2.3: Acetil CoA - un material de partida derivado de azúcar
- Page ID
- 70261

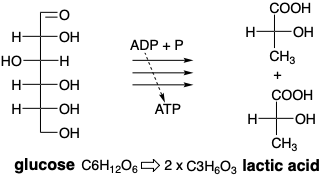
Todos los átomos de carbono de la glucosa están unidos al oxígeno. En contraste, muchos productos naturales complejos están mucho menos oxigenados. Por ejemplo, los ácidos grasos (Capítulo 3) son cadenas lineales largas de a menudo más de una docena de átomos de carbono sin oxígeno en absoluto excepto por un carbono terminal que está completamente oxidado a un ácido carboxílico. El catabolismo de la glucosa (descomposición) puede proceder anaeróbicamente (no requiere oxígeno) produciendo pequeñas moléculas biosintéticamente útiles en las que algunos átomos de carbono son menos y algunos más altamente oxidados. No se produce oxidación neta. Los átomos de oxígeno de la glucosa son simplemente reagrupados. El producto final del proceso es el ácido láctico, una molécula que es rica en oxígeno en un extremo y pobre en oxígeno en el otro. Quizás lo más importante para los organismos vivos, el catabolismo aneróbico de la glucosa también genera energía química en forma de ATP que puede ser utilizada, entre otras cosas, para potenciar las contracciones musculares.
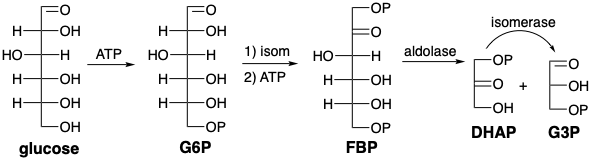
La glucólisis comienza con la fosforilación de la glucosa seguida de isomerización a fructosa 6-fosfato (F6P) que luego se fosforila posteriormente. El bisfosfato de fructosa se escinde bajo la influencia de la aldolasa en una reacción retero-aldólica para producir DHAP y G3P. La isomerización de DHAP produce entonces una segunda molécula de G3P.
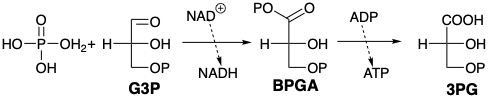
La transformación final de G3P en ácido láctico comienza con la eliminación del hidruro de la porción aldehído de las moléculas (oxidación) por el dinucleótido de nicotinamida-adenina (NAD\({}^\oplus\)). La reacción es catalizada por la enzima gliceraldehído 3-fosfato deshidrogenasa. La enzima se une a G3P como un hemitioacetal que transfiere fácilmente hidruro a un NAD unido a enzima\({}^\oplus\). El producto, un tioéster reactivo de ácido fosfoglicérico, acila fácilmente el fosfato para producir ácido bifosfoglicérico (BPGA), un anhídrido mixto reactivo. A continuación, BPGA fosforila ADP. De ahí que la energía química generada en esta oxidación se almacene en la energía de enlace fosfato del ATP.
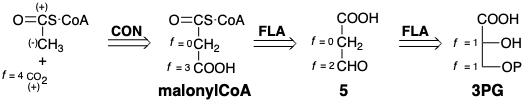
Para la biosíntesis de ácidos grasos (Capítulo 3), terpenos (Capítulo 4), o policétidos (capítulo 5), el ácido fosfoglicérico se desmonta adicionalmente para formar una molécula de\(\ce{CO2}\) y un tioéster de ácido acético con un tiol estructuralmente complejo, la coenzima A. Este tioéster, denominado acetil CoA, tiene un carbono que se reduce por completo, un grupo metilo. Dado que el acetil metilo es potencialmente nucleófilo y el carbono de\(\ce{CO2}\) es electrófilo, una estrategia obvia para la biosíntesis de acetil CoA y\(\ce{CO2}\) de 3PG utiliza malonil CoA como penúltima diana y explota la escisión polar de un enlace C-C durante la descarboxilación. El ajuste del nivel de funcionalidad de esta subdiana sugiere un precursor 5 que tiene el mismo nivel de funcionalidad general que el material de partida deseado 3PG. La conversión de 3PG a 5 se pudo lograr mediante la eliminación del agua seguida de la readición e hidrólisis del fosfato. Curiosamente, esta estrategia no se utiliza para la biosíntesis de acetil CoA aunque se utiliza la primera dislocación, aunque a la inversa, para generar malonil CoA a partir de acetil CoA (vide infra).
