiedad de rasgo individual. El miedo no se ve como estado basal (como lo es la ansiedad), sino como una respuesta conductual compleja, como el sobresalto o la congelación. Además de los factores que determinan la ansiedad innata (rasgo), varios factores ambientales o farmacológicos pueden interactuar con el fondo genético y determinar el nivel individual de ansiedad de estado y el fenotipo conductual final. La emocionalidad, a menudo utilizada como sinónimo de ansiedad y miedo, puede verse en un sentido más amplio, que comprende tanto la ansiedad de rasgo como del estado y el miedo relacionado con el estímulo. La emocionalidad es uno de los principales componentes subyacentes a la capacidad de un organismo para evaluar estímulos y escenarios estresantes, y para afrontarlos adecuadamente”. (Neumann et al., 2010)
Neumann et al. (2010) muestran que varios estudios clínicos y de laboratorio con humanos y roedores han demostrado que existen mecanismos complicados por los cuales se corregulan la ansiedad y la agresión. Hay varios neuroquímicos que se encuentran involucrados de formas complejas de la siguiente manera:
Glucocorticoides - La regulación del eje HPA (Hipotalámico-Pituitario-Adrenal) parece muy conectada con la experiencia de ansiedad y agresión. Sin embargo, la dirección no es sencilla, ya que tanto los glucocorticoides altos como los bajos están relacionados con una alta agresión.
Vasopresina - La vasopresina se produce en el hipotálamo, y la testosterona se produce en las gónadas, y ambas modulan la agresión así como la ansiedad en los machos. La vasopresina también está involucrada en la reducción de la ansiedad, así como en la vinculación de parejas.
Testosterona - Junto con la agresión, la testosterona aumenta en la pubertad, la castración reduce ambas (a través de la reducción en Vasopresina también). Existen algunos factores ambientales que tienen un efecto moderador, pero en general, la testosterona está claramente relacionada con la modulación de la agresión en los varones.
Serotonina - menor 5-HT parece estar ligada a una mayor agresión y violencia en rhesus femenino. Los factores genéticos y los factores estresantes de la vida temprana tienden a mediar estos efectos.
GABA -cuando se incrementan los niveles de ansiedad, la agresión aumenta dependiendo de las predisposiciones genéticas al mismo, y la actividad también aumenta en “regiones cerebrales incluyendo la amígdala central, BST (Núcleo de lecho de la estría terminal), tabique lateral, y NPV (núcleo paraventricular) que se asocian con estrés-, comportamiento relacionado con el miedo y la agresión”.
“La sorprendente evidencia de un solapamiento en los sistemas neuroendocrino y neuroquímico que regulan la agresión así como la ansiedad sugiere una fuerte correlación entre estos dos comportamientos. Así, la agresión y la ansiedad no siempre están co-reguladas, pero, en algunas circunstancias, estos comportamientos pueden estar bajo el control de los mismos genes y sustancias neuroactivas incluyendo esteroides sexuales, neuropéptidos y neuroaminas dentro de circuitos cerebrales específicos. Tal visión concuerda con los hallazgos clínicos. Por un lado, se observan comportamientos excesivos y violentos en humanos expuestos a experiencias adversas tempranas de la vida y en pacientes con trastornos relacionados con depresión y ansiedad, o TEPT. Por otro lado, existen datos bastante contradictorios sobre los efectos de los fármacos ansiolíticos en conductas antisociales y agresivas. En futuros estudios que se centren en los mecanismos neurobiológicos de (co) regulación de la agresión y la ansiedad, es necesario considerar modificaciones epigenéticas además de los parámetros neuronales y neuroendocrinos discutidos anteriormente”. (Neumann et al, 2010).
