
9.9: Captura de Carbono y Reserva de Carbohidratos
- Page ID
- 79338
Uno de los principales objetivos de la fotosíntesis es tomar el ATP y el NADPH producidos a través de la cadena de transporte de electrones y utilizarlos para almacenar energía en forma de carbohidratos. El evento clave en el almacenamiento de carbohidratos es la captura de dióxido de carbono de la atmósfera. Esa tarea la lleva a cabo la ribulosa-1,5-bisfosfato carboxilasa (generalmente abreviada como rubisco). Rubisco se encuentra dentro de los cloroplastos, en el estroma.

Rubisco es un agregado de dieciséis proteínas, pero contiene sólo dos tipos diferentes de proteínas; está compuesto por ocho moléculas de una proteína más ocho moléculas de la otra. Es una proteína enorme, con un peso molecular de 50 o 60 kDa. Su función principal es utilizar un azúcar de cinco carbonos, la ribulosa, como andamio sobre el que unir una molécula de dióxido de carbono. El resultado, después de la reducción con NADH, es un azúcar de seis carbonos.
En la imagen de abajo, puedes ver las moléculas de ribulosa-1,5-bisfosfato esperando su dióxido de carbono. En realidad hay ocho de ellos, pero están dispuestos en cuatro pilas cuando se ven desde este ángulo, con cuatro moléculas en el frente y cuatro detrás de ellas.
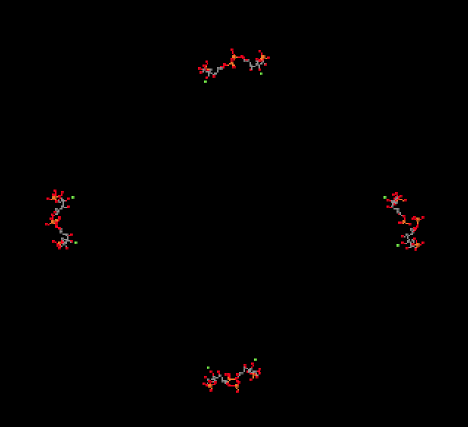
La captura del dióxido de carbono se logra a través de una simple reacción aldólica. En condiciones biológicas, esta reacción ocurre a través de la forma neutra enol de ribulosa-1,5-bisfosfato, en lugar del ion enolato aniónico, porque no hay una base lo suficientemente fuerte como para producir cantidades apreciables del ion enolato. Para llegar a la forma enol, se debe transferir un protón de una posición alfa al oxígeno de la cetona.

Proporcionar un mecanismo, con flechas curvas, para la captura del dióxido de carbono por el enol del bifosfato de ribulosa.
- Contestar
-
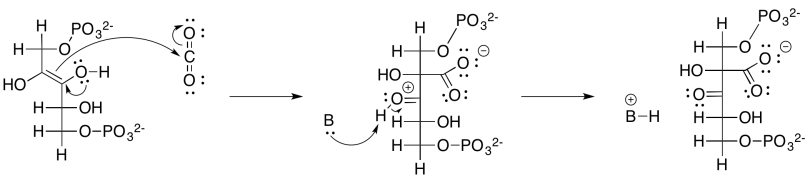
Ese intermedio, resultante de la reacción aldólica del bifosfato de ribulosa y el dióxido de carbono, no dura mucho. Se hidrata rápidamente para formar un diol geminal (grupos OH gemelos en un carbono), y el diol geminal se descompone a través de una reacción retro-aldólica. Recuerde, una reacción retro-aldólica es una reacción aldólica a la inversa. En lugar de un nucleófilo enolado que se agrega a un electrófilo de carbonilo, un oxígeno alcohólico dona pi para formar un nuevo carbonilo, y como resultado se produce un enolato como grupo saliente.

