
9.1: La celda- Una visión general
- Page ID
- 80600

Probablemente hayas estudiado la célula muchas veces, ya sea en la secundaria, o en clases universitarias de biología. Hay muchos sitios web disponibles que revisan células procariotas (bacterianas), arqueales y eucariotas (ver enlaces en la parte inferior). Este tutorial está diseñado específicamente desde el punto de vista de la química. Explora cuatro clases de biomoléculas (lípidos, proteínas, ácidos nucleicos y carbohidratos) y describe de manera gráfica simplificada dónde se encuentran, elaboran y degradan en la célula (es decir, su historia). Se centra en las células eucariotas, que a diferencia de las células procariotas más simples, tienen orgánulos internos rodeados de membranas que compartimentalizan las reacciones químicas. Primero se presenta una visión general de la célula. Los capítulos posteriores se concentrarán en biomoléculas específicas.
Pensemos en una célula como una fábrica química que diseña, importa, sintetiza, usa, exporta y degrada una variedad de químicos (en el caso de la célula, estos incluyen lípidos, proteínas, ácidos nucleicos e hidratos de carbono). También debe determinar o detectar la cantidad de productos químicos crudos y terminados que tiene disponibles y responder a sus necesidades propias y externas incrementando o cerrando la producción. La figura de la celda de abajo y en otros sitios enlazados basados en ella se puso a disposición con el amable permiso de Liliana Torres.
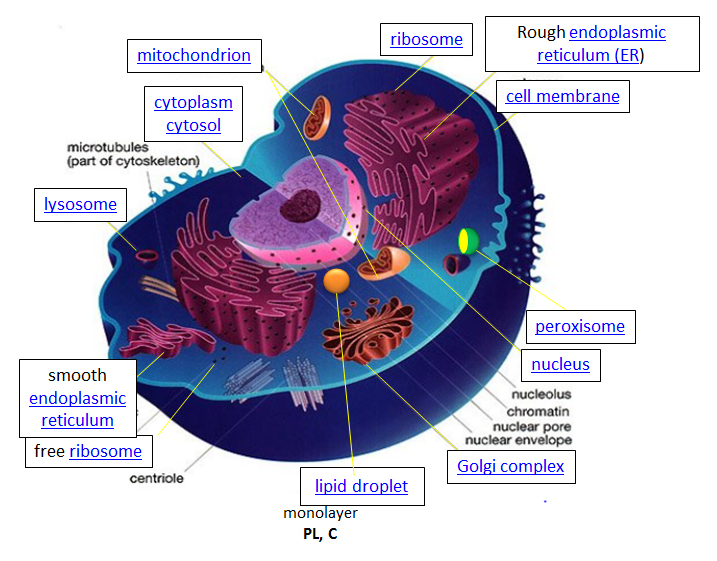
http://torresbioclan.pbworks.com/w/page/22377234/Spikefish%20About%20Cells
Uso con permiso de Liliana Torres. También en www.animalport.com/animal-cells.html
Diseño - El diseño de una célula reside principalmente en el plano para la célula, el código genético, que está compuesto por el ADN en el núcleo celular y una pequeña cantidad en las mitocondrias. Por supuesto, el plano del ADN debe ser leído (transcrito) por enzimas proteicas que a su vez fueron codificadas por el ADN. El código genético tiene el plan maestro que determina la secuencia de todas las proteínas celulares, que luego catalizan casi todas las demás actividades en la célula, incluyendo catálisis, motilidad, estructura arquitectónica, etc. A diferencia de ADN, ARN y polímeros proteicos, la longitud y secuencia de un polímero polisacárido es no impulsado por un molde sino por las enzimas que catalizan la polimerización.
Importación/Exportación: Muchos de los constituyentes químicos de la célula surgen no de la síntesis directa sino de la importación de moléculas pequeñas y grandes. Las moléculas importadas deben pasar a través de la membrana celular y en algunos casos a través de membranas adicionales si necesitan residir dentro de orgánulos unidos a la membrana. Las moléculas pueden moverse hacia la célula por difusión pasiva a través de la membrana, pero generalmente su movimiento es “facilitado” por un receptor de membrana. Las moléculas también pueden subir un gradiente de concentración en un proceso llamado “transporte activo”. Dada la naturaleza anfifílica de la bicapa (grupo de cabeza polar exterior, interior no polar), se esperaría que una molécula polar como la glucosa tenga dificultades para moverse a través de la membrana. De hecho, las proteínas transportadoras de membrana están involucradas en el movimiento de moléculas no polares y polares.