Naturaleza v. Nutrir
Tanto los factores genéticos como las experiencias juegan un papel en las conductas agresivas. La regulación genética de la liberación de serotonina parece desempeñar un papel en la agresión en muchos mamíferos, incluidos los humanos. Por ejemplo, Peeters et al. ' s (2020) concluyó que la versión corta del gen transportador de serotonina (alelo S del 5-HTTLPR) parece estar vinculada a una mayor agresión reactiva, y aquellos individuos con mayores tendencias de evitación hacia expresiones faciales enojadas. Sus hallazgos indican que los impulsos evaluativos en respuesta a las señales sociales juegan un papel importante en la mediación de la predisposición genética del polimorfismo 5-HTTLPR a una mayor expresión de agresión reactiva. Debido al papel principal de los MAO en el metabolismo de los neurotransmisores clave involucrados en el comportamiento agresivo, sobre todo la serotonina, no es sorprendente que un cuerpo sustancial de investigación haya vinculado fenotipos agresivos con el sistema MAO. Muchos resultados inspiraron tanto a la comunidad científica como a los medios de comunicación a referirse al MAO-A como el gen guerrero o criminal. Aunque los estudios farmacológicos iniciales apoyaron un papel de los IMAIS en la reducción de fenotipos agresivos, los datos de estos estudios fueron difíciles de interpretar debido a los efectos secundarios de los IMAIS y por su impacto en una miríada de comportamientos no relacionados. La evidencia sustantiva que respalda el papel de los genes MAO en la agresión proviene de estudios en ratones knockout (KO). Los modelos de knockout selectivos para el gen MAO-A, por ejemplo, mostraron una mayor agresividad en comparación con sus homólogos de tipo silvestre. Los estudios genéticos en humanos proporcionan evidencia de que la MAO-A está vinculada a la agresión pero solo cuando otros factores ambientales también están presentes durante el desarrollo (por ejemplo, abuso, factores estresantes). Otras líneas de investigación han demostrado que el producto génico de MAO-A (más que el gen per se) influye en rasgos violentos. Por ejemplo, la actividad de MAO-A cortical y subcortical in vivo -medida con tomografía por emisión de positrones (PET) - se asoció negativamente con la agresión de rasgos (atribución: Mentis, Dardiotis, Katsouni & Chrousos, 2021).
En las últimas dos décadas, el estudio de la relación entre la naturaleza y la crianza en la conformación del comportamiento humano ha encontrado un interés renovado. La genética conductual mostró que distintos polimorfismos de genes que codifican proteínas que controlan la función metabólica y sináptica de los neurotransmisores están asociados con la vulnerabilidad individual a experiencias aversivas, como eventos vitales estresantes y traumáticos, y pueden resultar en un mayor riesgo de desarrollar psicopatologías asociadas a la violencia. Por otro lado, estudios recientes indican que experimentar eventos aversivos modula la expresión génica al introducir cambios estables en el ADN sin modificar su secuencia, mecanismo conocido como “epigenética”. Por ejemplo, experimentar adversidades durante periodos de máxima sensibilidad al ambiente, como la vida prenatal, la infancia y la adolescencia temprana, puede introducir marcas epigenéticas duraderas en genes que afectan los procesos maduracionales en el cerebro, favoreciendo así la aparición de conductas disfuncionales, entre ellas exagerar la agresión en la edad adulta.
Las influencias ambientales adversas durante los períodos críticos de desarrollo se han correlacionado con marcadores epigenéticos que afectan la función del receptor de glucocorticoides. Como se ve en muchos estudios, la baja liberación de cortisol se correlaciona con un bajo autocontrol y una mayor agresión. La producción de oxitocina se relaciona con un mayor funcionamiento social y formación de apego. La secreción de oxitocina es estimulada por la atención materna temprana, mientras que las experiencias adversas en el desarrollo prenatal y temprano de la infancia se relacionan con una menor metilación del receptor de oxitocina (un proceso involucrado en la epigenética). La serotonina está relacionada con la regulación de la agresión. Los genes responsables de controlar la recaptación de serotonina de la hendidura sináptica se ven afectados por factores estresantes tempranos (incluyendo traumatismos así como condiciones generadas por la pobreza como la mala atención médica, la calidad de la vivienda y la exposición a barrios violentos) que alteran la anatomía y función cerebral como la cortical espesor y reactividad amígdala. (Ver Figura\(\PageIndex{4}\) para conocer las múltiples formas en que el estrés y los traumas conducen a modificaciones epigenéticas en los sistemas de ejes de oxitocina, serotonina y HPA que afectan el comportamiento agresivo: agresión reactiva y proactiva, hostilidad, problemas de externalización de la delincuencia, violencia, trastornos de conducta, física y agresión verbal y rasgos indecibles sin emociones.)