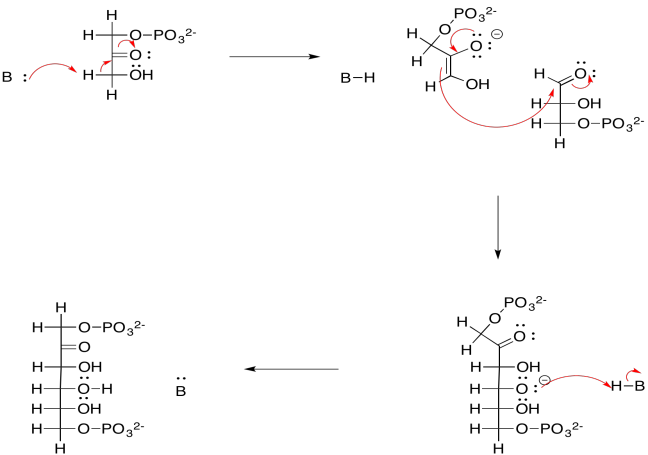
Proporcionar un mecanismo, con flechas curvas, para la escisión del diol geminal en dos moléculas de 3-fosfoglicerato.
- Contestar
-
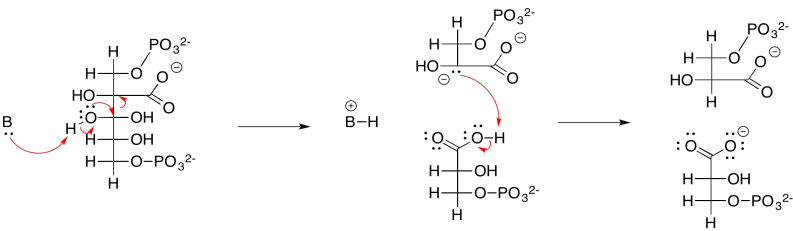
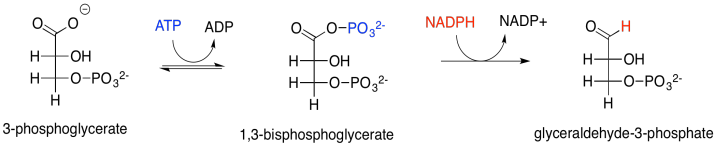
En este punto, entran en escena el ATP y el NADPH producidos en la cadena de transporte de electrones. El NADPH convierte el grupo carboxilato en un aldehído; en ese punto, el CO 2 se ha incorporado completamente a un azúcar normal, aunque el más pequeño que existe. El ATP se usa primero para activar el 3-fosfoglicerato. El NADPH no puede desplazar el óxido (O 2-) del 3-fosfoglicerato; el óxido es un grupo saliente terrible y el grupo carboxilato no es particularmente electrófilo con esa carga negativa colgando alrededor. El 3-fosfoglicerato sería un nucleófilo mucho mejor. Bueno, en lugar de reaccionar directamente con el NADPH, reacciona con el forst ATP. La fosfoglicerato quinasa activa el ATP, que luego está listo para comportarse como un electrófilo. El 3-fosfoglicerato cargado negativamente dona al grupo fosfato terminal en el ATP, desplazando al ADP. La quinasa ha hecho su trabajo, formando un fosfoanhídrido.
Pensemos en fosfoanhidros por un minuto. Son un poco como los anhidridos de ácido en estructura. Un anhídrido de ácido tiene un átomo de oxígeno entre dos carbonilos. Normalmente, la excelente capacidad de donación de pi de un átomo de oxígeno “protege” a un carbonilo unido de acercarse a los nucleófilos; en cierto sentido, el sitio electrófilo en el carbonilo ya está lleno por el par solitario del oxígeno. En un anhídrido, sin embargo, ese par solitario está en un tramado. Se divide entre dos carbonilos y así sólo puede donar la mitad a cualquiera de ellos. Como no le está haciendo mucho bien como donador pi, surge la otra mitad del comportamiento del oxígeno: también es un excelente aceptor sigma, porque es un átomo muy electronegativo. Eso deja al anhídrido ácido como un electrófilo muy bueno.

Por supuesto, los fosfoanhidridos se comportan exactamente de la misma manera, pero el oxígeno ostensiblemente donador de pi está atascado entre un grupo carbonilo y un grupo fosforilo. Es incapaz de donar pi de manera efectiva al carbonilo, por lo que su capacidad de extracción de electrones se hace cargo en su lugar. Un fosfoanhídrido es un excelente electrófilo, al igual que un anhídrido regular. Ambos están “en lo alto de la colina de esquí”, si recuerdas la analogía para la reactividad carboxiloide.
Por cierto, los polifosfatos tienden a ser bastante electrofílicos, también, que es uno de los factores que impulsan la alta reactividad del ATP.
