2.1: Fijación de Carbono - Biosíntesis de Az
- Page ID
- 70240

Estrategias para la biosíntesis de glucosa
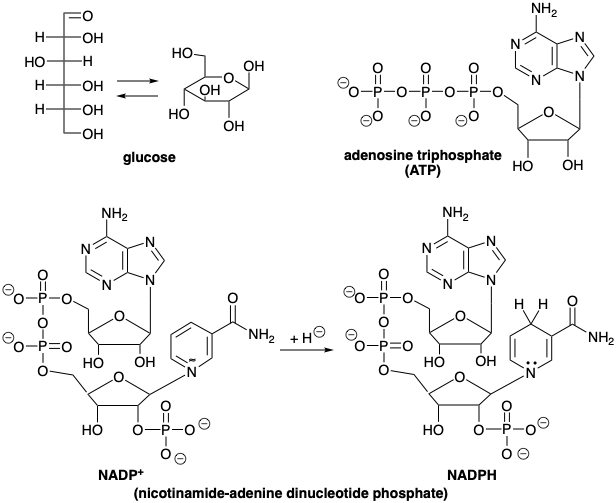
La glucosa es el material de partida orgánico definitivo a partir del cual todos los demás compuestos orgánicos de carbono pueden sintetizarse en la naturaleza. Los carbonos individuales de seis moléculas de dióxido de carbono son cosidos para formar glucosa por organismos fotosintéticos. La energía para esta reacción es proporcionada por la hidrólisis de adenosina trifosfato (ATP) para producir adenosina difosfato (ADP) y fosfato (P). Los átomos de hidrógeno son proporcionados por el derivado 1,4-dihidro (NADPH) del dinucleótido fosfato de nicotinamida-adenina (NADP +) y por protones. El NADPH es una fuente de hidruro, un “agente de transferencia de hidruro”, que es estable en el ambiente acuoso de biosíntesis, es decir, a pH fisiológico no reacciona con protones para generar hidrógeno molecular. El trifosfato ATP de alta energía es producido por una fosforilación fotoquímica de ADP para producir ATP. El agente reductor NADPH, una dihidropiridina, se produce en la misma reacción por reducción fotoquímica de NADP +. Otro producto importante de esta reacción es el oxígeno molecular que se necesita para el catabolismo oxidativo de productos naturales para proporcionar energía en forma de ATP, es decir, por fosforilación oxidativa de ADP.
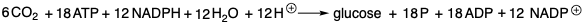
La elongación de una cadena de azúcar de cinco carbonos a una cadena de seis carbonos añadiendo una molécula del electrófilo de un carbono\(\ce{CO2}\) es una estrategia especialmente obvia para una síntesis de glucosa a partir de\(\ce{CO2}\). La adición de un nucleófilo de carbono a\(\ce{CO2}\) (f = 4) produciría un grupo carboxilo (f = 3). Esto sugiere que la primera dislocación de la diana podría ser la oxidación del grupo aldehído (f = 2) a un carboxilo. Existe un circuito consonante entre el grupo carboxilo y la funcionalidad oxígeno en la posición 3 de la subdiana de ácido glucónico resultante. Sin embargo, la expresión de la reactividad polar requerida en la posición 2 requiere conjugación como en el enol de ribulosa. Por lo tanto, el ajuste del nivel de funcionalidad en la posición 3 es una segunda dislocación lógica. La desconexión del carboxilo terminal (una condensación retro de Claisen) como tercera dislocación del objetivo sugiere ribulosa como material de partida.
