
8.2: Visión
- Page ID
- 144005

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Universidad de Illinois en Urbana-Champaign
La visión es la modalidad sensorial que transforma la luz en una experiencia psicológica del mundo que te rodea, con un mínimo esfuerzo corporal. Este módulo proporciona una visión general de los pasos más significativos en esta transformación y las estrategias que su cerebro utiliza para lograr esta comprensión visual del entorno.
objetivos de aprendizaje
- Describir cómo el ojo transforma la información de la luz en energía neuronal.
- Entender qué tipo de información le interesa al cerebro extraer del entorno y por qué es útil.
- Describir cómo se ha adaptado el sistema visual para hacer frente a diferentes condiciones de iluminación.
- Entender el valor de tener dos ojos.
- Entender por qué tenemos visión del color.
- Comprender la interdependencia entre la visión y otras funciones cerebrales.
¿Qué es la visión?
Piensa en el espectáculo de una noche estrellada. Miras hacia el cielo y miles de fotones de estrellas distantes se estrellan contra tu retina, una estructura sensible a la luz en la parte posterior de tu globo ocular. Estos fotones tienen millones de años y han sobrevivido a un viaje a través del universo, solo para encontrarse con uno de tus fotorreceptores. Mala suerte: en una milésima de segundo, este poquito de energía lumínica se convierte en el combustible de una reacción fotoquímica conocida como fotoactivación. La energía lumínica se convierte en energía neuronal y desencadena una cascada de actividad neuronal que, unas centésimas de segundo después, dará como resultado que tomes conciencia de esa estrella distante. Tú y el universo unidos por fotones. Ese es el asombroso poder de la visión. La luz te trae el mundo. Sin moverte, ya sabes lo que hay ahí fuera. Puedes reconocer a amigos que vienen a conocerte antes de que puedas escucharlos venir, frutos maduros de verdes en árboles sin tener que probarlos y antes de llegar a agarrarlos. También puedes decir qué tan rápido se mueve una pelota en tu dirección (¿Te golpeará? ¿Se le puede pegar?).
¿Cómo sucede todo eso? Primero, la luz ingresa al globo ocular a través de un pequeño agujero conocido como la pupila y, gracias a las propiedades refractivas de tu córnea y lente, esta señal de luz se proyecta bruscamente hacia la retina (consulta Recursos externos para ver enlaces a una descripción más detallada de la estructura del ojo). Allí, la luz es transducida en energía neural por cerca de 200 millones de células fotorreceptoras.

Aquí es donde la información que lleva la luz sobre objetos y colores distantes comienza a ser codificada por nuestro cerebro. Existen dos tipos diferentes de fotorreceptores: bastones y conos. El ojo humano contiene más bastones que conos. Las varillas nos dan sensibilidad bajo condiciones de iluminación tenue y nos permiten ver por la noche. Los conos nos permiten ver detalles finos en luz brillante y darnos la sensación de color. Los conos están apretados alrededor de la fóvea (la región central de la retina detrás de la pupila) y más escasamente en otros lugares. Las varillas pueblan la periferia (la región que rodea la fóvea) y están casi ausentes de la fóvea.
Pero la visión es mucho más compleja que solo atrapar fotones. La información codificada por los fotorreceptores se somete a un conjunto rápido y continuo de análisis cada vez más complejos para que, eventualmente, se pueda dar sentido a lo que hay ahí fuera. En la fóvea, la información visual se codifica por separado de pequeñas porciones del mundo (cada una aproximadamente la mitad del ancho de un cabello humano visto a la altura del brazo) para que eventualmente el cerebro pueda reconstruir con gran detalle las diferencias visuales finas de las ubicaciones en las que estás mirando directamente. Este fino nivel de codificación requiere mucha luz y va lento (neuralmente hablando). En contraste, en la periferia, hay una estrategia de codificación diferente: el detalle se sacrifica a cambio de sensibilidad. La información se resume en secciones más grandes del mundo. Esta agregación se produce rápidamente y permite detectar señales tenues bajo niveles de luz muy bajos, así como detectar movimientos bruscos en su visión periférica.