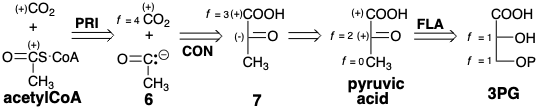
Una estrategia alternativa para la biosíntesis de acetil CoA a\(\ce{CO2}\) partir de 3PG implica la escisión del carbono acilo incipiente. Pero esto requiere umpolung de la electrofilicidad normal del carbono carbonilo en acetil CoA. Es decir, la escisión polar tendría que generar un sintón acil-nucleófilo umpolado 6 a partir de un sintón umpolado 7 de ácido pirúvico. La generación de ácido pirúvico a partir de 3PG solo requiere redistribución de la funcionalidad. Esta es la estrategia biosintética real para acetil CoA.
La biosíntesis de acetil CoA utiliza muchas de las reacciones involucradas en el catabolismo aneróbico de la glucosa. Por lo tanto, retomemos nuestra discusión sobre la biosíntesis del ácido láctico a partir de la glucosa (ver arriba). El ácido 3-fosfoglicérico (3PG) que resulta de la oxidación de G3P (ver arriba) sufre una transferencia del grupo fosforilo del 3- al 2-hidroxilo y posterior deshidratación a fosfoenolpiruvato (PEP). Este éster enólico es rico en energía ya que su hidrólisis genera un enlace C=O relativamente fuerte a expensas de un enlace C=C relativamente débil. PEP fosforila fácilmente ADP liberando ácido pirúvico. El átomo de carbono del grupo cetona carbonilo de este α-cetoácido es muy electrófilo y acepta fácilmente hidruro de NADH bajo catálisis de la enzima lactato deshidrogenasa.
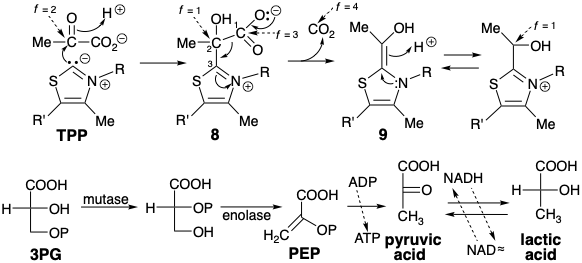
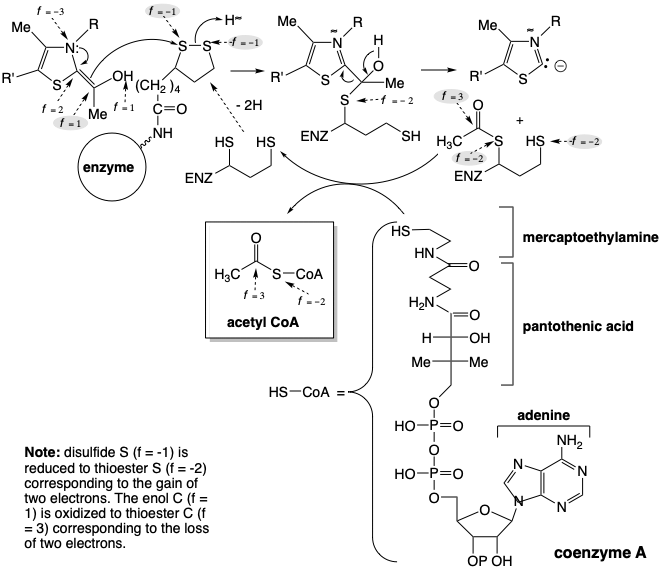
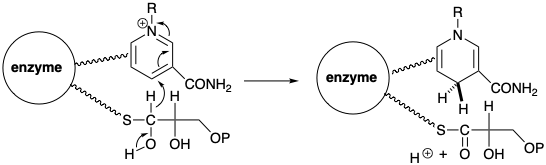
La biosíntesis de acetil CoA a partir de glucosa implica la descarboxilación del ácido pirúvico que se regenera por deshidrogenación del ácido láctico. Para permitir una reacción de descarboxilación polar, el ácido pirúvico se forma por el derivado de polionacarbonilintoamoro-carboxilato de ácido pirúvico. Al igual que en la reacción de transcetolasa, la reactividad polar de este grupo carbonilo es invertida temporalmente por el resto carbanión tiazol bifílico del pirofosfato de tiamina (TPP). Así, el carbono del anillo de tiazol nucleófilo del TPP se condensa con el carbono carbonílico altamente electrófilo del ácido pirúvico para dar un aducto 8 que se asemeja a un β-cetoácido. Esto se somete a descarboxilación por escisión retro de Claisen para suministrar hidroxietilideno TPP (9). El nivel de funcionalidad del carbono carboxilo incipiente es solo f = 1 en 9. Por lo tanto, se requiere oxidación de 9 para producir un nivel de funcionalidad acetilo (i. e. f = 3).
Esta oxidación se logra mediante un proceso polar que reduce concomitantemente un disulfuro a un ditiol. Así, 9 transfiere un grupo acetilo al disulfuro de un residuo de ácido lipoico unido a la enzima dihidrolipoil transacilasa. El grupo acetilo se transfiere luego del tiol unido a la enzima al grupo tiol de una coenzima (CoA) para dar acetil CoA.