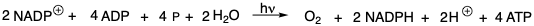
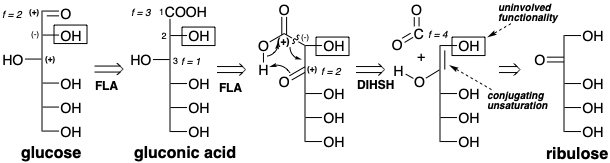
Curiosamente, la estrategia anterior no es operativa en la biosíntesis de glucosa aunque se utiliza, aunque a la inversa, para la generación de ribulosa a partir de glucosa por la vía del fosfogluconato (vide infra). Más bien, la biosíntesis de glucosa implica una estrategia diferente aunque los materiales de partida son efectivamente\(\ce{CO2}\) y ribulosa. La glucosa tiene una amplia funcionalidad para facilitar su construcción mediante estrategias conectivas C-C que implican la generación de cualquiera de sus enlaces C-C por reacciones polares. Por ejemplo, el enlace 3,4-podría generarse por reacción de un sintón nucleofílico correspondiente a los carbonos 1-3 con un fragmento electrófilo correspondiente a los carbonos 4-6. Esto sugiere una primera dislocación de la diana que implica el ajuste del nivel de funcionalidad para permitir la conjugación del oxígeno C-2 con la posición 3 para facilitar la generación de reactividad nucleofílica en C-3. Al acoplar la oxidación en la posición 2 con reducción en la posición 1, la primera dislocación es simplemente una isomerización de la glucosa diana, una aldosa, a fructosa, una cetosa. La desconexión polar del subblanco (una condensación retro aldólica) en la segunda dislocación tiene la importante consecuencia de dividir el objetivo en dos fragmentos funcionalizados de manera similar con esqueletos de carbono idénticos. Tales dislocaciones potencialmente revelan simetría latente que se define como la posibilidad de derivar dos mitades de un objetivo a partir de un material de partida común. Los precursores generados en la segunda dislocación, la dihidroxiacetona y el gliceraldehído se interconvierten fácilmente por isomerización a través de un intermedio de enodiol.
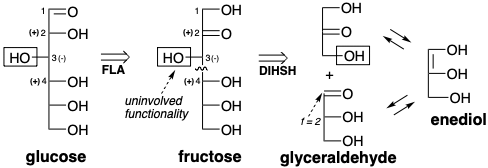
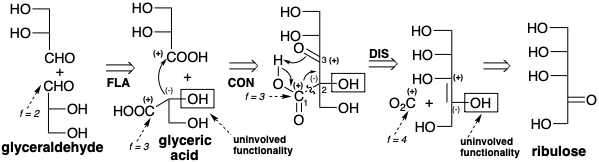
Una vez más, observamos que la\(\ce{CO2}\) incorporación de a un precursor por un proceso polar generará un grupo carboxilo. Esto sugiere un ácido carboxílico, ácido glicérico, que puede servir como precursor común tanto para la dihidroxiacetona como para el gliceraldehído. Reterosintéticamente esto implica dislocación de dihidroxiacetona a gliceraldehído (una isomerización) y dislocación de ambas moléculas de gliceraldehído al mismo precursor ácido (oxidación). La conexión polar de dos moléculas de ácido glicérico (una condensación de Claisen) sugiere una subdiana β-cetoácida de la que se puede desconectar un grupo carboxilo en la última dislocación de la diana que conduce a los precursores\(\ce{CO2}\) y ribulosa.
Biosíntesis de Glucosa
De hecho, la biosíntesis real implica la carboxilación de este azúcar de cinco carbonos, aunque en forma de derivado de bisfosfato, ribulosa-1,5-bisfosfato (RuBP). Una estrategia para la biosíntesis del rUBP subobjetivo podría proceder de manera gradual agregando carbono\(\ce{CO2}\) derivado de uno a la vez a una cadena de carbono en crecimiento. Tal proceso podría requerir una enzima diferente para catalizar la fijación de cada molécula\(\ce{CO2}\) por adición a diferentes subdianas. Sin embargo, se adopta en la naturaleza una estrategia mucho más ingeniosa. La fijación de carbono ocurre solo por la reacción de\(\ce{CO2}\) con RuBP. Por lo tanto, solo se requiere una sola enzima para catalizar el proceso. Seis moléculas de\(\ce{CO2}\) se combinan con seis moléculas del derivado de azúcar de cinco carbonos RubP para producir doce moléculas de ácido glicérico, dos de las cuales se utilizan para generar glucosa por la estrategia anterior. Los treinta carbonos de las diez moléculas de ácido glicérico restantes se reagrupan para regenerar seis RUBP de cinco carbonos. Así, RuBP también funciona como un catalizador para la\(\ce{CO2}\) bioconversión de glucosa.