La importancia del contraste
¿Qué pasa después? Bueno, podrías pensar que el ojo haría algo como registrar la cantidad de luz en cada lugar del mundo y luego enviar esta información a las áreas de procesamiento visual del cerebro (¡un sorprendente 30% de la corteza está influenciada por señales visuales!). Pero, de hecho, eso no es lo que hacen los ojos. Tan pronto como los fotorreceptores capturan la luz, el sistema nervioso se ocupa analizando las diferencias en la luz, y son estas diferencias las que se transmiten al cerebro. Al cerebro, resulta, le importa poco la cantidad total de luz proveniente de una parte específica del mundo, o en la escena en general. Más bien, quiere saber: ¿la luz que viene de este punto difiere de la luz que viene del punto siguiente? Coloca tu mano sobre la mesa frente a ti. El contorno de tu mano en realidad está determinado por la diferencia de luz —el contraste — entre la luz que viene de la piel de tu mano y la luz que viene de la mesa debajo. Para encontrar el contorno de tu mano, simplemente necesitamos encontrar las regiones en la imagen donde la diferencia de luz entre dos puntos adyacentes sea máxima. Dos puntos en tu piel reflejarán niveles similares de luz de vuelta a ti, al igual que dos puntos sobre la mesa. Por otro lado, dos puntos que caen a cada lado del contorno limítrofe entre tu mano y la mesa reflejarán una luz muy diferente.
El hecho de que el cerebro esté interesado en codificar el contraste en el mundo revela algo profundamente importante sobre las fuerzas que impulsaron la evolución de nuestro cerebro: codificar la cantidad absoluta de luz en el mundo nos dice poco sobre lo que hay ahí fuera. Pero si tu cerebro puede detectar la repentina aparición de una diferencia de luz en algún lugar frente a ti, entonces debe ser que algo nuevo esté ahí. Esa señal de contraste es información. Esa información puede representar algo que te guste (comida, un amigo) o algo peligroso que se acerca (un tigre, un acantilado). El resto de tu sistema visual trabajará duro para determinar qué es esa cosa, pero tan rápido como 10 ms después de que la luz entre en tus ojos, las células ganglionares de tus retinas ya han codificado todas las diferencias de luz del mundo frente a ti.
El contraste es tan importante que tus neuronas salen de su camino no sólo para codificar las diferencias en la luz sino para exagerar esas diferencias para ti, para que no las pierdas. Las neuronas logran esto a través de un proceso conocido como inhibición lateral. Cuando una neurona está disparando en respuesta a la luz, produce dos señales: una señal de salida para pasar al siguiente nivel en la visión, y una señal lateral para inhibir todas las neuronas que están al lado de ella. Esto tiene sentido en el supuesto de que las neuronas cercanas probablemente estén respondiendo a la misma luz proveniente de ubicaciones cercanas, por lo que esta información es algo redundante. La magnitud de la señal inhibitoria lateral que produce una neurona es proporcional a la entrada excitadora que recibe la neurona: cuanto más dispara una neurona, más fuerte es la inhibición que produce. La Figura 8.2.1 ilustra cómo la inhibición lateral amplifica las señales de contraste en los bordes de las superficies.
