
20.20: La Doble Hélice
- Page ID
- 75112

Hay más en la estructura del ADN que solo la secuencia primaria de bases nitrogenadas. La estructura secundaria también juega un papel bioquímico crucial. Cada molécula de ADN consiste en dos cadenas de nucleótidos envueltas una alrededor de la otra en una doble hélice y unidas por enlaces de hidrógeno. Este enlace de hidrógeno involucra solo las bases nitrogenadas. Cada una de las bases de purina puede formar enlaces de hidrógeno con una y solo una de las bases de pirimidina.
Así, la adenina puede formar enlaces de hidrógeno con timina y guanina con citosina, como se muestra en la Figura\(\PageIndex{1}\). Tenga en cuenta que en ambos casos hay una coincidencia exacta de átomos de hidrógeno en una base con átomos de nitrógeno u oxígeno en la otra. Obsérvese también que la distancia del enlace del azúcar al enlace del azúcar a través de cada uno de los pares de bases en la Figura\(\PageIndex{1}\) es casi exactamente la misma Esto explica por qué solo estas dos combinaciones ocurren en el ADN. Otras combinaciones (es decir, adenina-citosina) no son tan favorables energéticamente.
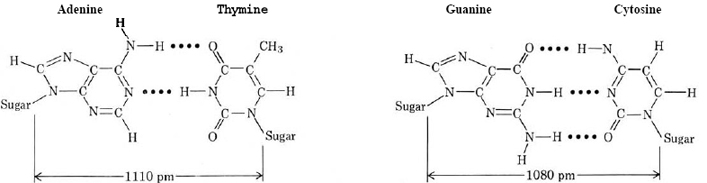
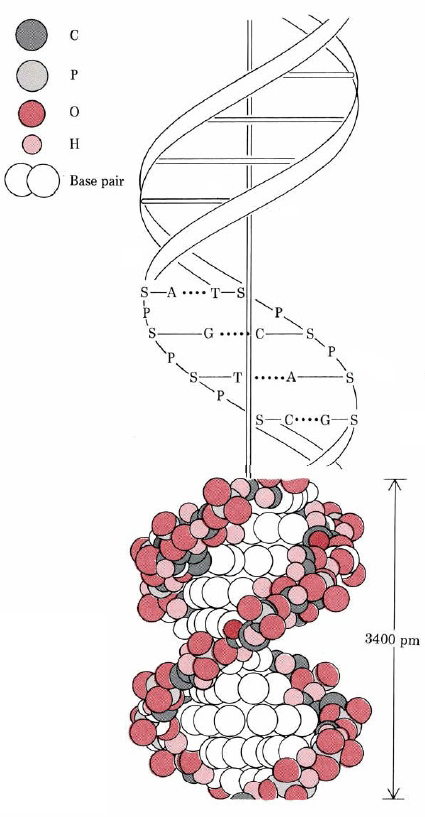
El espaciamiento entre los pares de bases es de 340pm, y hay 10.5 pares de bases en una vuelta completa de la hélice.
Se dice que las dos cadenas de nucleótidos en la doble hélice son complementarias entre sí. Debido al emparejamiento exacto de las bases siempre podemos decir las secuencias de bases en una cadena de esa en la otra. Así, si las primeras seis bases de una cadena son AGATCC, sabemos que las primeras seis bases en la otra serán TCTAGG. Ambas cadenas son, por tanto, representaciones alternativas de una misma información. Si una o dos bases quedan fuera de lugar en cualquiera de las cadenas, esto puede reconocerse debido a la falta de coincidencia con la hebra complementaria. Las enzimas reparadoras pueden entonces corregir la secuencia de bases a lo largo de la hebra incorrecta. Un punto final a hacer es que las dos hebras son antiparalelas. Esto significa que una hebra, de abajo hacia arriba, va del carbono 5' al carbono 3', mientras que la hebra complementaria va de 3' a 5' de abajo hacia arriba.
Este modelo de doble hélice para ADN fue sugerido por primera vez en 1953 por James D. Watson (nacido en 1928) y Francis Crick (1916 a 2004). Fue un hito importante en la historia de la ciencia, ya que marcó el nacimiento de un nuevo campo, la biología molecular, en el que las características de los organismos vivos podrían por fin comenzar a explicarse en términos de la estructura de sus moléculas. En 1962 Crick y Watson compartieron el Premio Nobel con M. F. H. Wilkins, cuyos datos cristalográficos de rayos X les habían ayudado a formular su modelo. Rosalind Franklin (1920-1958) que realizó los experimentos de cristalografía de rayos X no ganó el Premio Nobel, ya que no se otorgan póstumamente, sino que deben incluirse en cualquier discusión sobre el descubrimiento de la doble hélice. Un relato fascinante de este descubrimiento, que no siempre pone al autor en una luz favorable, se puede encontrar en el libro de Watson “La doble hélice”.