
28.9A: Vinculación genética y distancias
- Page ID
- 128948
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Los genes vinculados pueden desvincularse durante la recombinación; la probabilidad de que los genes se separen depende de su distancia entre sí.
Objetivos de aprendizaje
- Discutir cómo los genes vinculados se pueden heredar por separado
Puntos Clave
- Los alelos posicionados en el mismo cromosoma no siempre se heredan juntos porque durante la meiosis los genes enlazados pueden desvincularse.
- Frans Janssen sugirió que los cromosomas se desvinculan durante la recombinación homóloga, un proceso en el que los cromosomas homólogos intercambian segmentos de ADN.
- Alfred Sturtevant planteó la hipótesis de que los alelos que estaban más cerca en un gen tenían más probabilidades de heredarse juntos en lugar de alelos que estaban más separados y utilizó mediciones de recombinación entre genes para crear el primer mapa genético.
- Cuando los genes están perfectamente enlazados, tienen una frecuencia de recombinación de 0.
- Cuando los genes están desvinculados, tienen una frecuencia de recombinación de 0.5, lo que significa que el 50 por ciento de las crías son recombinantes y el otro 50 por ciento son de tipo parental.
Términos Clave
- recombinación homóloga: un tipo de recombinación genética en la que las secuencias de nucleótidos se intercambian entre dos moléculas de ADN similares o idénticas
- vinculación: la propiedad de los genes de ser heredados juntos
- sinapsis: la asociación de cromosomas maternos y paternos homólogos durante la parte inicial de la meiosis
Vinculación genética y distancias
El trabajo de Mendel sugirió que los rasgos se heredan independientemente unos de otros. Morgan identificó una relación 1:1 entre un rasgo segregante y el cromosoma X, lo que sugiere que la segregación aleatoria de cromosomas fue la base física del modelo de Mendel. Esto también demostró que los genes vinculados interrumpen los resultados predichos de Mendel. El hecho de que cada cromosoma pueda portar muchos genes vinculados explica cómo los individuos pueden tener muchos más rasgos de los que tienen cromosomas. Sin embargo, las observaciones de investigadores en el laboratorio de Morgan sugirieron que los alelos posicionados en el mismo cromosoma no siempre se heredaban juntos. Durante la meiosis, los genes vinculados de alguna manera se desvincularon.
Recombinación homóloga
En 1909, Frans Janssen observó chiasmata (el punto en el que las cromátidas están en contacto entre sí y pueden intercambiar segmentos) antes de la primera división de la meiosis. Sugirió que los alelos se desvinculan cuando los cromosomas intercambian físicamente segmentos. A medida que los cromosomas se condensaban y apareaban con sus homólogos, parecían interactuar en distintos puntos. Janssen sugirió que estos puntos correspondían a regiones en las que se intercambiaban segmentos cromosómicos. Ahora se sabe que el emparejamiento y la interacción entre cromosomas homólogos, conocidos como sinapsis, hace más que simplemente organizar los homólogos para la migración para separar las células hijas. Cuando se sinapsan, los cromosomas homólogos experimentan intercambios físicos recíprocos de ADN en sus brazos en un proceso llamado recombinación homóloga, o más simplemente, “cruzando”.
Mapas Genéticos
En 1913, Alfred Sturtevant, estudiante del laboratorio de Morgan, creó el primer “mapa cromosómico”, una representación lineal del orden de los genes y la distancia relativa en un cromosoma.Para construir un mapa cromosómico, Sturtevant asumió que los genes se ordenaban en serie en cromosomas filiformes. También asumió que la incidencia de recombinación entre dos cromosomas homólogos podría ocurrir con igual probabilidad en cualquier lugar a lo largo de la longitud del cromosoma. Operando bajo estos supuestos, Sturtevant planteó la hipótesis de que los alelos que estaban muy separados en un cromosoma tenían más probabilidades de disociarse durante la meiosis simplemente porque había una región más grande sobre la cual podría ocurrir la recombinación. Por el contrario, los alelos que estaban cerca uno del otro en el cromosoma probablemente se heredaran juntos. El número promedio de cruces entre dos alelos, o su frecuencia de recombinación, se correlacionó con su distancia genética entre sí, en relación con las ubicaciones de otros genes en ese cromosoma. Sturtevant dividió su mapa genético en unidades cartográficas, o centimorgans (cM), en las que una frecuencia de recombinación de 0.01 corresponde a 1 cM.