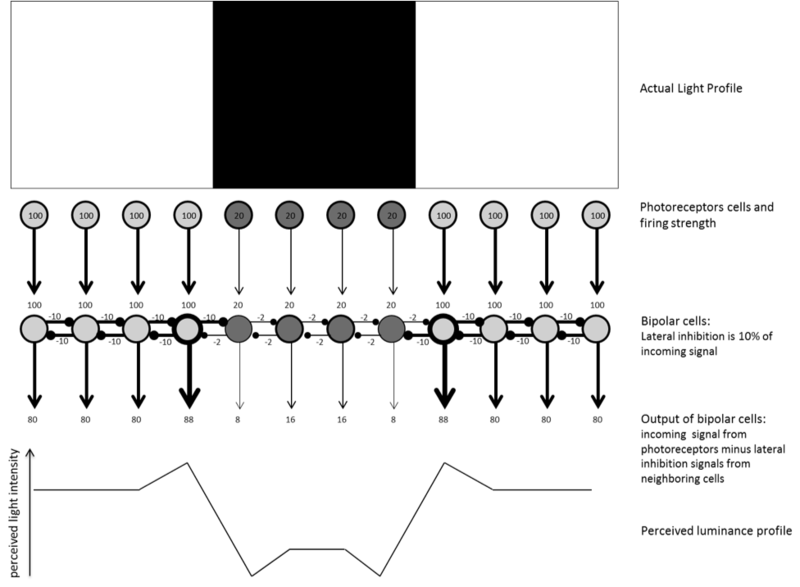
Sensibilidad a diferentes condiciones de luz

Pensemos por un momento en la gama de condiciones en las que su sistema visual debe operar día tras día. Cuando das un paseo al aire libre en un día soleado, hasta miles de millones de fotones entran en tus globos oculares cada segundo. Por el contrario, cuando te despiertas en mitad de la noche en una habitación oscura, podría haber tan poco como unos cientos de fotones por segundo entrando en tus ojos. Para hacer frente a estos extremos, el sistema visual se basa en las diferentes propiedades de los dos tipos de fotorreceptores. Las varillas son las principales responsables de procesar la luz cuando los fotones son escasos (¡solo un solo fotón puede hacer que una barra se dispare!) , pero lleva tiempo reponer el pigmento visual que las varillas requieren para la fotoactivación. Entonces, en condiciones brillantes, las varillas se blanquean rápidamente (Stuart & Brige, 1996) y no pueden mantenerse al día con el constante aluvión de fotones que las golpean. Ahí es cuando los conos se vuelven útiles. Los conos requieren más fotones para disparar y, más críticamente, sus fotopigmentos se reponen mucho más rápido que los fotopigmentos de las varillas, lo que les permite mantenerse al día cuando los fotones son abundantes.
¿Qué sucede cuando cambias abruptamente las condiciones de iluminación? Bajo luz brillante, tus varillas están blanqueadas. Cuando te mueves a un ambiente oscuro, tomará tiempo (hasta 30 minutos) antes de que se recuperen químicamente (Hurley, 2002). Una vez que lo hagan, comenzarás a ver cosas a tu alrededor que inicialmente no pudiste. Este fenómeno se llama adaptación a la oscuridad. Cuando pasas de la oscuridad a la luz brillante (al salir de un túnel en una carretera, por ejemplo), tus cañas quedarán blanqueadas en un incendio y quedarás cegado por la luz repentina durante aproximadamente 1 segundo. Sin embargo, ¡tus conos están listos para disparar! Su disparo se hará cargo y rápidamente comenzarás a ver en este nivel más alto de luz.
Un ajuste similar, pero más sutil, ocurre cuando el cambio en la iluminación no es tan drástico. Piense en su experiencia de leer un libro por la noche en su cama en comparación con leer al aire libre: la habitación puede sentirse bastante bien iluminada (suficiente para que pueda leer) pero las bombillas de su habitación no están produciendo los miles de millones de fotones que encuentra afuera. En ambos casos, sientes que tu experiencia es la de un ambiente bien iluminado. No sientes una experiencia como millones de veces más brillante que la otra. Esto se debe a que la visión (tanto de la percepción) no es proporcional: ver el doble de fotones no produce la sensación de ver el doble de brillante que una luz. El sistema visual sintoniza la experiencia actual favoreciendo una gama de valores de contraste más informativos en ese entorno (Gardner et al., 2005). Este es el concepto de ganancia de contraste: el sistema visual determina el contraste medio en una escena y representa mejor los valores alrededor de ese contraste medio, mientras ignora las diferencias de contraste más pequeñas. (Consulte la sección de Recursos Externos para una demostración.)