El resultado es que el 3-fosfoglicerato bastante nucleófilo se ha transformado en un solo paso en el 1,3-bisfosfoglicerato altamente electrófilo, un ejemplo de un fosfoanhídrido. Eso hace que sea mucho más fácil para el NADPH donar un hidruro nucleofílico al carbonilo, formando un aldehído. El mecanismo exacto, catalizado por la gliceraldehído fosfato deshidrogenasa, implica una etapa de transferencia de grupo. El carbonilo se transfiere del grupo fosfato a un residuo de cisteína en la enzima. La ventaja de la transferencia de grupos es principalmente entrópica: en lugar de que el NADPH y el fosfoanhídrido se unan a la enzima y reaccionan juntos, en este punto el NADPH simplemente debe unirse con la enzima y reaccionar con la enzima misma, porque el grupo glicerato se ha unido covalentemente a la enzima.
Además, el nuevo grupo funcional formado durante la transferencia de grupo es un tioéster. Los tioésteres, como los fosfoanhidros, son excelentes electrófilos. Aunque los donantes pi de la segunda fila, oxígeno y nitrógeno, son excepcionalmente buenos para donar a carbonilos, los donantes pi de la tercera fila, como el cloro y el azufre, no lo son. En cambio, su electronegatividad relativamente alta activa el carbonilo. Eso significa que la transferencia de grupos ocurre sin ninguna pérdida de reactividad, lo cual es esencial para la catálisis.

Proporcionar un mecanismo para la reducción de NADPH de 1,3-bisfosfosglicerato.
- Contestar
-
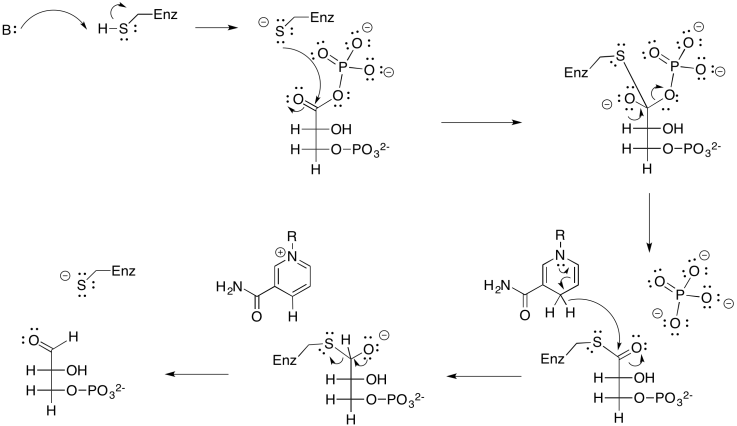
El gliceraldehído formado a través de la reducción por NADPH es el más simple de los carbohidratos; se puede pensar en él como la abuela de todos los azúcares, que se puede considerar que se forma a través de la extensión de la cadena de carbono del gliceraldehído. En este caso, realmente tenemos la forma fosforilada del azúcar, más que el propio azúcar. El gliceraldehído, o gliceraldehído-3-fosfato, se puede transformar fácilmente en otra molécula a través del tautomerismo. Ese es el movimiento de un protón de una posición a otra, y en este ejemplo conduce al dihidroxiacetonafosfato.
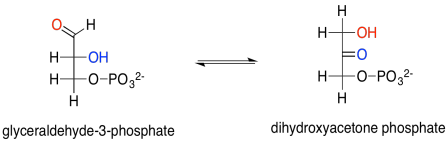
Mostrar el enol intermedio entre gliceraldehído-3-fosfato y dihidroxiacetona fosfato.
- Contestar
-
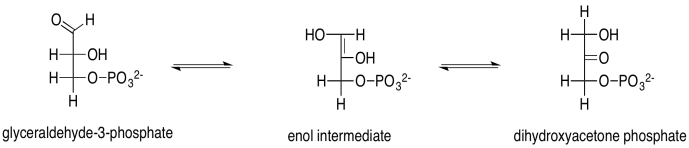
El gliceraldehído es, por supuesto, una aldosa, con su carbonilo al final de la cadena de tres carbonos, formando un grupo funcional aldehído. Los otros carbonos tienen grupos hidroxilo; eso es lo que lo convierte en un carbohidrato (un aldehído polihidroxilado o cetona). La dihidroxiacetona es una cetosa, con su carbonilo en el centro de la cadena, formando un grupo funcional cetona. Ese grupo está flanqueado por dos alcoholes. El hecho de que el gliceraldehído pueda convertirse en dihidroxiacetona es en parte significativo porque el potencial biosintético de este pequeño bloque de construcción se abre al permitirle formar el doble de estructuras.