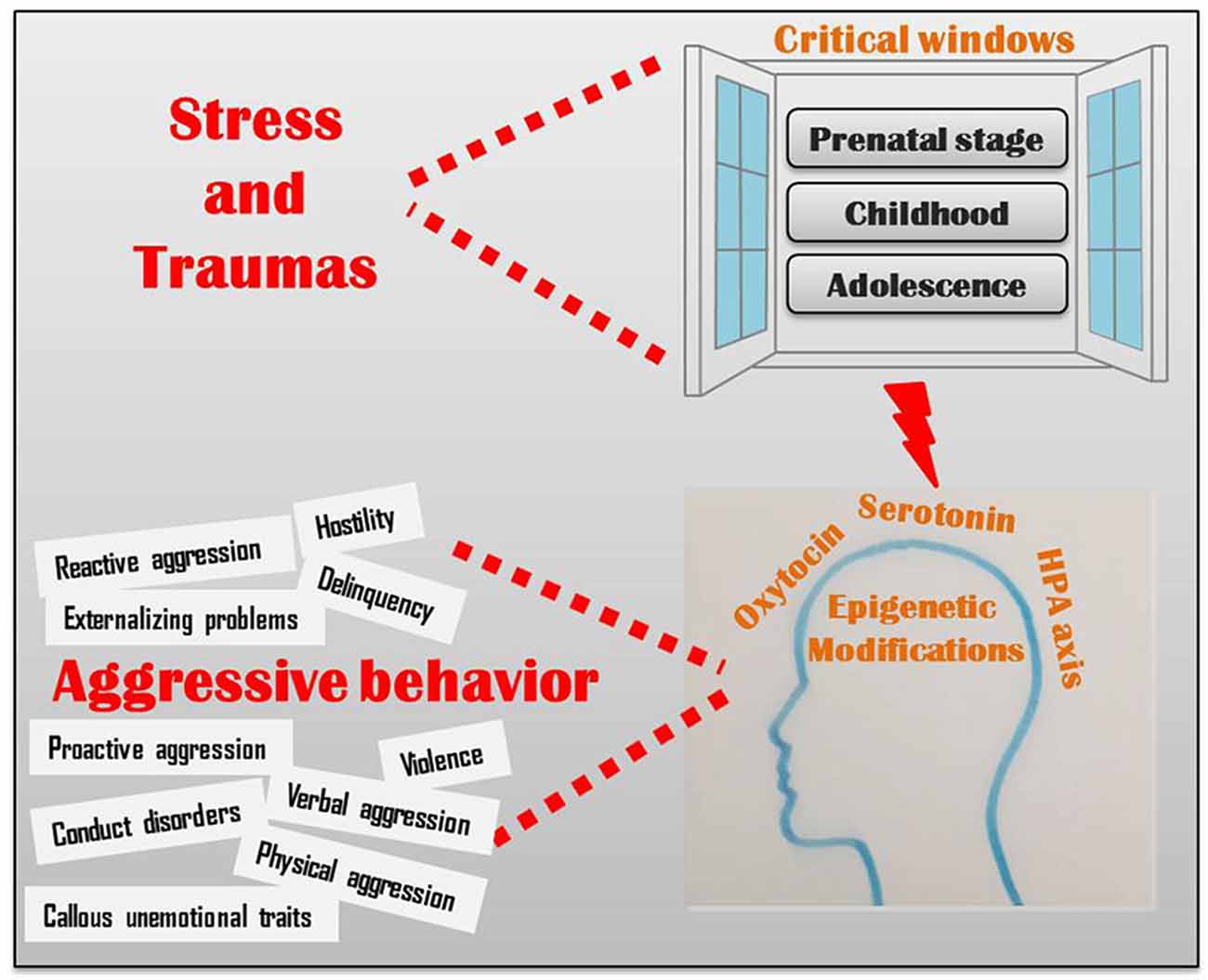
Palumbo, Mariotti, Ioffreda y Pellegrini (2018) concluyen que la epigenética arroja una nueva luz sobre la fina interacción entre la naturaleza y la crianza, al proporcionar una novedosa herramienta para comprender los eventos moleculares que subyacen a la relación entre genes, cerebro, medio ambiente y comportamiento. En conjunto, los resultados de los estudios que discutimos brevemente en el presente artículo, indican claramente que, en lo que respecta al comportamiento (humano), la naturaleza y la crianza no deben considerarse como dos factores distintos y separados, contrariamente al predominio alternante de cualquiera de los cuales se ha propuesto en diferentes fases históricas (Levitt, 2013; Moore, 2016). De hecho, distintos antecedentes genéticos modulan diferencialmente la susceptibilidad individual al medio ambiente y al mismo tiempo diversas condiciones ambientales afectan diferencialmente la expresión génica, de una manera íntima y fascinante que los científicos han comenzado a desenredar ahora. Los hallazgos de esta investigación allanan el camino hacia un enfoque novedoso para la comprensión del comportamiento humano, con importantes implicaciones también para las ciencias sociales, incluyendo la filosofía, la ética y el derecho.
Conclusiones
La ira y el miedo se consideran emociones básicas. Si bien son principalmente negativos, cumplen funciones de supervivencia muy importantes. Parecen tener claras bases genéticas y biológicas que han sido discutidas en la investigación anterior.
Referencias
- Barrett, L. F., Lindquist, K. A., Bliss-Moreau, E., Duncan, S., Gendron, M., Mize, J., et al. (2007). De ratones y hombres: ¿tipos naturales de emociones en el cerebro de los mamíferos? Una respuesta a Panksepp e Izard. Perspectiva. Psicólico. Sci. 2, 297—312. doi: 10.1126/ciencia.7652558
- Barrett, L. F., y Russell, J. A. (1999). La estructura del afecto actual. Curr. Dir. Psicólico. Sci. 8, 10.
- Barrett, L. F., y Wager, T. D. (2006). La estructura de la emoción: Evidencia a partir de estudios de neuroimagen. Curr. Dir. Psicólico. Sci. 5, 79—83.
- Blanchard, D. C., y Blanchard, R. J. (1972). Reacciones innatas y condicionadas a la amenaza en ratas con lesiones amígdaloides. J. Comp. Fisiol. Psychol. 81, 281—290.