El proceso de reconstrucción
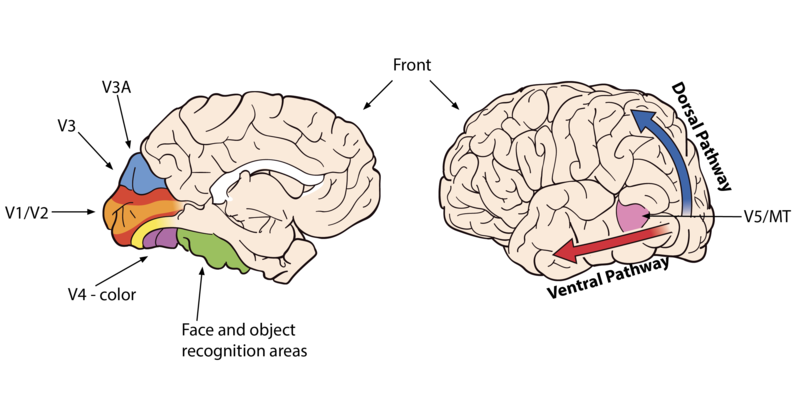
¿Qué sucede una vez que la información sale de tus ojos y entra en el cerebro? Las neuronas se proyectan primero en el tálamo, en una sección conocida como núcleo geniculado lateral. La información luego se divide y se proyecta hacia dos partes diferentes del cerebro. La mayoría de los cálculos con respecto a los movimientos oculares reflexivos se calculan en regiones subcorticales, la parte evolutivamente más antigua del cerebro. Los movimientos oculares reflexivos le permiten orientar rápidamente sus ojos hacia las áreas de interés y rastrear los objetos a medida que se mueven. Los cálculos más complejos, aquellos que eventualmente te permiten tener una experiencia visual del mundo, todos ocurren en la corteza, la región evolutivamente más nueva del cerebro. La primera parada en la corteza es en la corteza visual primaria (también conocida como V1). Aquí, el proceso de “reconstrucción” comienza con seriedad: a partir de la información de contraste que llega de los ojos, las neuronas comenzarán a computar información sobre el color y las líneas simples, detectando diversas orientaciones y espesores. También se calculan señales de movimiento a pequeña escala (Hubel & Wiesel, 1962).
A medida que la información comienza a fluir hacia otras áreas “superiores” del sistema, se realizan cálculos más complejos. Por ejemplo, los bordes se asignan al objeto al que pertenecen, los fondos se separan de los primeros planos, los colores se asignan a las superficies y se calcula el movimiento global de los objetos. Muchos de estos cálculos ocurren en áreas cerebrales especializadas. Por ejemplo, un área llamada MT procesa información de movimiento global; el área de lugar parahipocampo identifica ubicaciones y escenas; el área de la cara fusiforme se especializa en identificar objetos para los que se requieren discriminaciones finas, como rostros. Incluso hay una región cerebral especializada en el procesamiento de letras y textos. Estas áreas de reconocimiento visual se localizan a lo largo de la vía ventral del cerebro (también conocida como la vía Qué). Otras regiones del cerebro a lo largo de la vía dorsal (o vía de dónde y cómo) computarán información sobre el movimiento de uno mismo y de los objetos, lo que le permitirá interactuar con los objetos, navegar por el entorno y evitar obstáculos (Goodale y Milner, 1992).
Ahora que tienes una comprensión básica de cómo funciona tu sistema visual, puedes hacerte la pregunta: ¿por qué tienes dos ojos? Todo lo que discutimos hasta ahora podría calcularse con información proveniente de un solo ojo. Entonces, ¿por qué dos? Mirar el reino animal nos da una pista. Los animales que tienden a ser presas tienen ojos ubicados en lados opuestos de su cráneo. Esto les permite detectar depredadores cada vez que uno aparece en cualquier lugar a su alrededor. Los humanos, como la mayoría de los depredadores, tienen dos ojos apuntando en la misma dirección, codificando casi la escena exacta dos veces. Esta redundancia nos da una ventaja binocular: tener dos ojos no sólo te brinda dos posibilidades de captar una señal frente a ti, sino que la minúscula diferencia de perspectiva que obtienes de cada ojo es utilizada por tu cerebro para reconstruir el sentido del espacio tridimensional. Puedes obtener una estimación de lo lejos que están los objetos de ti, su tamaño y su volumen. Esto no es una hazaña fácil: la señal en cada ojo es una proyección bidimensional del mundo, como dos imágenes separadas dibujadas sobre tu retina. Sin embargo, tu cerebro te proporciona sin esfuerzo una sensación de profundidad al combinar esas dos señales. Este proceso de reconstrucción 3-D también se basa en gran medida en todos los conocimientos adquiridos a través de la experiencia sobre información espacial. Por ejemplo, su sistema visual aprende a interpretar cómo cambian el volumen, la distancia y el tamaño de los objetos a medida que se acercan o se alejan de usted. (Consulte la sección de Recursos Externos para ver demostraciones.)