- transportadores, proteínas portadoras y permeasas: Estas proteínas de membrana mueven moléculas de ligando específicas a través de una membrana, típicamente por debajo de un gradiente de concentración. Las simulaciones informáticas de la difusión facilitada de lactosa a través de la membrana se muestran en el siguiente enlace. Animación de la difusión de lactosa a través del receptor LacY
(El enlace anterior e inmediatamente a continuación son del grupo de Biofísica Teórica y Computacional del Instituto Beckman, Universidad de Illinois en Urbana-Champaign. Estas simulaciones dinámicas moleculares se realizaron con VMD/NAMD/Biocore/JMV/otro soporte de software desarrollado por el Grupo con soporte de NIH).
- canales iónicos - Estas proteínas de membrana permiten el flujo de iones a través de las membranas. Algunos están permanentemente abiertos (no unidos) mientras que otros están abiertos o cerrados dependiendo de la presencia de ligandos que se unen al canal proteico y al ambiente local de la proteína en la membrana. El flujo de iones a través del canal procede en una dirección termodinámicamente favorecida, que depende de su concentración y gradientes de voltaje a través de la membrana.
- poros: Algunas membranas (nucleares, mitocondrias) ensamblan proteínas (como las porinas) para formar poros grandes pero regulados. Las porinas se encuentran en las membranas mitocondriales mientras que las nucleoporinas se encuentran en la membrana nuclear. Las moléculas pequeñas generalmente pueden pasar a través de estos poros de membrana, mientras que una grande se selecciona en función de su tendencia a formar fuerzas de atracción intermoleculares transitorias con las proteínas de poro. El siguiente enlace muestra la difusión del agua a través de la acuaporina. animación de la difusión del agua a través del canal de acuaporina,
- endocitosis: Partículas muy grandes (por ejemplo, LDL o Lipoproteínas de Baja Densidad o virus) pueden ingresar a una célula a través de un proceso llamado endocitosis. Inicialmente la LDL o virus se une a un receptor en la superficie de la célula. Esto desencadena una serie de eventos que conducen a la invaginación de la membrana celular en ese punto. Esto finalmente se pellizca para formar una vesícula endosómica que está rodeada por una proteína llamada clatrina. Los endosomas “tempranos” pueden recoger nuevas proteínas y otros constituyentes, así como desprenderlos a medida que se mueven y maduran a través de la célula. Durante este proceso de maduración, las bombas de proteínas en el endosoma conducen a una disminución en el pH endosómico que puede conducir a cambios de conformación en la estructura de la proteína y desprendimiento de proteínas. Finalmente, el endosoma “tardío” alcanza y fusiona con el lisosoma, un orgánulo interno que contiene enzimas degradativas. Los componentes no degradados como los ácidos nucleicos virales o el colesterol, se entregan a la célula. Este transporte también puede ir en la dirección inversa y reciclar los receptores a la membrana celular. Asimismo, las vesículas pellizcadas del complejo de Golgi pueden fusionarse con endosomas, sobreviviendo algunos componentes al proceso de reingresar al Golgi.
Sintetizar/Degradar: Las células tienen que sintetizar y degradar moléculas pequeñas, así como proteínas poliméricas más grandes, carbohidratos y ácidos nucleicos. Las vías anabólica (sintética) y catabólica (degradativa) a menudo están compartimentadas en tiempo y espacio dentro de una célula. Por ejemplo la síntesis de ácidos grasos se lleva a cabo en el citoplasma pero la oxidación de ácidos grasos se lleva a cabo en las mitocondrias. Las proteínas se sintetizan en el citoplasma o se completan en el retículo endoplásmico (para proteínas de membrana y exportadas) mientras que se degradan en el lisosoma o lo más importante en una gran estructura multimolecular en la célula llamada proteasoma.
Consideremos algunas características clave de una celda antes de entrar en los detalles.
Las células y sus compartimentos internos tienen concentraciones reguladas de iones e iones hidronio.
Como era de esperar, el pH del citosol (la sustancia acuosa que rodea a todos los orgánulos dentro de la célula) varía de aproximadamente 7.0-7.4, dependiendo del estado metabólico de la célula. Algunos orgánulos tienen transportadores de protones que pueden alterar significativamente el pH dentro de un orgánulo. Por ejemplo, el pH dentro del lisosoma, un orgánulo degradativo, es de aproximadamente 4.8. El colapso del gradiente de pH a través de la membrana mitocondrial interna es suficiente para impulsar la síntesis termodinámicamente desfavorecida de ATP.
