
4.14: Dibujos para Compuestos Biológicos Grandes
- Page ID
- 80561
Las estructuras lineales serán el tipo de dibujo estructural más común que veas en estas páginas. También verá algunas otras utilizadas para fines específicos, y esas representaciones se explican aquí.
A veces incluso dibujar una estructura de líneas es demasiado. Las proteínas a menudo contienen miles de átomos. Presentar toda esa información sobre la unión podría dificultar la lectura del dibujo, especialmente en una forma tridimensional.
Una imagen de una molécula se puede presentar de varias maneras. Un modelo de bola y palo da una idea bastante buena de dónde están todos los átomos y cómo están conectados. Las bolas representan los átomos y los palos representan los enlaces. A continuación se muestra un ejemplo de una imagen de bola y barra de una proteína, proteína tirosil fosfato bovina de bajo peso molecular, que se une a fosfato inorgánico.

Sin embargo, hay un par de problemas para ver una proteína de esta manera. El primer problema es que normalmente no sabemos dónde están los átomos de hidrógeno, por lo que faltan en la estructura de bola y palo. ¿Por qué es eso?
Se debe a que la estructura de la proteína generalmente se determina experimentalmente mediante cristalografía de rayos X. Eso significa que alguien se ha tomado la molestia de aislar una muestra pura de proteína de un organismo. Disuelvieron la proteína con mucho cuidado, luego la encerraron para que formara cristales. Hay diferentes formas de hacerlo, pero generalmente implica enfriar la solución, de manera que el solvente ya no tenga suficiente energía para mantener todas las moléculas disueltas, o bien evaporar un poco del solvente, de manera que ya no haya suficiente solvente para mantener todo disuelto. Este proceso puede llevar semanas. En ese punto, podían levantar cuidadosamente un solo cristal de la solución y montarse en un difractómetro de rayos X. Cuando los rayos X brillan a través del cristal, esencialmente rebotan en los átomos y se dispersan en diferentes direcciones. Al medir dónde aterrizan los rayos X, y usar algún software de computadora que explote esos números imaginarios de los que aprendiste en la secundaria pero que no pensaste que fueran útiles para nada, el investigador puede determinar la ubicación de cada átomo en la molécula (si tienen suerte). Sin embargo, los átomos de hidrógeno son demasiado pequeños para localizarlos con certeza.
- La difractometría de rayos X puede revelar la ubicación de cada átomo en una molécula, que no sea hidrógeno.
- La difractometría de rayos X requiere que el compuesto sea cristalino.
El otro problema es que un modelo de bola y varilla de una proteína contiene muchísimos átomos. La estructura se vuelve bastante complicada. El ejemplo aquí es una proteína bastante pequeña; su peso molecular es de solo unos 35,000 D (D significa Daltons; un Dalton es solo otro término para unidad de masa atómica o amu, pero se usa más comúnmente en bioquímica y química macromolecular). En proteínas más grandes, es difícil darle sentido a la pila de átomos que vemos en una estructura de bola y palo.
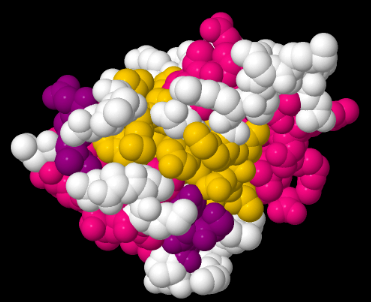
Del mismo modo, podríamos ver la proteína en un modelo de relleno de espacio. El modelo de llenado de espacio probablemente da una mejor imagen de cómo sería la molécula en términos de cuánto espacio ocuparía, o cuál sería su forma general. La ventaja de esta visión es que permite a un bioquímico ver cómo otras moléculas podrían interactuar con la proteína.
Por ejemplo, tal vez una proteína actúa como enzima; cataliza reacciones que son esenciales para nuestra supervivencia. La proteína tirosil fosfatasa bovina de bajo peso molecular es un ejemplo. Es bovino, lo que significa que se encuentra en vacas. Es una fosfatasa, lo que significa que elimina los grupos fosfato de otras proteínas. Ese es un evento regulatorio importante en bioquímica. Agregar un grupo fosfato o quitar un grupo fosfato de otra molécula, incluyendo otra proteína, a menudo actúa como un interruptor de “encendido/apagado”, para que un organismo pueda controlar cuándo ocurren ciertas reacciones. Esta es una tirosil fosfatasa, por lo que elimina específicamente el fosfato que se ha agregado a un residuo de tirosina en la proteína sustrato. Al observar el modelo de llenado de espacio, un bioquímico puede ver dónde puede encajar la tirosina del sustrato en la tirosil fosfatasa bovina de bajo peso molecular, para que la enzima pueda hacer su trabajo y cortar el fosfato.
