
2.8: Día 16- ADN y Lípidos
- Page ID
- 78952

15
Día 16: ADN y lípidos
D16.1 ADN
El ácido desoxirribonucleico (ADN) consiste en dos cadenas poliméricas que se enrollan una alrededor de la otra, formando una doble hélice. Las unidades monómeras de las cadenas de ADN se denominan nucleótidos. Un nucleótido consiste en un grupo fosfato y una nucleobase unida a un azúcar (desoxirribosa).
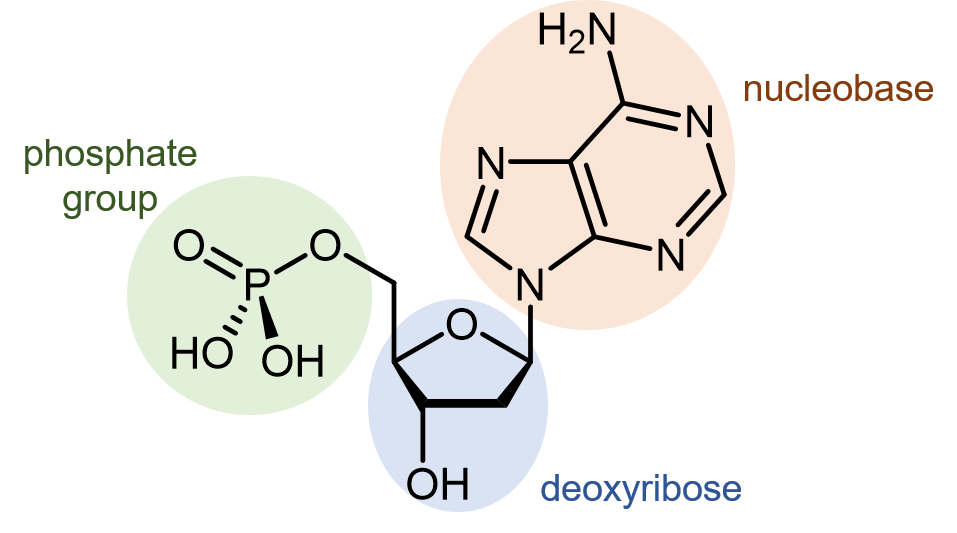
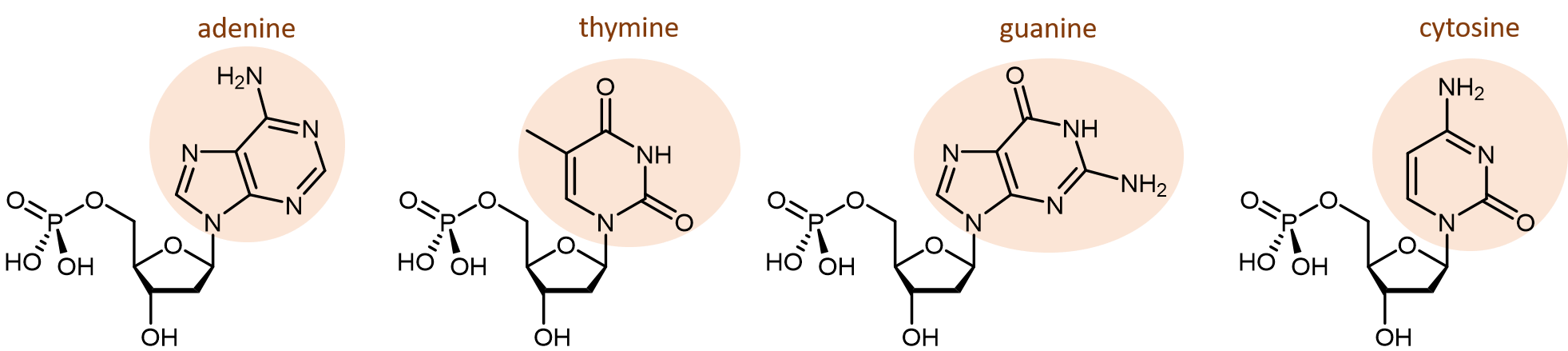
Hay cuatro nucleobases diferentes que se encuentran en el ADN. Se llaman adenina (abreviada A), timina (T), guanina (G) y citosina (C). Los nucleótidos que contienen esas nucleobases se denominan monofosfato de desoxiadenosina, monofosfato de desoxitimidina, monofosfato de desoxiguanosina y monofosfato de desoxicitidina (haga clic en cada nombre para ver un modelo molecular giratorio).
La reacción de polimerización que forma una cadena de ADN es una reacción de condensación entre el grupo fosfato y la desoxirribosa (específicamente el grupo -OH), lo que da como resultado la formación de enlaces fosfodiéster que unen los nucleótidos. En organismos que contienen ADN el pH es de aproximadamente 7, y a ese pH se ionizan los grupos fosfato. Así, el ADN es un anión poliatómico enorme con una carga negativa por cada unidad monomérica a lo largo de la cadena. La carga negativa se equilibra con cargas positivas en iones metálicos, moléculas de proteína y otros cationes que rodean el ADN.