En comparación con el líquido extracelular, la concentración de iones potasio es mayor dentro de la célula, mientras que las concentraciones de iones sodio, cloruro y calcio son mayores en el exterior de la célula (ver tabla a continuación). Estos gradientes de concentración son mantenidos por transportadores y canales iónicos y requieren gasto de energía en última instancia en forma de hidrólisis de ATP. Los cambios en estas concentraciones son parte integral del sistema de señalización utilizado por la célula para detectar y responder a los cambios en sus entornos externos e internos.
El agua se encuentra en forma masiva así como unida a macromoléculas como proteínas y polisacáridos. Se esperaría que estas aguas tuvieran diferentes propiedades. La siguiente tabla muestra las concentraciones de iones aproximadas en la celda.
| Ion | Interior (mm) | Exterior (mm) |
| Na + | 140 | 5 |
| K + | 12 | 140 |
| Cl- | 4 | 15 |
| Ca 2+ | 1 uM | 2 |
Las celdas tienen un marco interno que proporciona soporte estructural arquitectónico e interno.
La arquitectura “citoesquelética” de a (con “cables” moleculares y estructuras “similares a vigas”) no es diferente de una fábrica.

http://www.cybercom.net/~copters/trips/pictures/factory_inside.jpg http://en.Wikipedia.org/wiki/File:FluorescentCells.jpg
El marco interno o citoesqueleto de una célula, está compuesto por microfilamentos, filamentos intermedios y microtúbulos. Estas están compuestas por proteínas monoméricas que se autoensamblan para formar la arquitectura interna. Partes del citoesqueleto se pueden ver en la foto de arriba (tomada de Wikipedia).
En la imagen se muestran microfilamentos de monómeros de actina (que se tiñen con un fluoróforo rojo/naranja) y microtúbulos que ofrecen más soporte estructural hechos de monómeros de tubulina (teñidos de verde) junto con el núcleo teñido de azul. Los orgánulos son apoyados y organizados por el citoesqueleto (principalmente microtúbulos). Incluso la membrana celular está soportada por debajo de la valva interna por actina (naranja teñida) y microfilamentos de espectrina. Las proteínas motoras como la miosina (que se mueve a lo largo de microfilamentos de actina) y la dineína y cinesina (que se mueven a lo largo de los microtúbulos de tubulina) transportan carga (vesículas, orgánulos) de manera direccional La célula no es una colección desorganizada de moléculas y orgánulos. Más bien es un producto altamente organizado para una óptima producción química, uso y degradación.
Las celdas tienen una variedad de formas. Algunas células inmunes circulantes deben deslizarse a través de las células que recubren las paredes capilares para migrar a los sitios de infección. El mismo proceso ocurre cuando las células tumorales metastatizan y escapan a otros sitios del cuerpo. Para ello, la célula debe cambiar drásticamente de forma, una respuesta que requiere la disociación de los polímeros del citoesqueleto en monómeros que están disponibles posteriormente para su repolimerización.
La celda es un lugar asombrosamente concurrido.
En los laboratorios de química, normalmente trabajamos con soluciones diluidas de moléculas de soluto en un disolvente. Probablemente hayas escuchado que el cuerpo está compuesto por 68% de agua, pero la concentración de agua obviamente depende del entorno celular. Las moléculas de soluto como las proteínas y los carbohidratos están densamente empaquetadas. Las células están tan abarrotadas que el espacio entre moléculas más grandes como la proteína es menor que el tamaño de la proteína. Los estudios han demostrado que la estabilidad de una proteína se incrementa en tal condición, lo que ayudaría a mantener la proteína en el estado plegado, nativo. Otra consecuencia de las altas concentraciones es que promovería la unión o autoagregación de moléculas similares, así como las que no me gustan que desde una perspectiva de equilibrio no se producirían en soluciones diluidas. Por lo tanto, el estudio de biomoléculas en soluciones diluidas en el laboratorio puede no revelar las complejidades reales de las interacciones y actividades de la misma molécula in vivo. Recientemente los investigadores han agregado un copolímero neutro de sacarosa y epiclorhidrina a las células in vitro. Estas partículas indujeron la organización de moléculas extracelulares secretadas por la célula, formando una “matriz” extracelular organizada que indujo la organización de los microfilamentos en el interior de la célula así como induciendo cambios en la actividad celular. 1 La actividad enzimática in vitro de una enzima clave en la glucólisis aumenta drásticamente en condiciones de hacinamiento. 2 Otro resultado del hacinamiento puede ser la asociación espacial y temporal de enzimas clave en vías metabólicas específicas, permitiendo una difusión más fácil de sustratos y productos dentro de las enzimas colocalizadas.