Hay una serie de reacciones posteriores que tienen el efecto de interconvertir rápidamente estos azúcares en diferentes carbohidratos en una especie de reservorio de carbono. La planta puede aprovechar este charco de carbono para diversos fines, ya sea para enviar azúcares a otra parte de la planta o para regenerar un poco más de ribulosa con el fin de agarrar más dióxido de carbono. Al agarrar más dióxido de carbono, ese charco de carbono se hace cada vez más grande. La serie de reacciones que gobiernan este reservorio de carbono se llama el ciclo Calvino. Realmente es más una red que un ciclo, con moléculas capaces de saltar de un lado a otro entre varias partes del ciclo, en lugar de todo lo que sigue monótonamente a un paso cerrado.
Por ejemplo, la reacción aldólica de esas dos moléculas de fosfato de triosa —dihidroxiacetona fosfato y gliceraldehído-3-fosfato— conduce a fructosa-1,6-bisfosfato. Esta reacción se logra a través de la ayuda de una enzima, la aldolasa. La fructosa, o azúcar de frutas, es uno de los pocos azúcares que se absorben fácilmente a través de nuestro sistema digestivo.
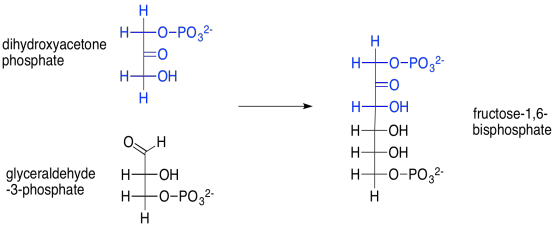
Proporcionar un mecanismo para la formación de fructosa-1,6-bisfosfato, arriba
- bajo condiciones básicas
- bajo condiciones ácidas
- Contestar a
-
a)
- Respuesta b
-
b)

Muchas reacciones biológicas aldólicas ocurren realmente a través de un intermedio de enamina.
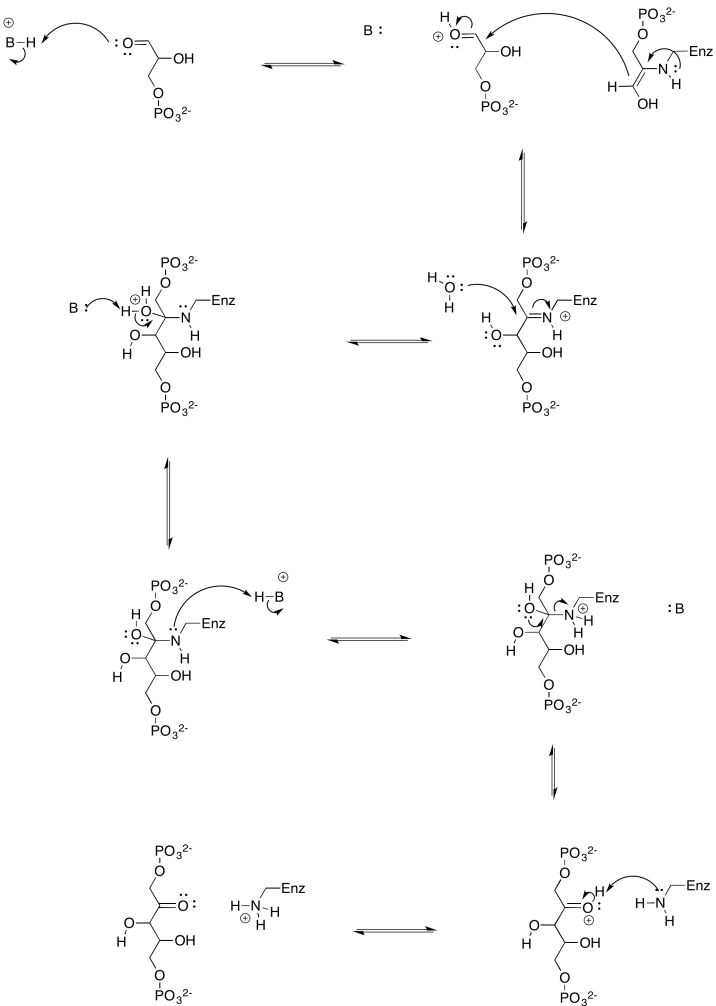
- Proporcionar un mecanismo para la formación de una enamina a partir de fosfato de dihidroxiacetona.