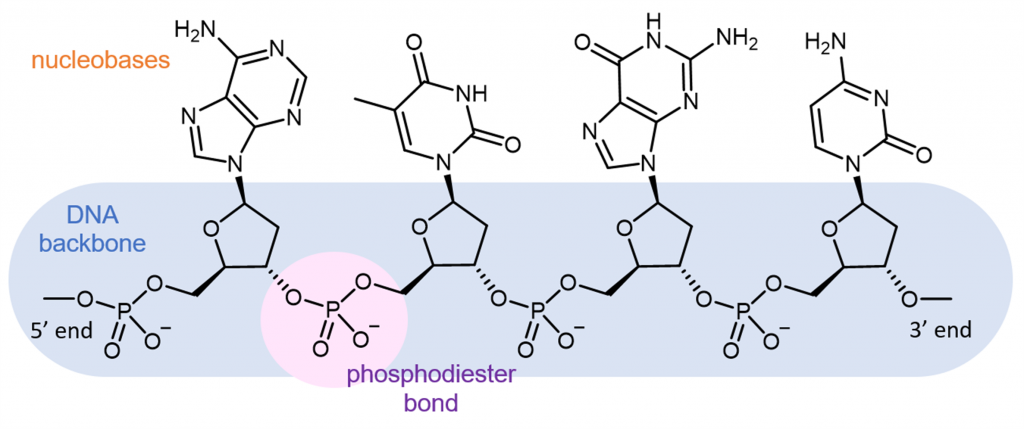
Cada cadena polimérica de ADN tiene un esqueleto de grupo de azúcar alterno y enlace fosfodiéster, con nucleobases aromáticas que contienen nitrógeno que forman cadenas laterales unidas a la cadena principal.
Una cadena de ADN tiene un grupo fosfato libre en un extremo (llamado extremo 5' porque el fosfato está unido a la posición carbono-5 en la ribosa) y un grupo hidroxilo libre (-OH) en el otro extremo (llamado extremo 3' porque el hidroxilo está unido a la posición carbono-3 en la ribosa). Por convención, las secuencias se nombran de 5′ a 3′, y solo las nucleobases están incluidas en el nombre. Por ejemplo, la molécula mostrada anteriormente es ATGC (no CGTA).
Dos cadenas individuales de ADN se combinan para hacer el anión molecular de doble hélice (típicamente llamado solo la molécula de ADN). Las hebras corren en direcciones opuestas: en un extremo de la doble hélice está el extremo 5' de una hebra y el extremo 3' de la otra hebra. Los dos hilos se envuelven entre sí para formar dos hélices entrelazadas. La figura giratoria muestra cómo se orientan las dos hélices. Tenga en cuenta que una de las brechas entre las hebras entrelazadas es más ancha que la otra brecha. Las dos cadenas separadas se mantienen en esta orientación mediante enlaces de hidrógeno entre nucleobases en el centro de la molécula.
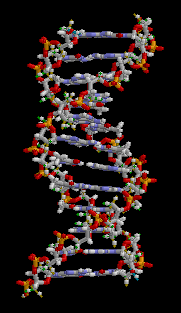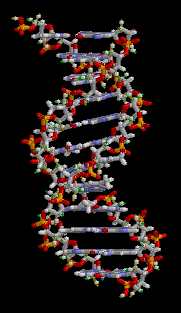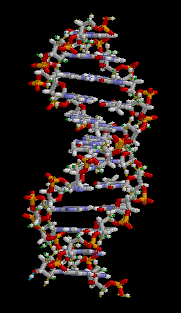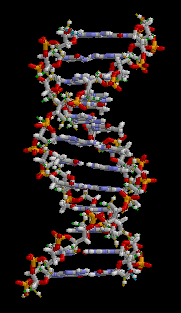
Emparejamiento de bases D16.2
Una de las cosas más notables del ADN es que la cantidad de adenina siempre es igual a la cantidad de timina y la cantidad de guanina siempre es igual a la cantidad de citosina. De un organismo a otro, la cantidad de adenina puede variar, pero la cantidad de timina siempre varía de la misma manera. Estas igualdades implican que las nucleobases ocurren en pares: adenina emparejada con timina y guanina emparejada con citosina. Estos pares de nucleobases se denominan pares de bases complementarias.
De seis combinaciones posibles, ¿por qué estas dos son las parejas complementarias? Primero, las nucleobases tienen diferentes tamaños: T y C tienen anillos simples y son más pequeños; A y G tienen anillos dobles y son más grandes. Si A y G se emparejan, su mayor tamaño obligaría a la doble hélice a abultarse. Si T y C se emparejan, su tamaño más pequeño provocaría pellizcos de la doble hélice. Solo un emparejamiento entre una nucleobase de un solo anillado y una nucleobase de doble anillado tiene el espaciado consistente adecuado.