- Bradley, M. M., Codispoti, M., Cuthbert, B. N., y Lang, P. J. (2001). Emoción y motivación I: reacciones defensivas y apetitivas en el procesamiento de imágenes. Emoción 1, 276—298. https://doi.org/10.1037/1528-3542.1.3.276
- Buckholtz, J. W. & Meyer-Lindenberg, A. (2008). MAOA y la arquitectura neurogenética de la agresión humana. Tendencias Neurosci. https://doi.org/10.1016/j.tins.2007.12.006
- Casos, O. et al. (1995). Comportamiento agresivo y cantidades alteradas de serotonina y norepinefrina cerebrales en ratones carentes de MAOA. Ciencia (80-) https://doi.org/10.1126/science.7792602
- Christie, I. C., y Friedman, B. H. (2004). Especificidad autonómica de la emoción discreta y dimensiones del espacio afectivo: un enfoque multivariado. Int. J. Psicofisiol. 51, 143—153. https://doi.org/10.1016/j.ijpsycho.2003.08.002
- Colibazzi, T., Posner, J., Wang, Z., Gorman, D., Gerber, A., Yu, S., et al. (2010). Sistemas neuronales que someten valencia y excitación durante la experiencia de las emociones inducidas. Emoción 10, 377—389. https://doi.org/10.1037/a0018484
- Davis, M. (1992). El papel de la amígdala en el miedo y la ansiedad. Annú. Rev. Neurosci. 15, 353—375. doi/10.1146/anurev.ne.15.030192.002033
- Davis, M. (1997). Neurobiología de las respuestas al miedo: el papel de la amígdala. J. Neuropsiquiatría Clin. Neurosci. 9, 382—402. https://doi.org/10.1176/jnp.9.3.382
- Ekman, P., Levenson, R. W., y Friesen, W. V. (1983). La actividad del sistema nervioso autónomo distingue entre las emociones. Ciencia 221, 1208—1210. doi: 10.1126/ciencia.6612338
- Feinstein, J. S., Buzza, C., Hurlemann, R., Follmer, R. L., Dahdaleh, N. S., Coryell, W. H., et al. (2013). Miedo y pánico en humanos con daño bilateral de amígdala. Nat. Neurosci. 16, 270—222. https://doi.org/10.1016/j.cub.2010.11.042
- Ficks, C. A. & Waldman, I. D. (2014). Genes candidatos a agresión y comportamiento antisocial: un metaanálisis de estudios de asociación de 5HTTLPR y MAOA-UVNTR. Comportarse. Genet. https://doi.org/10.1007/s10519-014-9661-y
- Godar, S. C., Fite, P. J., McFarlin, K. M. & Bortolato, M. (2016). El papel de la monoaminooxidasa A en la agresión: desarrollos traslacionales actuales y desafíos futuros. Prog. Neuropsicofarmacol. Biol. Psiquiatría https://doi.org/10.1016/j.pnpbp.2016.01.001
- Ikemoto, S., y Panksepp, J. (1999). El papel de la dopamina nuclear accumbens en el comportamiento motivado: una interpretación unificadora con especial referencia a la búsqueda de recompensas. Cerebro Res. Cerebro Res. Rev. 31, 6—41. https://doi.org/10.1016/S0165-0173(99)00023-5
- Izard, C. E. (1992). Emociones básicas, relaciones entre emociones y relaciones emociones-cognición. Psicólico. Rev. 99, 561—565. https://doi.org/10.1037/0033-295X.99.3.561
- Izard, C. E. (2007). Emociones básicas, tipos naturales, esquemas emocionales y un nuevo paradigma. Perspectiva. Psicólico. Sci. 2, 260—280.
- Johnson, P. L., Fitz, S. D., Hollis, J. H., Moratalla, R., Lightman, S. L., Shekhar, A., et al. (2011). Inducción de c-Fos en circuitos cerebrales relacionados con “pánico/defensa” tras breve exposición a gases hipercárbicos. J. Psicofarmacol. 25, 26—36. https://doi.org/10.1177/0269881109353464
- LeDoux, J. E. (2000). Circuitos de emoción en el cerebro. Annú. Rev. Neurosci. 23, 155—184. https://doi.org/10.1146/annurev.neuro.23.1.155
- LeDoux, J. (2012). Repensar el cerebro emocional. Neurona 73, 653—676. https://doi.org/10.1016/j.neuron.2012.02.004
- Lench, H. C., Flores, S. A., y Bench, S. W. (2011). Las emociones discretas predicen cambios en la cognición, el juicio, la experiencia, el comportamiento y la fisiología: un metaanálisis de las elicitaciones experimentales de emociones. Psicólico. Bol. 137, 834—855. https://doi.org/10.1037/a0024244
- Levitt, M. (2013). Percepciones de la naturaleza, crianza y comportamiento. Vida Sci. Soc. Política, 9 (13). doi: 10.1186/2195-7819-9-13
- Lindquist, K. A., Wager, T. D., Kober, H., Bliss-Moreau, E., y Barrett, L. F. (2012). La base cerebral de la emoción: una revisión metaanalítica. Comportarse. Cerebro. Sci. 35, 121—143. doi:10.1017/S0140525x11000446
- Moore, D. S. (2016). Epigenética conductual. Wiley Interdiscip. Rev. Syst. Biol. Med. 9:e1333. doi: 10.1002/wsbm.1333
- Moors, A., Ellsworth, P. C., Scherer, K. R., y Frijda, N. H. (2013). Teorías de valoración de la emoción: estado del arte y desarrollo futuro. Emot. Rev. 5, 119—124.