- Proporcionar un mecanismo para la formación de fructosa-1,6-bisfosfato a través de la enamina de fosfato de dihidroxiacetona.
- Contestar a
-
a)

- Respuesta b
-
b)
¿Por qué se usa la catálisis de enamina en vías bioquímicas? ¿Qué lo hace más rápido que una vía normal de enol?
- Contestar
-
La vía regular del enol es impulsada por la pi-donación de un átomo de oxígeno. La vía de la enamina es impulsada por pi-donación a partir de un nitrógeno. El nitrógeno es menos electronegativo y mejor donante pi que el oxígeno, por lo que la vía de la enamina es más rápida.
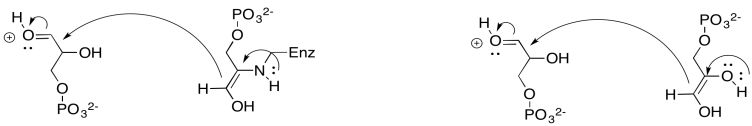
El siguiente paso en el ciclo Calvino es solo una desfosforilación, llevada a cabo con la ayuda de una fosfatasa. En este caso, la enzima se llama fructosa-1,6-bisfosfatasa. Las fosforilaciones y desfosforilaciones de sustratos, como la fosforilación y la desfosforilación de enzimas, son a menudo pasos reguladores clave, necesarios para promover una reacción que involucra a una enzima.

Al siguiente paso. Aquí es cuando las cosas se ponen un poco funky. Dos moléculas de carbohidratos se someten a una especie de metátesis juntas. En una reacción de metátesis, los compuestos se dividen y recombinan de una nueva manera. En esta reacción, la fructosa-6-fosfato se combina con otro gliceraldehído-3-fosfato. Después de la reacción, los dos azúcares viejos se han cambiado por dos nuevos: eritrosa-4-fosfato y xilulosa-5-fosfato. Parece que la fructosa se ha dividido y dos de sus carbonos se han conectado con la cadena de gliceraldehído.
¿Cómo sucede eso? La parte principal de la molécula de fructosa-6-fosfato se convierte en eritrosa-4-fosfato. Se puede imaginar que mucho sucede si el grupo hidroxi del carbono número tres realiza una donación de pi para formar el nuevo grupo aldehído. Ese es el grupo OH más alto, en negro, en fructosa-6-fosfato. Eso explica la formación de eritrosa-4-fosfato. El problema es, que la donación de pi requeriría que algo deje carbono tres. El grupo lábil sería un carbonilo aniónico. Probablemente no hayas visto antes a tal grupo que se va, por una buena razón.
No abandonemos esta idea, todavía. Si ese grupo de salida sí se formó (y no lo hace), eso explicaría la formación de xilulosa-5-fosfato, también. El anión a base de carbonilo, si actuara como nucleófilo, formaría un nuevo enlace carbono-carbono, abriendo el carbonilo del gliceraldehído-3-fosfato para formar el grupo hidroxi en lo que ahora es el carbono tres de xilulosa-5-fosfato (más arriba a la izquierda, en negro).
Hay más en esta reacción que un solo paso. El problema es que necesitamos formar un mejor grupo de salida. Eso requerirá un esfuerzo extra. La adición del iluro de pirofosfato de tiamina (TPP) ayudará. Al agregar temporalmente al carbonilo de fructosa-6-fosfato, podemos sortear el problema de tener una forma aniónica en el carbono carbonilo.
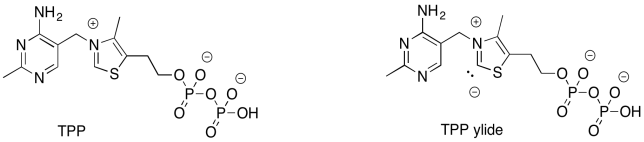
Esta misma estrategia surge durante el ciclo del ácido cítrico (o ciclo de TCA), donde es posible que haya visto antes el iluro de TPP. En ese ciclo, la descarboxilación del piruvato es imposible porque no hay carbono alfa sobre el cual estabilizar el anión que se forma vía descarboxilación. La carga negativa sobre el carbono carbonílico puede parecer una buena idea, porque está cerca del oxígeno electronegativo. Sin embargo, a diferencia de una posición alfa, el par solitario no puede deslocalizar completamente en el oxígeno. Eso es porque el par solitario asociado con el anión —el par solitario que se forma a partir del enlace roto— debe ser ortogonal al enlace pi.