Esto deja cuatro combinaciones posibles: A-C, A-T, G-C y G-T. De estos, los pares A-T y G-C maximizan el número de enlaces de hidrógeno a través del eje helicoidal compartido:
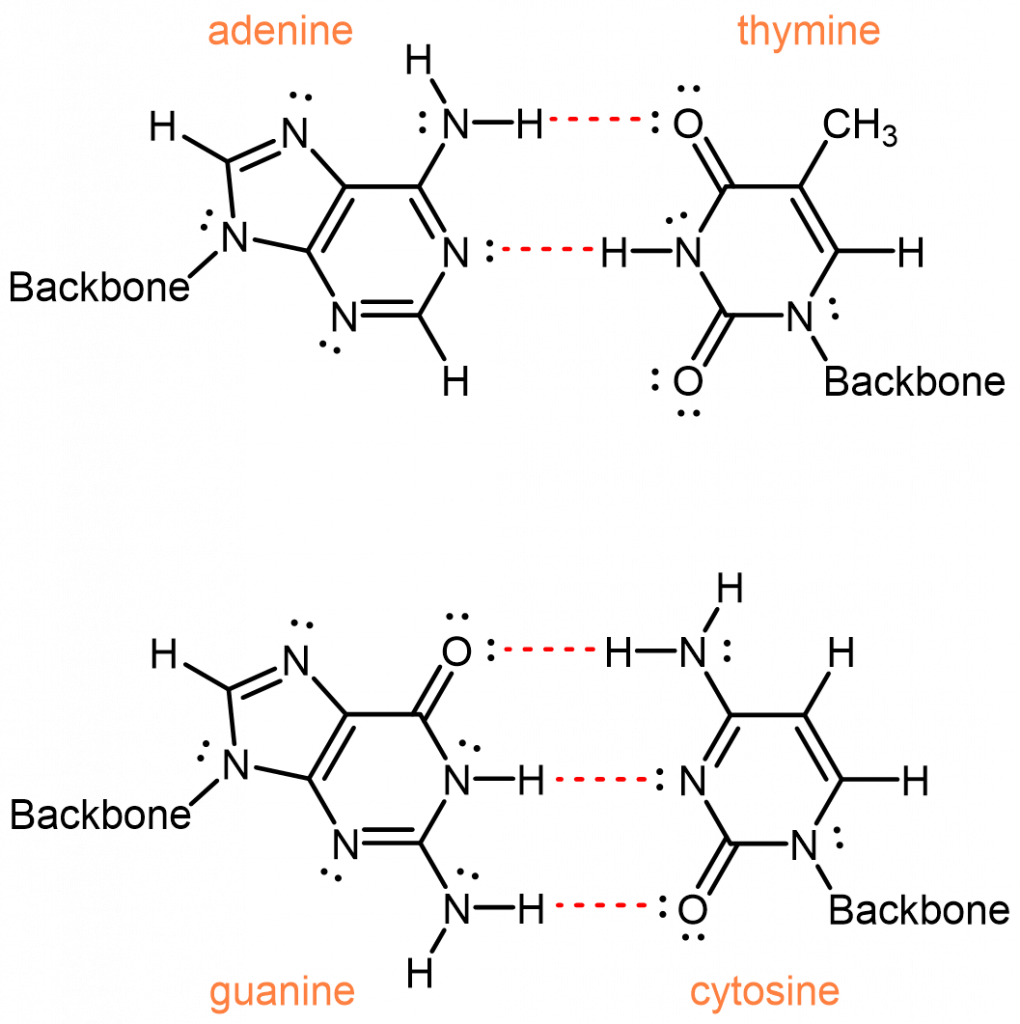
Los emparejamientos A-C y G-T, por otro lado, tendrían menos interacciones de enlaces de hidrógeno. Para lograr cualquier enlace de hidrógeno, tendría que haber un posicionamiento incómodo de las nucleobases entre sí.
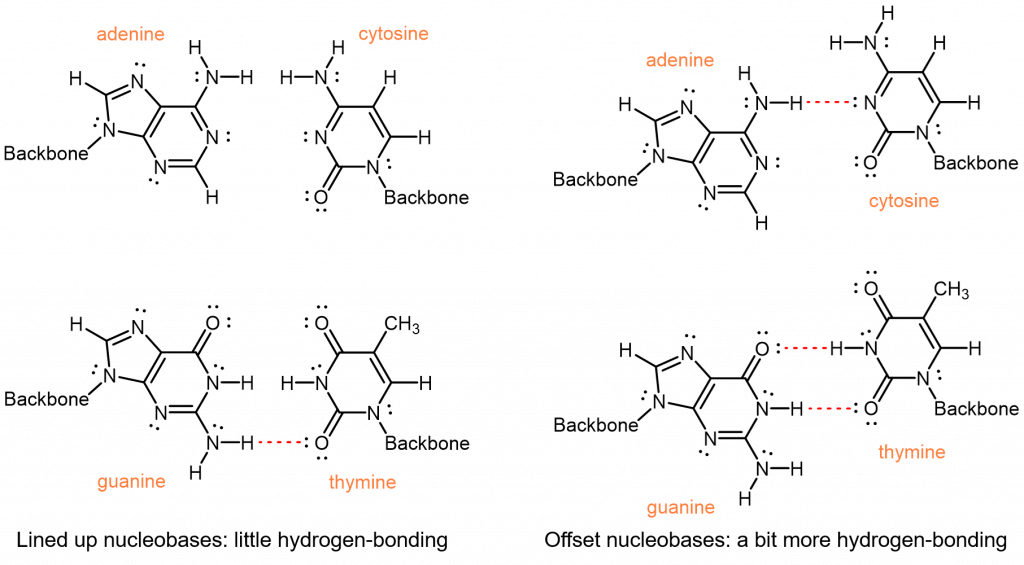
Por lo tanto, las interacciones intermoleculares, específicamente los enlaces de hidrógeno eficientes, entre adenina y timina, y entre guanina y citosina, dan como resultado la especificidad observada para los pares de bases complementarios.