El problema es que la mayoría de las proteínas se parecen más o menos a manchas al ojo inexperto. Para facilitar un poco las cosas, existen atajos que los bioquímicos utilizan a la hora de visualizar proteínas.
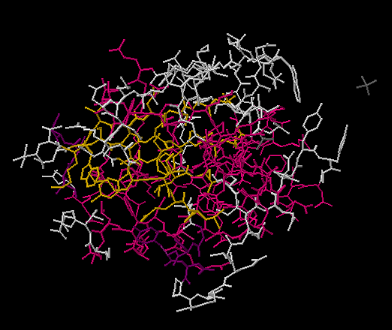
Una abreviatura utilizada en bioquímica a veces se llama estructura de estructura metálica. En una estructura de estructura metálica, ninguno de los átomos está etiquetado y se omiten todos los hidrógenos. Eso significa que es difícil distinguir un carbono de un nitrógeno. Sin embargo, las macromoléculas biológicas como el ADN y las proteínas están hechas de unos simples bloques de construcción. Un lector muy experimentado recordaría las estructuras de estos bloques de construcción. Ella podría decir qué átomos van en qué lugar a partir de las formas de los bloques de construcción.
Esa sigue siendo una imagen bastante complicada. Hay otras opiniones que se simplifican aún más.
Una proteína está hecha de aminoácidos unidos en una cadena. La cadena no cuelga en línea larga y recta. Se pliega en una bobina, y la forma de esa bobina influye en su actividad biológica. A veces un bioquímico puede estar interesado en cómo se bobina la cadena de aminoácidos. Una estructura de cadena principal solo muestra los átomos que están conectados directamente en una fila de un extremo de la cadena al siguiente. Cualquier átomo que cuelgue a lo largo de los lados de la cadena queda fuera.
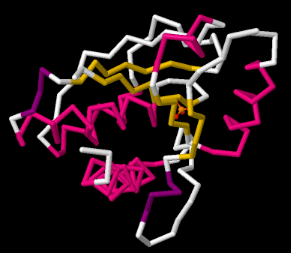
Esas partes que quedaron fuera son bastante importantes en cuanto a la función de la proteína. Son las partes que hacen que la alanina sea diferente de la lisina, por ejemplo. No obstante, si estás tratando de ver algunas formas básicas en la proteína, simplemente terminan interponiéndose en el camino en la imagen.
El orden en el que una cadena de aminoácidos se conecta entre sí se llama la estructura primaria de una proteína. Por ejemplo, los aminoácidos glicina, alanina, tirosona y arginina podrían estar conectados en el orden gly-ala-tyr-arg, o ala-gly-arg-tyr, o tyr-gly-ala-arg, y así sucesivamente. Estos compuestos son isómeros entre sí. Saber qué isómero tenemos nos dice algo sobre la proteína.
La forma en que se organiza la cadena se llama estructura secundaria. Muy a menudo, las proteínas se arrollan en espirales o hélices. Esta estructura helicoidal es similar a la del ADN. Otra posible estructura secundaria es una lámina beta, que es algo ondulada, un poco como una lámina de acero corrugado. Estas estructuras secundarias se mantienen en su lugar mediante enlaces de hidrógeno entre diferentes aminoácidos. (Puede ver el capítulo de relación estructura-propiedad para revisar los enlaces de hidrógeno).
En el dibujo de la columna vertebral de tirosil fosfatasa bovina de bajo peso molecular, se puede ver dónde partes de la cadena principal se enrollan en hélices alfa. Estas partes están codificadas en rosa en la imagen. También se pueden ver algunas secciones que forman hojas beta plisadas. Esas son las secciones amarillas, en zig-zag una al lado de la otra.
Por lo general, toda la proteína no adopta la misma estructura secundaria. Puede haber una sección que se bobina en una hélice y otra sección que forme una lámina beta. El dominio helicoidal y el dominio de lámina beta se disponen y se empaquetan de alguna manera. La disposición de los dominios en la proteína se denomina estructura terciaria.
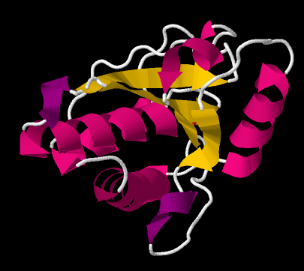
Ver la estructura terciaria en una proteína puede ser difícil incluso con una estructura de cadena principal. En cambio, se utilizan dibujos de cinta o estructuras de dibujos animados para transmitir cómo se disponen las láminas beta y las hélices en una proteína.
A veces una proteína está hecha de más de una cadena de aminoácidos. La disposición de diferentes bobinas de aminoácidos todos pegados entre sí se llama estructura cuaternaria. La tirosil fosfatasa bovina de bajo peso molecular es una estructura monomérica; solo está hecha de una cadena proteica, por lo que no vemos una estructura cuaternaria en este caso.