El TPP viene al rescate. La adición de iluro de TPP a la cetona instala un ion iminio, convirtiendo el carbono carbonilo anterior en una posición alfa (es alfa a una imina en lugar de un carbonilo, pero funciona de la misma manera). El anión requerido ahora está completamente estabilizado por resonancia.
En la reacción de transcetolasa, el anión también necesita estabilizarse en la posición carbonilo. El iluro de TPP puede agregarse al carbonilo del azúcar, instalando una imina junto al carbono carbonilo anterior. Una vez realizada la reacción, el TPP puede ser desplazado nuevamente para formar un nuevo carbonilo sobre xilulosa-5-fosfato. Todo este proceso es catalizado por una enzima, la transcetolasa.
Proporcionar un mecanismo para la conversión mediada por TPP de fructosa-6-fosfato y 3-gliceraldehído fosfato en eritrosa-4-fosfato y xililosa-4-fosfato.
- Contestar
-
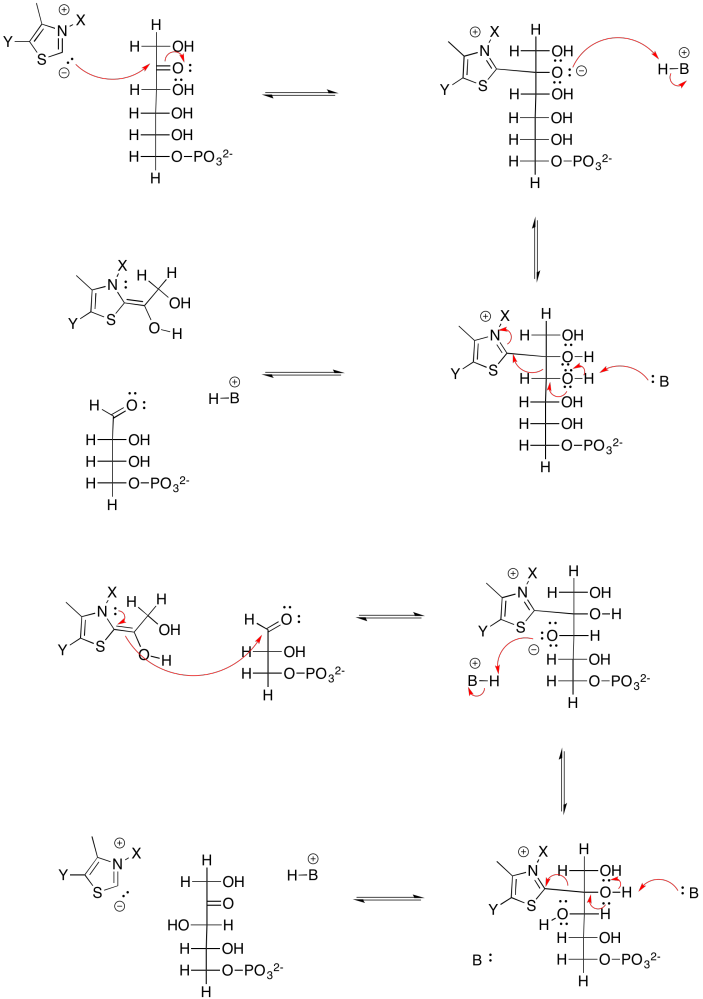
Esa no es la única reacción catalizada por la transcetolasa. La sedulosa-7-fosfato también reacciona con gliceraldehído-3-fosfato, en este caso formando ribulosa-5-fosfato y xilulosa-5-fosfato. Tenga en cuenta que primer producto: ribosa. La ribosa es la forma cetosa del azúcar aldosa, la ribulosa. La ribulosa fue el azúcar con el que comenzamos; su forma de bisfosfato fue absorbida por rubisco para atrapar CO 2. Estamos muy cerca de completar un ciclo, y estamos casi listos para capturar otra molécula de dióxido de carbono, agregando más stock a la reserva de carbono.