Debido a la naturaleza del emparejamiento de bases complementarias, si conoces la secuencia de una hebra de ADN, puedes predecir la secuencia de la hebra que se emparejará con, o “complementarla”.
Durante la replicación del ADN, las hebras de la doble hélice del ADN se separan primero. Luego, las enzimas catalizan la síntesis de nuevo ADN en la dirección 5' a 3', utilizando las dos cadenas originales como modelos, o plantillas, para las cadenas complementarias. Dos moléculas de ADN completas, cada una copia exacta del original, son el resultado.
Debido a la especificidad del emparejamiento de bases y una actividad correctora de la enzima que sintetiza nuevo ADN, menos de un error en 100 millones se incorpora a las nuevas cadenas de ADN. Así, la información genética se transmite con precisión a la siguiente generación.
Lípidos D16.3
Las grasas y aceites forman parte de una clase de biomoléculas llamadas lípidos, que se definen vagamente como biomoléculas que son insolubles en agua pero solubles en disolventes orgánicos como el hexano o el cloroformo. Aquí se discuten dos clases importantes de lípidos: glicerolípidos y fosfolípidos. Las estructuras moleculares de los lípidos dan lugar a diferentes interacciones intermoleculares que influyen en sus propiedades físicas y funciones biológicas.
Glicerolípidos D16.4
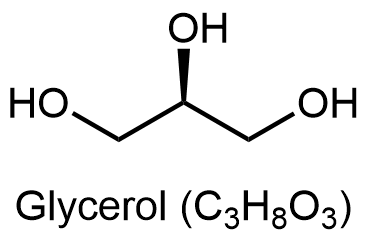
Los glicerolípidos están compuestos por glicerol y ácidos grasos. El glicerol tiene tres átomos de carbono, cada uno de los cuales tiene un grupo hidroxilo (-OH) unido a él:
Los ácidos grasos son cadenas hidrocarbonadas largas y no ramificadas con un grupo ácido carboxílico en un extremo.
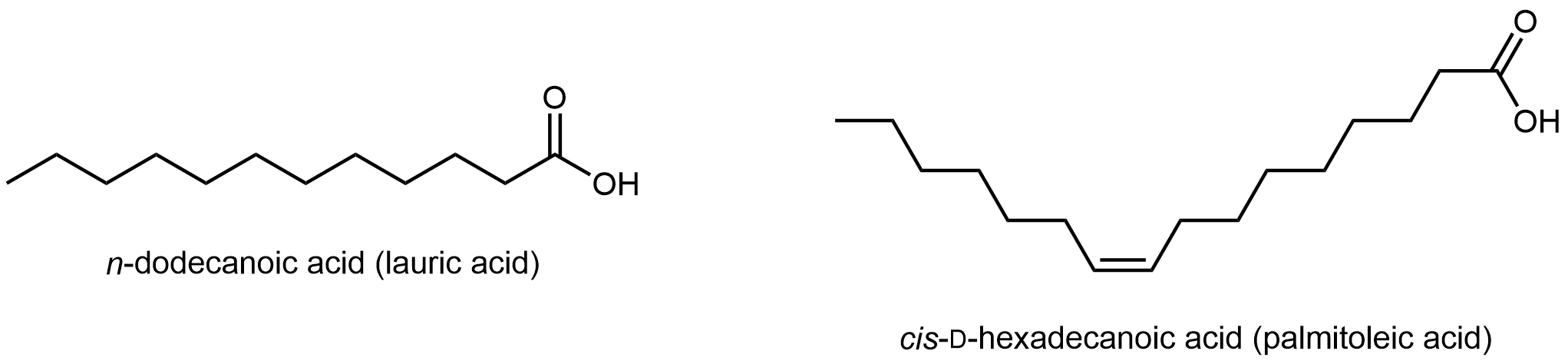
Los ácidos grasos se nombran sistemáticamente en función del número de átomos de carbono y los dobles enlaces C=C en la cadena. Pero sus nombres comunes, como el ácido láurico, suelen utilizarse para identificarlos.
Tenga en cuenta que solo hay enlaces simples C-C en el ácido láurico; similar a los hidrocarburos, dicho ácido graso es un ácido graso saturado. En contraste, el ácido palmitoleico tiene un doble enlace C=C, es decir, tiene un grado de insaturación. El enlace π puede sufrir la adición de H2 y formar un ácido graso saturado. Los lípidos elaborados a partir de ácidos grasos saturados e insaturados tienen diferentes propiedades.
Los glicerolípidos se forman cuando tres ácidos grasos se unen al glicerol. Catalizado por enzimas, cada grupo hidroxilo en glicerol puede sufrir condensación con el grupo ácido carboxílico de un ácido graso, formando un enlace éster.