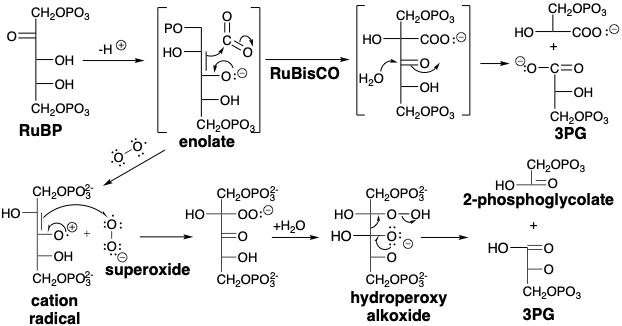
La formación fotosintética de glucosa (en realidad en “las reacciones oscuras de la fotosíntesis”) implica una intrincada serie de reacciones conocidas como el ciclo Calvino. En los esquemas de reacción adjuntos, P se usa para representar un éster de fosfato [P = —\(\ce{PO3^2-}\)]. El ciclo de fijación de carbono comienza con la carboxilación de RuBP (ver más abajo), una reacción que es catalizada por la ribulosa bisfosfato carboxilasa oxidasa (RuBisCO) que es probablemente la proteína más abundante en la Tierra. Así, la carboxilación del enol de RubP conduce a un supuesto intermedio b-ceto-ácido que se escinde fácilmente por el agua en una condensación retro de Claisen para dar dos moléculas de ácido 3-fosfoglicérico (3PG). Dada la importancia de esta química para la biosíntesis de moléculas orgánicas y el éxito de las formas de vida basadas en carbono, es de destacar que la catálisis de RubiSco de esta reacción es apenas viable. A niveles ambientales de dióxido de carbono y oxígeno, el catalizador consume solo unas pocas moléculas de\(\ce{CO2}\) por segundo en contraste con muchas enzimas que procesan miles o decenas de miles de moléculas de sustrato por segundo. Además, RubisCO cataliza otra reacción que compite con la carboxilación, la escisión oxidativa de RubP a 3-PG más 2-fosfoglicolato. Esta escisión oxidativa implica presumiblemente la transferencia de electrones desde el intermedio de enolato al oxígeno para producir superóxido y un radical catiónico. La formación de enlaces entre estos dos radicales genera entonces un hidroperoxi alcóxido. La fragmentación de este intermedio es impulsada por la generación exotérmica de dos grupos carbonilo. La relación de carboxilación versus escisión oxidativa es solo de aproximadamente 4 a 1 y es aún menos favorable a temperaturas más altas. Así, las plantas en ambientes de alto calor almacenan dióxido de carbono durante las horas calurosas de sol intenso, y generan glucosa en las horas más frías en ausencia de sol y su calor abrasador.
La biosíntesis de todos los azúcares, incluyendo la glucosa y la regeneración de RuBP, utilizan 3PG como el materisl común y único de partida. El 3PG se reduce primero a un aldehído, gliceraldehído-3-fosfato (G3P), por NADPH. El ATP facilita la reducción al convertir el carboxilo en un derivado más electrófilo, un anhídrido carbólico-fosfórico. El resto de las reacciones del ciclo Calvino redistribuyen los treinta y seis átomos de carbono de doce G3Ps para producir una molécula de glucosa (seis átomos de carbono) y regenerar seis moléculas de RubP (treinta átomos de carbono). El ciclo se resumirá en el Cuadro 1 a continuación.
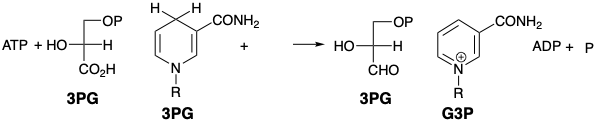
La aldosa G3P se transforma en la cetosa dihidroxiacetona fosfato (DHAP) bajo la influencia de la enzima isomerasa. El esqueleto de seis átomos de carbono de la glucosa se ensambla luego mediante una condensación aldólica de G3P con DHAP. El producto inicial, bisfosfato de fructosa (FBP), se hidroliza (a F6P), se isomeriza (a G6P) que se hidroliza adicionalmente para producir glucosa.