La experiencia del color
Quizás uno de los aspectos más bellos de la visión es la riqueza de la experiencia de color que nos brinda. Uno de los retos que tenemos como científicos es entender por qué la experiencia del color humano es lo que es. Quizás hayas escuchado que los perros solo tienen 2 tipos de fotorreceptores de color, mientras que los humanos tienen 3, los pollos tienen 4, y los camarones mantis tienen 16. ¿Por qué existe tal variación entre especies? Los científicos creen que cada especie ha evolucionado con diferentes necesidades y utiliza la percepción del color para señalar información sobre los alimentos, la reproducción y la salud que son únicos para sus especies. Por ejemplo, los humanos tienen una sensibilidad específica que permite detectar ligeros cambios en el tono de la piel. Se puede decir cuando alguien está avergonzado, excitado o enfermo. Detectar estas señales sutiles es adaptativo en una especie social como la nuestra.
¿Cómo se codifica por colores en el cerebro? Las dos teorías principales de la percepción del color se propusieron a mediados del siglo XIX, aproximadamente 100 años antes de que se encontrara evidencia fisiológica que las corroborara a ambas (Svaetichin, 1956). La teoría de la tricromacia, propuesta por Young (1802) y Helmholtz (1867), propuso que el ojo tuviera tres tipos diferentes de células sensibles al color a partir de la observación de que cualquier color se puede reproducir combinando luces de tres lámparas de diferente tonalidad. Si puedes ajustar por separado la intensidad de cada luz, en algún momento encontrarás la combinación correcta de las tres luces para que coincida con cualquier color del mundo. Este principio se usa hoy en día en televisores, pantallas de computadoras y cualquier pantalla a color. Si miras suficientemente de cerca un píxel, encontrarás que está compuesto por una luz azul, roja y verde, de intensidades variables. En cuanto a la retina, los humanos tienen tres tipos de conos: conos S, conos M y conos L (también conocidos como conos azules, verdes y rojos, respectivamente) que son sensibles a tres longitudes de onda diferentes de luz.
Casi al mismo tiempo, Hering hizo un descubrimiento desconcertante: algunos colores son imposibles de crear. Mientras que puedes hacer verdes amarillentos, rojos azulados, azules verdosos y amarillos rojizos combinando dos colores, nunca se puede hacer un verde rojizo o un amarillo azulado. Esta observación llevó a Hering (1892) a proponer la teoría del color del Proceso Oponente: el color se codifica a través de tres canales del oponente (rojo-verde, azul-amarillo y negro-blanco). Dentro de cada canal, se calcula constantemente una comparación entre los dos elementos del par. En otras palabras, los colores se codifican como diferencias entre dos tonalidades y no como simples combinaciones de tonalidades. Nuevamente, lo que le importa al cerebro es el contraste. Cuando un elemento es más fuerte que el otro, se percibe el color más fuerte y se suprime el más débil. Puedes experimentar este fenómeno siguiendo el siguiente enlace.
nobaproject.com/activos/modules/module-visio...
Cuando ambos colores en un par están presentes en igual extensión, la percepción del color se cancela y percibimos un nivel de gris. Por eso no se puede ver un verde rojizo o un amarillo azulado: se cancelan entre sí. Por cierto, si te estás preguntando de dónde viene la señal amarilla, resulta que se calcula promediando las señales de cono M y L. ¿Estos colores son colores exclusivamente humanos? Algunos piensan que son: el contraste rojo-verde, por ejemplo, está finamente afinado para detectar cambios en el tono de la piel humana para que puedas saber cuándo alguien se ruboriza o se pone pálido. Entonces, la próxima vez que salgas a pasear con tu perro, mira el atardecer y pregúntate, ¿qué color ve mi perro? ¡Probablemente ninguno de los tonos anaranjados que haces!