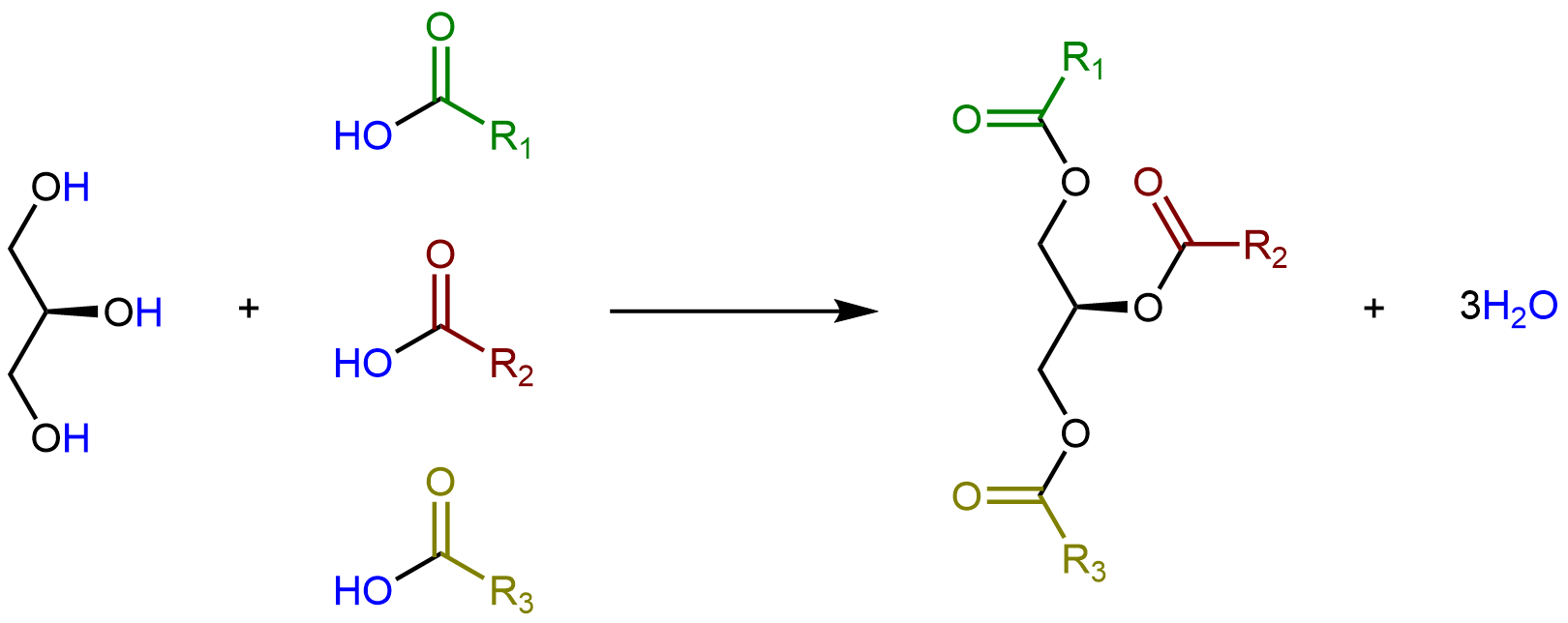
Las longitudes de las cadenas de ácidos grasos (por ejemplo, R1, R2, R3) y el número de dobles enlaces C=C dentro de ellas determinan el punto de fusión de un glicerolípido. Las cadenas más largas tienen fuerzas de dispersión de Londres más fuertes que las cadenas más cortas, lo que lleva a un punto de fusión Las cadenas saturadas pueden empacar entre sí regularmente, dando lugar a fuerzas de dispersión de Londres más fuertes entre las cadenas y puntos de fusión más altos. Las cadenas insaturadas, especialmente con dobles enlaces cis, que obligan a la cadena a doblarse, tampoco pueden apilarse juntas, por lo que las fuerzas de dispersión de Londres son más débiles.