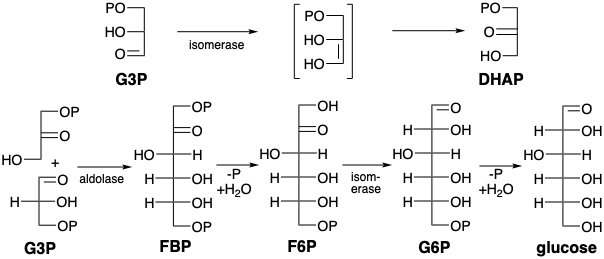
Regeneración de Bifosfato de Ribulosa
El análisis retrosintético revela que una síntesis polar de Ru5P a partir de G3P requiere un sintón umpoled, el 2-hidroxiacetil carbanión. 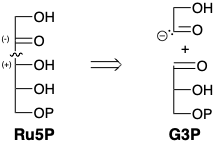 Una condición límite, las condiciones de reacción acuosa de biosíntesis limitan la elección de equivalentes sintéticos para este sintón. Es instructivo considerar que se requiere un sintón 1 similar en la condensación de benzoína, un acoplamiento polar catalizado por iones cianuro de dos grupos carbonilo benzaldehído electrófilos que se pueden lograr en solución acuosa. El cianuro invierte la reactividad polar de un carbono carbonílico de benzaldehído por adición nucleofílica seguida de una transferencia de protones que genera el carbanión 2, un equivalente sintético del sintón 1. La generación de aniones en el carbono carbonilo anterior se ve favorecida por la conjugación con el nitrilo en 2. Es la bifilicidad del cianuro la que es la base de su capacidad para invertir la reactividad polar de un carbono de aldehído carbonilo. La condensación de 2 con una segunda molécula de benzaldehído proporciona el alcóxido 3 que proporciona el alcóxido 4 por transferencia de protones. La expulsión de cianuro de 4 luego regenera el catalizador y produce benjuí. Las reacciones análogas catalizadas por cianuro de otros aldehídos se denominan genéricamente condensaciones de benzoína. Todos los pasos de la condensación del benjuí son reversibles. Por lo tanto, el sintón 2 umpolado no solo puede generarse por reacción de cianuro con benzaldehído, sino también por condensación retro de benzoína de benzoína.
Una condición límite, las condiciones de reacción acuosa de biosíntesis limitan la elección de equivalentes sintéticos para este sintón. Es instructivo considerar que se requiere un sintón 1 similar en la condensación de benzoína, un acoplamiento polar catalizado por iones cianuro de dos grupos carbonilo benzaldehído electrófilos que se pueden lograr en solución acuosa. El cianuro invierte la reactividad polar de un carbono carbonílico de benzaldehído por adición nucleofílica seguida de una transferencia de protones que genera el carbanión 2, un equivalente sintético del sintón 1. La generación de aniones en el carbono carbonilo anterior se ve favorecida por la conjugación con el nitrilo en 2. Es la bifilicidad del cianuro la que es la base de su capacidad para invertir la reactividad polar de un carbono de aldehído carbonilo. La condensación de 2 con una segunda molécula de benzaldehído proporciona el alcóxido 3 que proporciona el alcóxido 4 por transferencia de protones. La expulsión de cianuro de 4 luego regenera el catalizador y produce benjuí. Las reacciones análogas catalizadas por cianuro de otros aldehídos se denominan genéricamente condensaciones de benzoína. Todos los pasos de la condensación del benjuí son reversibles. Por lo tanto, el sintón 2 umpolado no solo puede generarse por reacción de cianuro con benzaldehído, sino también por condensación retro de benzoína de benzoína.