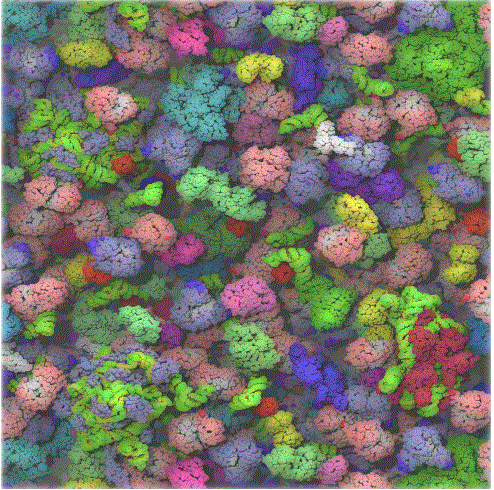
La simulación por computadora utilizó 50 tipos diferentes de las macromoléculas más abundantes del citoplasma de E. coli y 1008 moléculas individuales. Renderización del modelo de citoplasma al final de una simulación dinámica. El ARN se muestra como verde y amarillo. Esta cifra se preparó con VMD
Los componentes celulares experimentan transiciones de fase para formar subestructuras dentro de la celda.
Una pregunta despleja es cómo se forman las subestructuras dentro de una celda. Esto incluye no solo la biogénesis de orgánulos como las mitocondrias sino también partículas más pequeñas como gránulos de polisaacharida, gotitas lipídicas, partículas de proteína/ARN (incluyendo el ribosoma) así como el nucleolo del núcleo celular. Podría ser más fácil considerar este problema usando dos ejemplos del mundo lipídico, gotitas lipídicas y balsas de membrana. Usted está muy familiarizado con las transiciones de fase que ocurren cuando se agrega al agua un líquido no polar soluble ahorrador. A una concentración suficientemente alta, se excede la solubilidad del líquido no polar y se produce una transición de fase como lo demuestra la aparición de dos fases líquidas separadas. El mismo proceso ocurre cuando los triglicéridos se fusionan en gotitas lipídicas con proteínas asociadas en su exterior. Otro ejemplo ocurre dentro de una membrana celular cuando los lípidos con cadenas alquílicas saturadas se autoasocian con el colesterol de membrana (que contiene un sistema de anillo plano rígido) para formar una balsa lipídica caracterizada por una mayor eficiencia de empaquetamiento, rigidez y grosor que otras partes de la membrana. Estas balsas lipídicas a menudo reclutan proteínas involucradas en procesos de señalización dentro de las membranas celulares. Este proceso de separación de fases también se llama desmezcla líquido/líquido ya que dos sustancias “similares a líquidos” se separan.
Parece que las proteínas que interactúan con el ARN están compuestas por aminoácidos menos diversos y tienen estructuras más flexibles (“más líquidas) que permiten su interacción preferencial con el ARN para formar partículas grandes de ARN-proteína (como el ribosoma y otras estructuras de procesamiento de ARN) de una manera que imita el líquido /desmezcla líquida. Todas estas interacciones son solo manifestaciones de las diversas fuerzas intermoleculares que estudiaste en clases de química anteriores.
Ha estudiado la estructura de lípidos, proteínas, ácidos nucleicos y carbohidratos en clases introductorias de biología y/o química.
Una lista de sitios web y applets sobre la estructura celular eucariota:
- Tutorial celular de UCLA
- Simulación de células vegetales y animales (requiere Java)
- Descargar Applet del Apple Store para iPad, Itouch, iPhone: 3D Cell
Referencias:
1. Zeiger AS, Loe FC, Li R, Raghunath M, Van Vliet KJ (2012) El hacinamiento macromolecular dirige la organización de la matriz extracelular y el comportamiento de las células madre mesenquimales. PLoS ONE 7 (5): e37904. doi:10.1371/periodista.pone.0037904.
2. Apratim Dhara, Antonios Samiotakisb, Simon Ebbinghausa, Lea Nienhausa, Dirar Homouzb, Martin Gruebelea y Margaret S. Cheungb. La estructura, función y plegamiento de la fosfoglicerato quinasa están fuertemente perturbados por el hacinamiento macromolecular. Proc. Natl Acad Sci. www.pnas.org/cgi/doi/10.1073/pnas.1006760107
3. Anthony A. Hyman y Kai Simons. Más allá de las transiciones de fase de petróleo y agua en celdas. Ciencia 337, 1047 (2012); DOI: 10.1126/ciencia.1223728
4. McGuffee SR, Elcock AH (2010) Difusión, hacinamiento y estabilidad proteica en un modelo molecular dinámico del citoplasma bacteriano. PLoS Comput Biol 6 (3): e1000694. doi:10.1371/periodista.pcbi.1000694