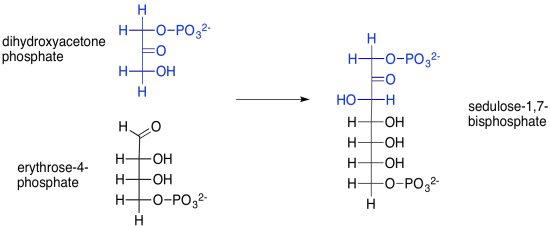
Pero, ¿de dónde viene esa sedulosa, de todas formas? Ese es el producto de otra reacción aldólica, esta vez entre fosfato de dihidroxiacetona y eritrosa-4-fosfato. La eritrosa-4-fosfato fue el otro producto de nuestra primera reacción transcetolasa.
Estamos a punto de terminar el ciclo completo de Calvino. Primero, la ribosa-5-fosfato solo necesita ser convertida en ribulosa-5-fosfato. Eso conlleva un mecanismo ceto-enol, en el que se transfiere un protón alfa al oxígeno carbonilo. La reacción se acelera en presencia de la enzima, ribosa fosfato isomerasa.
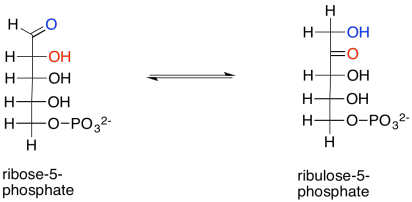
Mostrar el enol intermedio entre ribosa-5-fosfato y ribulosa-5-fosfato.
- Contestar
-

La ribulosa también se produce en otra reacción dentro del ciclo de Calvino. La ribulosa-5-fosfato es solo el epímero de xilulosa-5-fosfato. Es decir, es el estereoisómero formado al retirar un protón de la posición alfa quiral de xilulosa-5-fosfato y ponerlo de nuevo en el lado equivocado. La configuración en esa posición se cambia de (S) a (R), produciendo en su lugar la ribulosa-5-fosfato diastereomérico. Esta reacción reversible es emprendida por la enzima, ribulosa fosfato epimerasa.

La xilulosa-5-fosfato fue uno de los productos de ambas reacciones catalizadas por transcetolasa; una de esas reacciones también produjo ribosa-5-fosfato. Eso significa que la misma reacción proporciona dos precursores a la ribulosa-5-fosfato (xilulosa-5-fosfato y ribosa-5-fosfato) y otra reacción, catalizada por la misma enzima, produce una tercera (otra molécula de xilulosa-5-fosfato).
El nombre, ribulosa fosfato epimerasa, sugiere que la misma enzima realmente lleva a cabo la reacción inversa: la conversión de ribulosa-5-fosfato en xilulosa-5-fosfato. ¿Cómo podría regularse una reacción tan reversible?
- Contestar
-
Las reacciones que están en equilibrio suelen estar bajo el control del sustrato. Debido a que la enzima puede catalizar la reacción en ambas direcciones, la dirección de la reacción impulsada por la enzima está controlada por la cantidad relativa de reactivos a cada lado de la reacción. Por ejemplo, si la concentración de xilulosa aumentara mucho, entonces la cantidad de xilulosa unida por la enzima también aumentaría, por lo que la enzima desplazaría la xilulosa en ribulosa y con ello mantendría el equilibrio. Si la concentración de ribulosa aumentara mucho, entonces la enzima uniría más ribulosa y la ribulosa se convertiría en xilulosa.
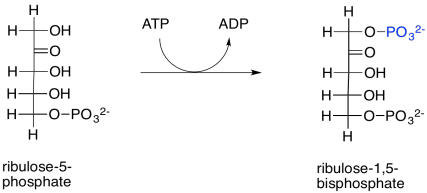
El ribulosa-5-fosfato estará listo para unirse al rubisco y tomar una molécula de dióxido de carbono tan pronto como sea fosforilada por la ribulosa fosfato quinasa. Esta reacción es en gran parte irreversible como está escrito debido al gasto de ATP.
Estructuras cristalinas de rayos X: Taylor, T.C.; Andersson, I. La estructura del complejo entre rubisco y su sustrato natural ribulosa 1,5-bisfosfato. J. Mol. Biol. 1997 265: 432-444. Imágenes obtenidas a través del Banco de Datos de Proteínas RCSB (1RCX).