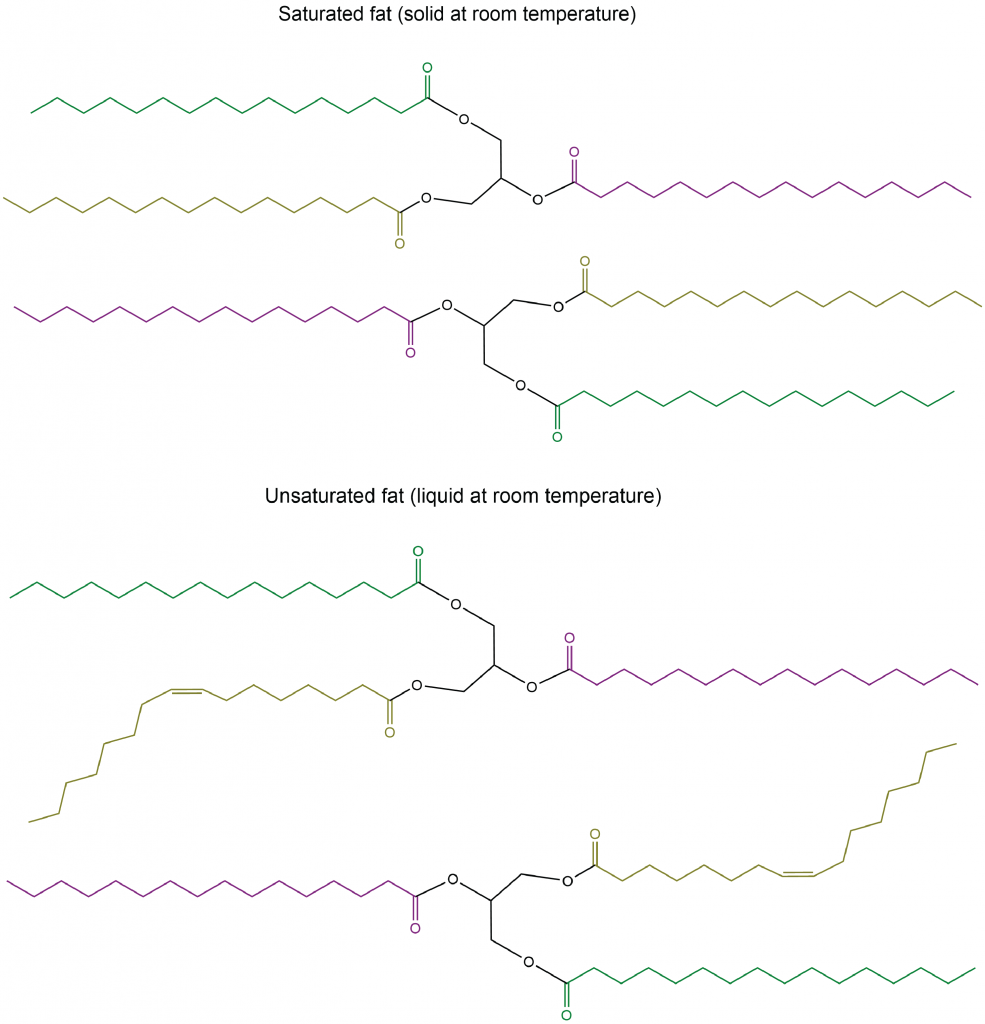 Figura 2. Aquí se muestran ejemplos de glicerolípidos saturados e insaturados (dos moléculas de cada uno), que ilustran diferencias en el empaquetamiento de moléculas en estado sólido y consecuentemente diferencias en las fuerzas de dispersión de Londres.
Figura 2. Aquí se muestran ejemplos de glicerolípidos saturados e insaturados (dos moléculas de cada uno), que ilustran diferencias en el empaquetamiento de moléculas en estado sólido y consecuentemente diferencias en las fuerzas de dispersión de Londres.
Los glicerolípidos con un punto de fusión más alto que suelen ser sólidos a temperatura ambiente se denominan grasas o ceras. Los glicerolípidos con un punto de fusión más bajo que suelen ser líquidos a temperatura ambiente se denominan aceites.
Un papel importante de los glicerolípidos es en el almacenamiento de energía. Los átomos de carbono en los ácidos grasos están principalmente involucrados en los enlaces C-H, y por lo tanto pueden oxidarse y en el proceso liberar energía. En los organismos vivos, esto se lleva a cabo de manera controlada a través de enzimas. Pero también podemos quemar grasas y ceras (por ejemplo, en una vela) para obtener energía en forma de luz.
D16.5 Fosfolípidos
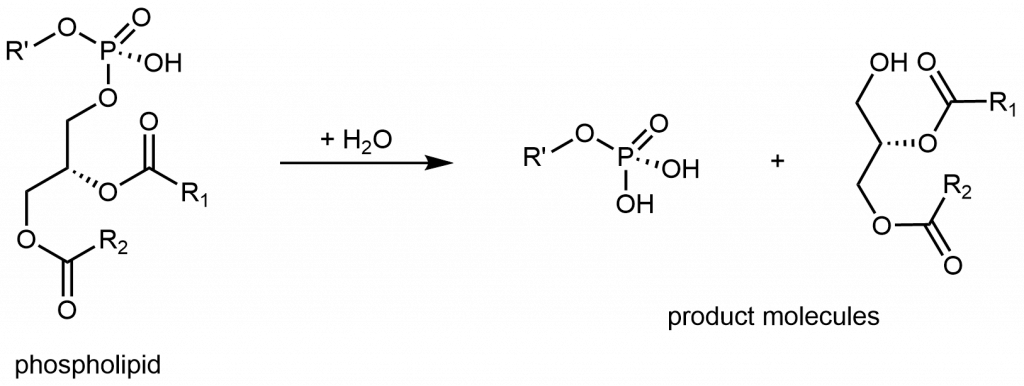
Si uno de los ácidos grasos en un glicerolípido es reemplazado por un grupo fosfato hidrofílico, el resultado es una molécula de fosfolípido.
Debajo de la estructura lineal hay una forma común y simplificada de representar una molécula de fosfolípido. El grupo fosfato, junto con los grupos éster polares, a menudo se llama la cabeza hidrofílica de la molécula (círculo rojo). Las cadenas de hidrocarburos de ácidos grasos a menudo se denominan colas hidrófobas de la molécula (líneas onduladas negras). Las moléculas con partes tanto hidrofóbicas como hidrófilas se denominan anfífilos o moléculas anfifílicas.