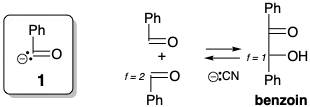
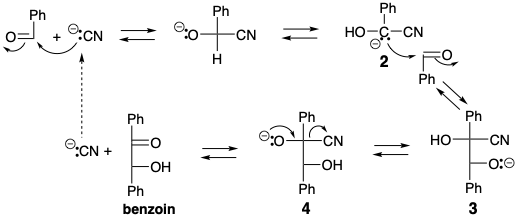
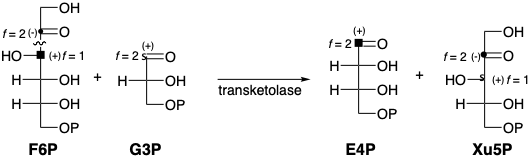
Una condensación retro de benzoína similar de fructosa 6-fosfato (F6P), un intermedio generado en la biosíntesis de glucosa a partir de gliceraldehído 3-fosfato (G3P), podría proporcionar un equivalente sintético del 2-hidroxiacetil carbanión requerido para la regeneración biosintética de RuBP a partir de G3P. De hecho, la biosíntesis de RubP a partir de G3P implica la transferencia de un grupo 2-hidroxiacetilo de F6P a G3P que es promovido por la enzima transcetolasa.
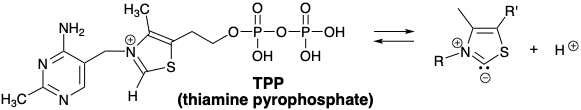
Huelga decir que el cianuro no es el cocatalizador que enmascara la reactividad electrofílica habitual de un grupo carbonilo e imparte reactividad nucleofílica a él en la naturaleza. El equivalente biológico del ion cianuro es un carbanión generado por la desprotonación del anillo de tiazol en pirofosfato de tiamina (TPP).
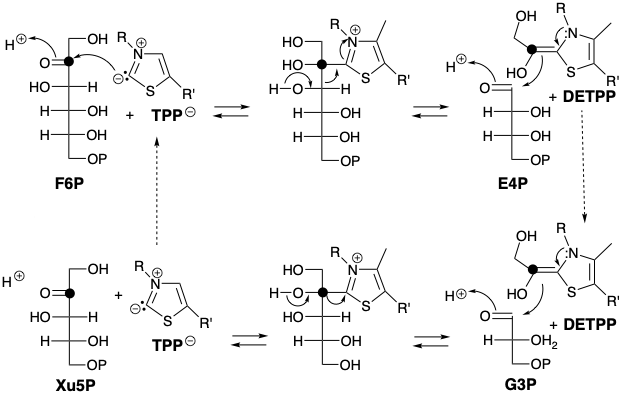
El nucleófilo de carbanión tiamina se condensa con el carbono carbonílico electrófilo de una cetosa (por ejemplo, F6P) para producir un derivado de 2-hidroxiiminio. Este último experimenta fácilmente una reacción de tipo retro aldol que conduce a una aldosa, por ejemplo, eritrosa 4-fosfato (E4P), que tiene dos carbonos menos que la cetosa original. El equivalente nucleofílico de 2-hidroxiacetilo resultante, pirofosfato de 2- (1,2-dihidroxietilideno) tiamina (DETPP) puede condensarse con una aldosa diferente, por ejemplo G3P, para regenerar TPP y una cetosa, por ejemplo, xilosa 5-fosfato (Xu5P) que tiene dos carbonos más que la aldosa.
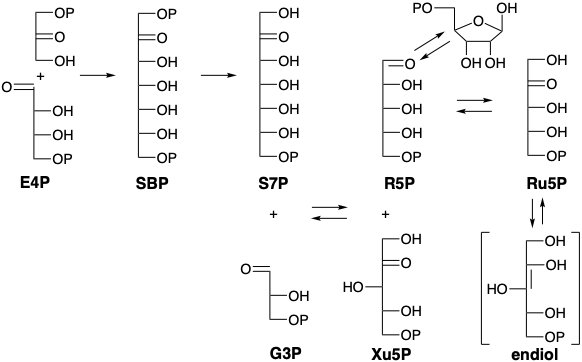
Otras reacciones en el ciclo de Calvin son la condensación promovida por aldolasa de E4P con DHAP para producir sedoheptulosa bisfosfato (SBP), hidrólisis de este último al monofosfato (S7P), transcetolasa promovió la transferencia de hidroxiacilo de S7P a G3P para dar Xu5P más ribosa 5-fosfato (R5P), isomerización del último a ribulosa 5-fosfato (Ru5P), epimerización de Xu5P para dar Ru5P y fosforilación de este último para regenerar RubP. El Ciclo Calvino se resume en el gráfico 1.