Entonces ahora, puedes hacerte la pregunta: ¿todos los humanos experimentan el color de la misma manera? Las personas daltónicas, como puedes imaginar, no ven todos los colores que vemos el resto de nosotros, y esto se debe a que carecen de uno (o más) conos en su retina. Por cierto, hay algunas mujeres que en realidad tienen cuatro juegos diferentes de conos en sus ojos, y investigaciones recientes sugieren que su experiencia de color puede ser (pero no siempre es) más rica que la de las personas de tres conos. Sin embargo, una pregunta ligeramente diferente es si todas las personas de tres conos tienen las mismas experiencias internas de colores: ¿el rojo dentro de tu cabeza es el mismo rojo dentro de la cabeza de tu mamá? Esa es una pregunta casi imposible de responder que ha sido debatida por los filósofos durante milenios, sin embargo, datos recientes sugieren que de hecho podría haber diferencias culturales en la forma en que percibimos el color. Resulta que no todas las culturas categorizan los colores de la misma manera, por ejemplo. Y algunos grupos “ven” diferentes tonalidades de lo que nosotros en el mundo occidental llamaríamos el “mismo” color, como colores categóricamente diferentes. La tribu Berinmo en Nueva Guinea, por ejemplo, parece experimentar tonos verdes que denotan hojas vivas como pertenecientes a una categoría de color completamente diferente al tipo de tonos verdes que denotan hojas moribundas. También los rusos parecen experimentar tonos claros y oscuros de azul como diferentes categorías de colores, de una manera que la mayoría de los occidentales no lo hacen. Además, la investigación actual de imágenes cerebrales sugiere que los cerebros de las personas cambian (aumento en el volumen de materia blanca) cuando aprenden nuevas categorías de color. Estos son hallazgos intrigantes y sugerentes, con certeza, que parecen indicar que nuestro entorno cultural puede, de hecho, tener algún impacto (pequeño) pero definitivo en la forma en que las personas usan y experimentan los colores en todo el mundo.
Integración con otras modalidades
La visión no es un sistema encapsulado. Interactúa con otras modalidades sensoriales y depende de ellas. Por ejemplo, cuando mueves la cabeza en una dirección, tus ojos se mueven reflexivamente en la dirección opuesta para compensar, permitiéndote mantener tu mirada en el objeto que estás mirando. Este reflejo se llama reflejo vestibulo-ocular. Se logra integrando información tanto del sistema visual como del vestibular (que conoce el movimiento y la posición del cuerpo). Puedes experimentar esta compensación de manera bastante simple. Primero, mientras mantienes la cabeza quieta y tu mirada mirando hacia adelante, mueve el dedo frente a ti de lado a lado. Observe cómo la imagen del dedo aparece borrosa. Ahora, mantén tu dedo firme y míralo mientras mueves la cabeza de lado a lado. Observa cómo tus ojos se mueven reflexivamente para compensar el movimiento de tu cabeza y cómo la imagen del dedo se mantiene nítida y estable. La visión también interactúa con tu sistema propioceptivo, para ayudarte a encontrar dónde están todas las partes de tu cuerpo, y con tu sistema auditivo, para ayudarte a entender los sonidos que las personas hacen cuando hablan. Puedes conocer más sobre esto en el módulo Noba sobre percepción multimodal (http://noba.to/cezw4qyn).
Finalmente, la visión también suele estar implicada en un fenómeno de mezcla de sensaciones conocido como sinestesia. La sinestesia ocurre cuando una señal sensorial da lugar a dos o más sensaciones. El tipo más común es la sinestesia grafema-color. Aproximadamente 1 de cada 200 individuos experimenta una sensación de color asociada con letras, números o palabras específicas: el número 1 siempre puede verse como rojo, el número 2 como naranja, etc. Pero las formas más fascinantes de la sinestesia mezclan sensaciones de modalidades sensoriales completamente diferentes, como el sabor y el color o música y color: el sabor del pollo podría provocar una sensación de verde, por ejemplo, y el timbre del violín un púrpura profundo.
Observaciones Finales
Estamos en un momento emocionante en nuestra comprensión científica de la visión. Acabamos de comenzar a obtener una comprensión funcional del sistema visual. No está lo suficientemente evolucionado para que recreemos sistemas visuales artificiales (es decir, todavía no podemos hacer robots que “vean” y entiendan las señales de luz como nosotros), pero estamos llegando ahí. Hace poco, grandes avances en la ciencia de la visión han permitido a los investigadores mejorar significativamente las prótesis retinianas: circuitos fotosensibles que se pueden implantar en la parte posterior del globo ocular de personas ciegas que se conectan a áreas visuales del cerebro y tienen la capacidad de restaurar parcialmente un “visual experiencia” a estos pacientes (Nirenberg & Pandarinath, 2012). Y usando imágenes cerebrales magnéticas funcionales, ahora podemos “decodificar” de tu actividad cerebral las imágenes que viste en tus sueños mientras dormías (¡Horikawa, Tamaki, Miyawaki, & Kamitani, 2013)! Sin embargo, todavía queda mucho más por entender. Considera esto: si la visión es un proceso de construcción que lleva tiempo, lo que veamos ahora ya no es lo que tenemos delante. Sin embargo, los humanos pueden hacer increíbles hazañas sensibles al tiempo como golpear una bola rápida de 90 mph en un juego de béisbol. Parece entonces que una función fundamental de la visión no es solo saber lo que está sucediendo a tu alrededor ahora, sino en realidad hacer una inferencia precisa sobre lo que estás a punto de ver a continuación (Enns & Lleras, 2008), para que puedas mantenerte al día con el mundo. Comprender cómo se logra esta función predictiva de la visión orientada al futuro en el cerebro es probablemente el próximo gran desafío en este fascinante ámbito de la investigación.