En un ambiente acuoso, como una célula viva, las moléculas anfifílicas se disponen espontáneamente en estructuras ordenadas. La más importante de ellas es una bicapa lipídica, dos capas de moléculas de fosfolípidos con las colas hidrófobas entremezcladas y las cabezas hidrófilas en contacto con el agua. Como muestra el siguiente diagrama, se excluye el agua del centro de la bicapa donde están las colas de ácidos grasos, mientras que los grupos de cabeza polares tienen acceso al agua. Las bicapas fosfolipídicas forman la base para las membranas celulares y todos los demás límites de orgánulos que se encuentran en las células vivas. Las membranas que se encuentran en las células también contienen muchos otros componentes, como ácidos grasos libres, otros lípidos y proteínas.
 Figura 3. Bicapa lipídica. Las áreas azules en la parte superior e inferior representan moléculas de agua. Las esferas anaranjadas representan grupos fosfato y áreas verdes las partes de glicerol de las cabezas hidrofílicas. El área gris en el centro son cadenas de hidrocarburos en las colas hidrófobas de las moléculas lipídicas. La representación muestra 128 moléculas lipídicas. Fuente: “Un campo de fuerza lipídica ámbar Tutorial: Lipid 14 Edition” de Benjamin D. Madej y Ross C. Walker, http://ambermd.org/tutorials/advance... /#Introduction.
Figura 3. Bicapa lipídica. Las áreas azules en la parte superior e inferior representan moléculas de agua. Las esferas anaranjadas representan grupos fosfato y áreas verdes las partes de glicerol de las cabezas hidrofílicas. El área gris en el centro son cadenas de hidrocarburos en las colas hidrófobas de las moléculas lipídicas. La representación muestra 128 moléculas lipídicas. Fuente: “Un campo de fuerza lipídica ámbar Tutorial: Lipid 14 Edition” de Benjamin D. Madej y Ross C. Walker, http://ambermd.org/tutorials/advance... /#Introduction.
Dado que el centro de la bicapa es no polar, solo las moléculas no polares, como O 2, CO y CO 2, pueden cruzar la bicapa. Los iones, como K +, Na + y Cl —, están completamente bloqueados por bicapas. Debido a la gran cantidad de átomos que tendrían que ser reordenados para permitir el paso de moléculas grandes, los ácidos grasos, los carbohidratos complejos, las proteínas y los ácidos nucleicos también están bloqueados por bicapas.
16.6 Proteínas, Lípidos y Ácidos Grasos
El paso de moléculas grandes o iones a través de bicapas lipídicas, puede ser controlado por moléculas de proteína que están incrustadas en una bicapa y se extienden fuera de la bicapa en ambos lados. En estas proteínas que abarcan la membrana, las cadenas laterales enfrentadas a las membranas lipídicas suelen ser hidrofóbicas; las cadenas laterales que entran en contacto con el espacio acuoso a ambos lados de la membrana pueden ser polares.

Muchas proteínas se anclan a las membranas celulares formando un enlace covalente a un ácido graso. Por ejemplo, el grupo amina (-NH 2) en el extremo N de una proteína o en una cadena lateral de lisina puede sufrir condensación con el grupo ácido carboxílico (-COOH) en un ácido graso que se encuentra en una membrana celular para formar un enlace amida. Tal enlace covalente mantiene la proteína asociada con la membrana.
Varias vitaminas necesarias para una mayor vida animal solo son solubles en lípidos. Estas vitaminas están compuestas casi en su totalidad por carbono e hidrógeno, lo que significa que son hidrofóbicas como las colas de ácidos grasos de los lípidos. Una dieta completamente carente de grasa será deficiente en estas vitaminas a menos que las vitaminas se tomen como pastillas.
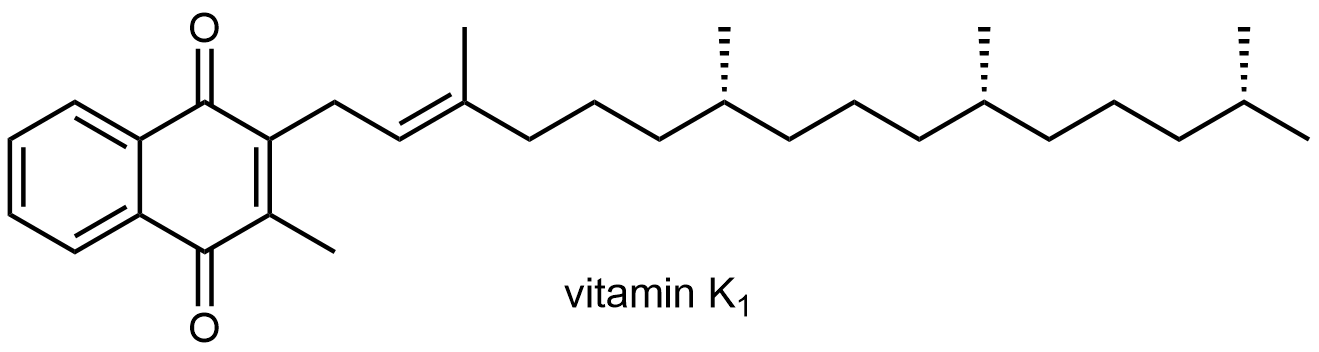
Por ejemplo, la vitamina K 1 es necesaria para la coagulación de la sangre:
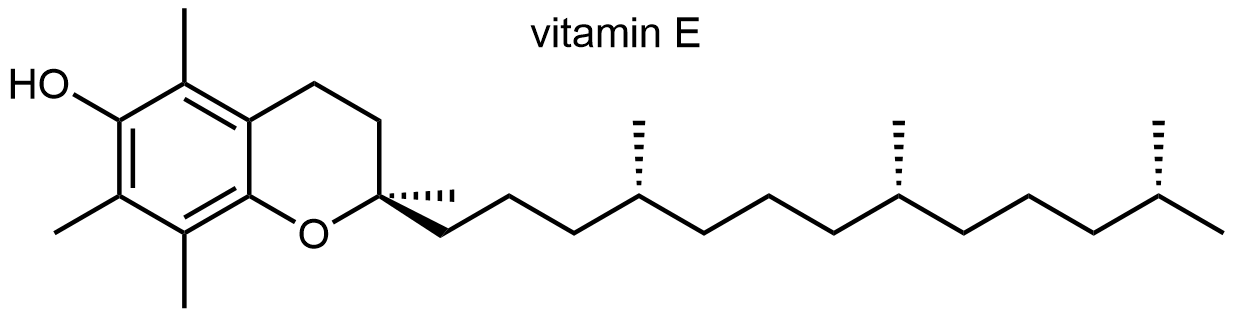
El α-tocoferol (Vitamina E) previene la oxidación inapropiada de los lípidos de membrana. La deficiencia en Vitamina E también causa infertilidad en ratas.