- Panksepp, J. (2005). Conciencia afectiva: sentimientos emocionales centrales en animales y humanos. Consciente. Cogn. 14, 30—80. https://doi.org/10.1016/j.concog.2004.10.004
- Neumann I.D., Veenema, A.H. & Beiderbeck, D.I. (2010) Agresión y ansiedad: contexto social y vínculos neurogiológicos. Frente. Comportarse. Neurosci. 4 (12). doi: 10.3389/fnbeh.2010.00012
- Panksepp, J. (2007). Neurlogizar la psicología de los afectos. Perspectiva. Psicólico. Sci. 2, 281—296.
- Peeters, D. G. A., Lange, W.-G., von Borries A. K. L., Franke, B., Volman I., Homberg J. R., Verkes R.-J., Roelofs, K. (2020). Las tendencias a evitar amenazas moderan el vínculo entre la variación genética del transportador de serotonina y la agresión reactiva. Fronteras en la Neurociencia del Comportamiento, 14, 172. doi:10.3389/fnbeh.2020.562098
- Popova, N. K. (2006). De los genes al comportamiento agresivo: el papel del sistema serotoninérgico. BioEnsayos https://doi.org/10.1002/bies.20412
- Roseman, I. (2013). Valoración en el sistema emocional: coherencia en las estrategias de afrontamiento. Emot. Rev. 5, 141—149.
- Ruisch, I. H., Dietrich, A., Glennon, J. C., Buitelaar, J. K. & Hoekstra, P. J. (2019). Interacción entre variantes genéticas implicadas en todo el genoma y factores ambientales relacionados con el comportamiento antisocial infantil en la cohorte ALSPAC del Reino Unido. Eur. Arco. Psiquiatría Clin. Neurosci. https://doi.org/10.1007/s00406-018-0964-5
- Russell, J. A. (2003). Afecto central y construcción psicológica de la emoción. Psicólico. Rev. 110, 145—172. https://doi.org/10.1037/0033-295X.110.1.145
- Russell, J. A., y Barrett, L. F. (1999). Afecto central, episodios emocionales prototípicos, y otras cosas llamadas emoción: disecar al elefante. J. Pers. Soc. Psychol. 76, 805—819. https://doi.org/10.1037/0022-3514.76.5.805
- Shih, J. C. & Chen, K. (1999). Los ratones knock-out de los genes MAO-A y -B muestran un comportamiento claramente diferente. Neurobiología, 7, 235—246 (1999).
- Sohrabi, S. (2015). El gen criminal: el vínculo entre MAOA y agresión (REVISIÓN). BMC Proc. https://doi.org/10.1186/1753-6561-9-s1-a49
- Takahashi, A., Quadros, I. M., de Almeida, R. M. M. & Miczek, K. A. (2012). Comportamiento y farmacogenética de la conducta agresiva. Curr. Parte superior. Comportarse. Neurosci. https://doi.org/10.1007/7854_2011_191 Sohrabi, S. (2015). El gen criminal: el vínculo entre MAOA y agresión (REVISIÓN). BMC Proc. https://doi.org/10.1186/1753-6561-9-s1-a49
- Takahashi, A., Quadros, I. M., de Almeida, R. M. M. & Miczek, K. A. (2012). Comportamiento y farmacogenética de la conducta agresiva. Curr. Parte superior. Comportarse. Neurosci. https://doi.org/10.1007/7854_2011_191
- Tracy, J. L., y Randles, D. (2011). Cuatro modelos de emociones básicas: una revisión de Ekman y Cordaro, Izard, Levenson, y Panksepp y Watt. Emot. Rev. 3, 397—405.