Recursos Externos
- Video: Conocimientos adquiridos y su impacto en nuestra interpretación tridimensional del mundo - 3D Street Art
- Video: Conocimientos adquiridos y su impacto en nuestra interpretación tridimensional del mundo - Ilusiones anamórficas
- Video: Conocimientos adquiridos y su impacto en nuestra interpretación tridimensional del mundo - Ilusión óptica
- Web: Increíble biblioteca con fenómenos visuales e ilusiones ópticas, explicó
- http://michaelbach.de/ot/index.html
- Web: Anatomía del ojo
- http://www.eyecareamerica.org/eyecare/anatomy/
- Web: Demostración de adaptación de ganancia de contraste
- http://www.michaelbach.de/ot/lum_contrast-adapt/
- Web: Demostración de contornos ilusorios e inhibición lateral. Bandas Mach
- http://michaelbach.de/ot/lum-MachBands/index.html
- Web: Demostración de contraste ilusorio e inhibición lateral. La grilla de Hermann
- http://michaelbach.de/ot/lum_herGrid/
- Web: Más información sobre qué y dónde y cómo vías
- http://www.scholarpedia.org/article/...where_pathways
Preguntas de Discusión
- Al correr en la oscuridad, se recomienda que nunca mires directamente al suelo. ¿Por qué? ¿Cuál sería una mejor estrategia para evitar obstáculos?
- La mayoría de las células ganglionares en el ojo se especializan en detectar gotas en la cantidad de luz proveniente de una ubicación determinada. Es decir, aumentan su velocidad de disparo cuando detectan menos luz proveniente de una ubicación específica. ¿Por qué podría ser más importante la ausencia de luz que la presencia de luz? ¿Por qué sería evolutivamente ventajoso codificar este tipo de información?
- Hay un agujero en cada uno de tus globos oculares llamado disco óptico. Aquí es donde las venas ingresan al globo ocular y donde las neuronas (los axones de las células ganglionares) salen del globo ocular. ¿Por qué no ves dos agujeros en el mundo todo el tiempo? Cierra un ojo ahora. ¿Por qué no ves ahora un agujero en el mundo? Para “experimentar” un punto ciego, siga las instrucciones de este sitio web: http://michaelbach.de/ot/cog_blindSpot/index.html
- Imagina que te dieron la tarea de probar las habilidades de percepción del color de una especie de monos recién descubierta en el Pacífico Sur. ¿Cómo lo harías?
- Un aspecto importante de las emociones es que las percibimos en nosotros mismos de la misma manera que percibimos otras percepciones como la visión. ¿Se te ocurre un ejemplo donde el concepto de ganancia de contraste pueda utilizarse para comprender las respuestas de las personas a los eventos emocionales?
El vocabulario
- Ventaja binocular
- Se beneficia de tener dos ojos en lugar de un solo ojo.
- Conos
- Fotorreceptores que operan en ambientes iluminados y pueden codificar detalles visuales finos. Hay tres tipos diferentes (S o azul, M o verde y L o rojo) que son cada uno sensibles a tipos de luz ligeramente diferentes. Combinados, estos tres tipos de conos permiten tener visión de color.
- Contraste
- Diferencia relativa en la cantidad y tipo de luz proveniente de dos ubicaciones cercanas.
- Ganancia de contraste
- Proceso donde se puede ajustar la sensibilidad de su sistema visual para que sea más sensible a los niveles de contraste que prevalecen en el entorno.