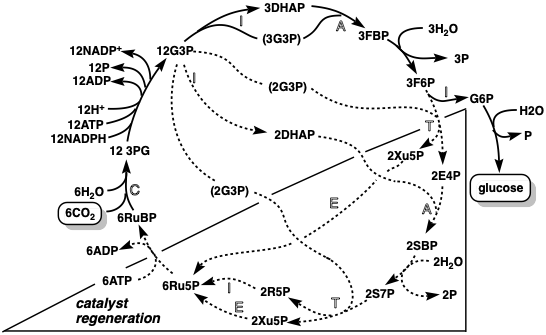
Resumen de Biosynthetic Carbon Fijation
(1) Solo hay una reacción que convierte el dióxido de carbono en materiales de partida orgánicos: la generación de dos 3PG a partir de RuBP y\(\ce{CO2}\). Este es el paso #1 en la biosíntesis de todos los productos naturales. (2) RubP sirve como catalizador en un ciclo que\(\ce{CO2}\) convierte seis en una molécula de glucosa. (3) El rUBp consumido en el paso #1 es regenerado por una serie de reacciones, que reagrupan los átomos de diez moléculas del azúcar de tres carbonos G3P en seis moléculas del RuBP de cinco carbonos. (4) Toda la formación de enlaces C-C y escisión implica condensaciones (aldol, claisen, benzoína) que son fácilmente reversibles. Además, la aldolasa, la enzima que cataliza la formación de glucosa a partir de azúcares de tres carbonos, también cataliza su regeneración. Como veremos, nuevas transformaciones biosintéticas de la glucosa en ácidos grasos, terpenos o policétidos comienzan con la escisión de la glucosa (glucólisis) por esta reacción retro aldólica.
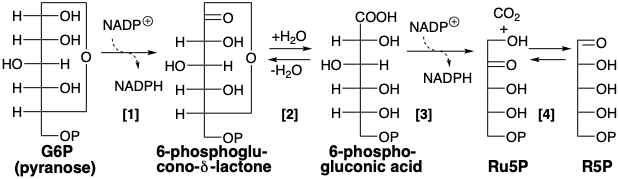
Ribulosa de Glucosa
Antes de proceder con un examen de estrategias para la síntesis de los materiales de partida de glucosa, primero volvamos a la estrategia simple esbozada anteriormente que no se utiliza para la biosíntesis de glucosa. Por lo tanto, en lugar de servir como una ruta sintética a la glucosa de RuBP, esta estrategia a la inversa se usa en la naturaleza para producir azúcares de cinco carbonos a partir de glucosa. Esta vía para la degradación de la glucosa es importante para la biosíntesis porque genera pentosas para la síntesis de ácidos nucleicos. La vía comienza con la oxidación de la glucosa 6-fosfato a 6-fosfogluconato. Se conoce como la vía del fosfogluconato, la derivación de hexosa monofosfato o la vía de pentosa fosfato. Muchas de las enzimas y reacciones de esta vía también están involucradas en la biosíntesis de glucosa a partir de\(\ce{CO2}\) reacciones oscuras de la fotosíntesis. La vía fosfogluconato produce 2NADPH + CO 2 + R5P a partir de glucosa y 2NADP +. Se requieren cuatro enzimas: [1] 6-glucosa fosfato deshidrogenasa, [2] lactonasa, [3] 6-fosfogluconato deshidrogenasa y [4] fosfopentosa isomerasa.