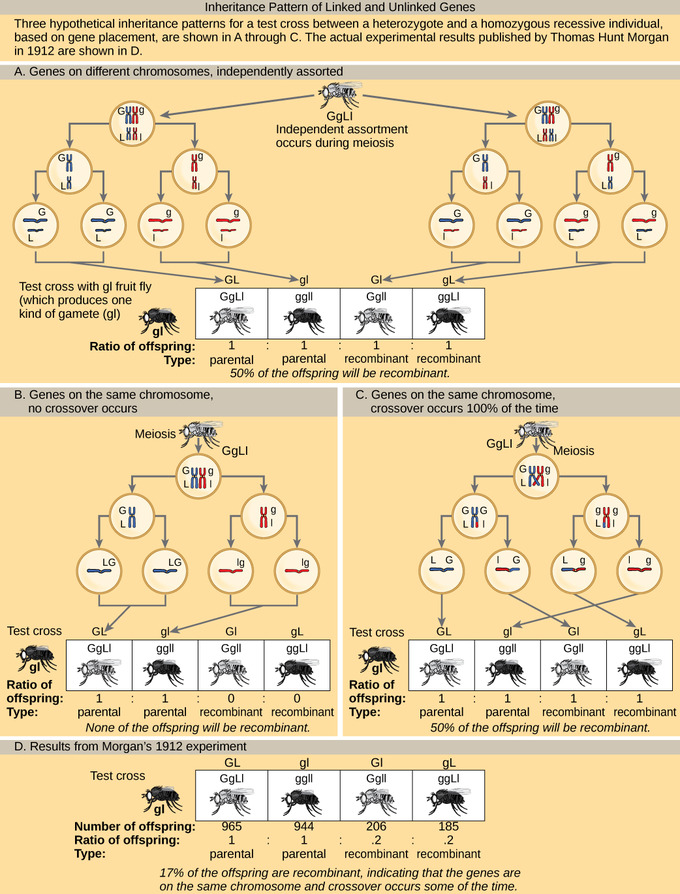
Patrones de Herencia de Genes Desenlazados y Vinculados: En (a), dos genes se localizan en diferentes cromosomas por lo que ocurre un surtido independiente durante la meiosis. Las crías tienen las mismas posibilidades de ser del tipo parental (heredar la misma combinación de rasgos que los padres) o de un tipo no parental (heredar una combinación de rasgos diferente a la de los padres). En (b), dos genes están muy unidos entre sí en un mismo cromosoma de manera que no se produce ningún cruce entre ellos. Los genes son, por lo tanto, siempre heredados juntos y todos los descendientes son del tipo parental. En (c), dos genes están muy separados en el cromosoma de tal manera que el cruzamiento ocurre durante cada evento meiótico. La frecuencia de recombinación será la misma que si los genes estuvieran en cromosomas separados. d) La frecuencia real de recombinación de la longitud del ala de la mosca de la fruta y el color corporal que Thomas Morgan observó en 1912 fue del 17 por ciento. Una frecuencia de cruce entre 0 por ciento y 50 por ciento indica que los genes están en el mismo cromosoma y el cruce ocurre algunas veces.
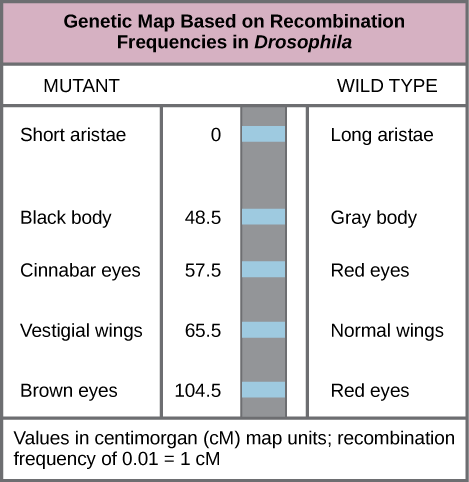
Mapas genéticos: Este mapa genético ordena los genes de Drosophila sobre la base de la frecuencia de recombinación.
Al representar alelos en un mapa lineal, Sturtevant sugirió que los genes pueden variar desde estar perfectamente unidos (frecuencia de recombinación = 0) hasta estar perfectamente desenlazados (frecuencia de recombinación = 0.5) cuando los genes están en diferentes cromosomas o los genes están separados muy lejos en el mismo cromosoma. Los genes perfectamente desenlazados corresponden a las frecuencias predichas por Mendel para agruparse independientemente en un cruce dihíbrido. Una frecuencia de recombinación de 0.5 indica que el 50 por ciento de las crías son recombinantes y el otro 50 por ciento son de tipo parental. Es decir, cada tipo de combinación de alelos se representa con igual frecuencia. Esto permitió a Sturtevant calcular distancias entre varios genes en un mismo cromosoma.