- Adaptación a la oscuridad
- Proceso que te permite llegar a ser sensible a niveles muy pequeños de luz, para que realmente puedas ver en la casi ausencia de luz.
- Inhibición lateral
- Una señal producida por una neurona destinada a suprimir la respuesta de las neuronas cercanas.
- Teoría del proceso del oponente
- Teoría de la visión del color que asume que hay cuatro colores básicos diferentes, organizados en dos pares (rojo/verde y azul/amarillo) y propone que los colores en el mundo se codifican en términos de la oponencia (o diferencia) entre los colores de cada par. Hay un par negro/blanco adicional responsable de codificar el contraste de luz.
- Fotoactivación
- Una reacción fotoquímica que se produce cuando la luz incide en los fotorreceptores, produciendo una señal neural.
- Corteza visual primaria (V1)
- Región cerebral ubicada en la corteza occipital (hacia la parte posterior de la cabeza) responsable del procesamiento de información visual básica como la detección, grosor y orientación de líneas simples, color y movimiento a pequeña escala.
- Varillas
- Fotorreceptores que son muy sensibles a la luz y son en su mayoría responsables de la visión nocturna.
- Sinestesia
- La mezcla de dos o más experiencias sensoriales, o la activación automática de una experiencia sensorial secundaria (indirecta) debido a ciertos aspectos de la estimulación sensorial primaria (directa).
- Teoría de la tricromacia
- Teoría que propone que toda tu percepción del color se basa fundamentalmente en la combinación de tres (no dos, no cuatro) señales de color diferentes.
- Reflejo vestibulo-ocular
- Coordinación de información de movimiento con información visual que te permita mantener tu mirada sobre un objeto mientras te mueves.
- Qué camino
- Vía de procesamiento neuronal en el cerebro que es responsable de tu capacidad de reconocer lo que te rodea.
- Vía de dónde y cómo
- Vía de procesamiento neuronal en el cerebro que se encarga de que sepas dónde están las cosas en el mundo y cómo interactuar con ellas.
Referencias
- Enns, J. T., & Lleras, A. (2008). Nuevas evidencias para la predicción en la visión humana. Tendencias en Ciencias Cognitivas, 12 ¸ 327—333.
- Gardner, J. L., Sun, P., Waggoner, R. A., Ueno, K., Tanaka, K., & Cheng, K. (2005). Adaptación y representación del contraste en la corteza visual temprana humana. Neurona, 47, 607—620.
- Goodale, M. A., & Milner, A. D. (1992). Separar las vías visuales para la percepción y la acción. Tendencias en Neurociencia, 15, 20—25.
- Helmholtz, H. von. (1867). Handbuch der Physiologischen Optik. Leipzig: Leopold Voss.
- Hering, E. (1892). Grundzüge der Lehre vom Lichtsinn. Berlín, Alemania: Springer.
- Horikawa, T., Tamaki, M., Miyawaki, Y., & Kamitani, Y. (2013). Decodificación neuronal de imágenes visuales durante el sueño. Ciencia, 340 (6132), 639—642.
- Hubel, D. H., & Wiesel, T. N. (1962). Campos receptivos, interacción binocular y arquitectura funcional en la corteza visual del gato. Revista de Fisiología, 160, 106—154.
- Hurley, J. B. (2002). Derramando luz sobre la adaptación. Revista de Fisiología General, 119, 125—128.
- Nirenberg, S., & Pandarinath, C. (2012). Estrategia protésica retiniana con capacidad para restaurar la visión normal. Actas de la Academia Nacional de Ciencias, 109 (37), 15012—15017.
- Stuart, J. A., & Brige, R. R. (1996). Caracterización de los eventos fotoquímicos primarios en bacteriorodopsina y rodopsina. En A. G. Lee (Ed.), Rhodopsina y receptores unidos a proteína G (Parte A, Vol. 2, pp. 33—140). Greenwich, CT: JAI.
- Svaetichin, G. (1956). Curvas de respuesta espectral de conos individuales, Actaphysiologica Scandinavia, Suppl. 134, 17—46.
- Young, T. (1802). Conferencia Bakeriana: Sobre la teoría de la luz y los colores. Transacción filosófica de la Royal Society Londres, 92, 12—48.