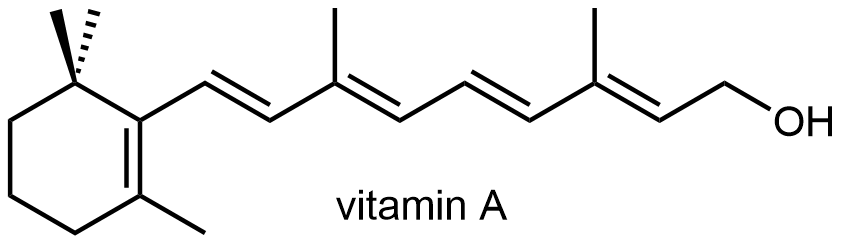
El retinol (Vitamina A) es necesario para el crecimiento juvenil normal. La deficiencia de vitamina A también conduce a la ceguera nocturna.
El colesterol lipídico de cuatro anillos forma la base para otra clase de lípidos. El colesterol es un bloque de construcción para muchas moléculas importantes utilizadas para diversos fines en organismos superiores. Se oxida parcialmente en el hígado a sales biliares, que actúan como detergentes para solubilizar los ácidos grasos en los alimentos y permitir que sean absorbidos por el organismo. Muchas hormonas, entre ellas los estrógenos y la testosterona, también se derivan del colesterol. El siguiente diagrama muestra las estructuras del colesterol, ácido glicólico (el ácido biliar más común), estradiol y testosterona (dos hormonas bien conocidas que son derivadas del colesterol). Como puede ver, las cuatro son moléculas son anfífilos, ya que tienen regiones hidrofóbicas e hidrófilas.

Los lípidos también desempeñan un papel en la señalización intracelular. Algunos estímulos, como el olor a pimientos morrones en las células receptoras olfativas, activan una enzima que descompone los fosfolípidos en moléculas más pequeñas. Estas moléculas, derivados de glicerol y ácidos grasos, luego actúan sobre otras enzimas en la célula.
Día 16 Problema de Podios Pre-Clase: Sustancias Polifluoroalquílicas
La presencia de sustancias per- y polifluoroalquilo (PFAS) en el medio ambiente es un importante problema de salud pública. Algunos sitios en Wisconsin y muchos en Michigan y otros estados han sido identificados como contaminados por estos compuestos. Se les ha denominado “químicos para siempre” porque no se degradan de forma natural. Consulte el artículo de Ciencia en este enlace para obtener más información. El artículo de Science dice que uno de los problemas de las PFAS es el hecho de que algunas de las moléculas involucradas tienen estructuras que permiten que el agua y las gotas aerotransportadas las lleven a largas distancias lejos de sus fuentes.
1. Uno de los PFAS es el ácido perfluorooctanoico, el cual se utiliza en la fabricación de polímeros de teflón. (Perfluoro significa que todos los átomos de H en una cadena de hidrocarburo han sido reemplazados por átomos de F). Escriba una estructura de Lewis para el ácido perfluorooctanoico.
2. Consideremos la estructura del ácido perfluorooctanoico y la afirmación anterior de que “algunas de las moléculas involucradas tienen estructuras que permiten que el agua y las gotitas aerotransportadas las lleven largas distancias”. Utilizar modelos de estructura molecular y fuerzas intermoleculares para construir una explicación de este hecho.
3. Otras sustancias perfluoroalquílicas tienen cadenas de átomos de carbono más largas que el ácido perfluorooctanoico. ¿En qué tipo de tejido corporal esperarías encontrar estas sustancias? Utilizar modelos de estructura molecular y fuerzas intermoleculares para responder a la pregunta y construir una explicación.
Dos días antes de la próxima sesión de toda la clase, esta pregunta de Podia se pondrá en vivo en Podia, donde podrás enviar tu respuesta.