- van Honk, J., y Schutter, D. J. L. G. (2006). De la valencia afectiva a la dirección motivacional La asimetría frontal de la emoción revisada. Psicólico. Sci. 17, 963—965. https://doi.org/10.1111/j.1467-9280.2006.01813.x
- Vassos, E., Collier, D. A. & Fazel, S. (2014). Metaanálisis sistemáticos y sinopsis de campo de estudios de asociación genética de violencia y agresión. Mol. Psiquiatría https://doi.org/10.1038/mp.2013.31
- Vytal, K., y Hamann, S. (2010). Soporte de neuroimagen para correlatos neuronales discretos de emociones básicas: un metanálisis basado en voxel. J. Cogn. Neurosci. 22, 2864—2885. https://doi.org/10.1162/jocn.2009.21366
- Wager, T. D., Phan, K. L., Liberzon, I., y Taylor, S. F. (2003). Valencia, género y lateralización de la anatomía funcional cerebral en la emoción: un metaanálisis de hallazgos de neuroimagen. Neuroimagen 19, 513—531. https://doi.org/10.1016/S1053-8119(03)00078-8
- Waltes, R., Chiocchetti, A. G. & Freitag, C. M. (2016). Las bases neurobiológicas de la agresión humana: una revisión sobre los mecanismos genéticos y epigenéticos. Am. J. Med. Genet. Parte B Neuropsiquiatr.Genet. https://doi.org/10.1002/ajmg.b.32388
- Wilensky, A. E., Schafe, G. E., Kristensen, M. P., y LeDoux, J. E. (2006). Repensar el circuito del miedo: el núcleo central de la amígdala es requerido para la adquisición, consolidación y expresión del condicionamiento pavloviano del miedo. J. Neurosci. 26, 12387—12396. https://doi.org/10.1523/JNEUROSCI.4316-06.2006
- Youdim, M. B. H., Edmondson, D. & Tipton, K. F. (2006). El potencial terapéutico de los inhibidores de la monoaminooxidasa. Nat. Rev. Neurosci. https://doi.org/10.1038/nrn1883
Atribuciones
Qué podemos aprender sobre las emociones estudiando la psicopatía por Abigail Marsh, en Fronteras en Neurociencia Humana licenciada CC BY 3.0
El modelo de excitación y valencia del psicólogo Russell por http://imagine-it.org/gamessurvey/, con licencia CC BY 3.0, vía Wikimedia Commons
Emociones individuales por U3161650, CC BY-SA 4.0, vía Wikimedia Commons
Imagen de Base de Datos Centro de Ciencias de la Vida (DBCLS) con licencia CC-BY 4.0 vía Wikimedia commons.
De los genes guerreros a las soluciones traduccionales: nuevos conocimientos sobre las monoaminooxidasas (MAO) y la agresión. por Alexios-Fotios A. Mentis, Efthemios Dardiotis, Eleni Katsouni, George P. Chrousos. en Psiquiatría Transl licenciada CC-BY 4.0
Genes y Comportamiento Agresivo: Mecanismos Epigenéticos Subyacentes a la Susceptibilidad Individual a Ambientes Aversivos por Sara Palumbo, Veronica Mariotti, Caterina Ioffreda y Silvia Pellegrini en Fronteras en Neurociencia del Comportamiento (2018) licenciado CC BY 4.0