
12.4: El potencial de acción
- Page ID
- 123050

Objetivos de aprendizaje
- Describir los componentes de la membrana que establecen el potencial de la membrana en reposo
- Describir los cambios que ocurren en la membrana que resultan en el potencial de acción
Las funciones del sistema nervioso —sensación, integración y respuesta— dependen de las funciones de las neuronas subyacentes a estas vías. Para entender cómo las neuronas son capaces de comunicarse, es necesario describir el papel de una membrana excitable en la generación de estas señales. La base de esta comunicación es el potencial de acción, lo que demuestra cómo los cambios en la membrana pueden constituir una señal. Mirar la forma en que funcionan estas señales en circunstancias más variables implica una mirada a los potenciales graduados, los cuales serán cubiertos en la siguiente sección.
Membranas celulares eléctricamente activas
La mayoría de las células del cuerpo hacen uso de partículas cargadas, iones, para acumular una carga a través de la membrana celular. Anteriormente, se demostró que esto era parte de cómo funcionan las células musculares. Para que los músculos esqueléticos se contraigan, basado en el acoplamiento excitación-contracción, requiere la entrada de una neurona. Ambas células hacen uso de la membrana celular para regular el movimiento iónico entre el fluido extracelular y el citosol.
Como aprendiste en el capítulo sobre células, la membrana celular es la principal responsable de regular lo que puede atravesar la membrana y lo que permanece en un solo lado. La membrana celular es una bicapa de fosfolípidos, por lo que solo las sustancias que pueden pasar directamente a través del núcleo hidrófobo pueden difundirse sin ayuda. Las partículas cargadas, que son hidrófilas por definición, no pueden pasar a través de la membrana celular sin ayuda (Figura\(\PageIndex{1}\)). Las proteínas transmembrana, específicamente las proteínas de canal, lo hacen posible. Varios canales de transporte pasivo, así como bombas de transporte activo, son necesarios para generar un potencial transmembrana y un potencial de acción. De especial interés es la proteína portadora conocida como la bomba de sodio/potasio que mueve los iones de sodio (Na +) fuera de una célula y los iones de potasio (K +) a una célula, regulando así la concentración de iones en ambos lados de la membrana celular.
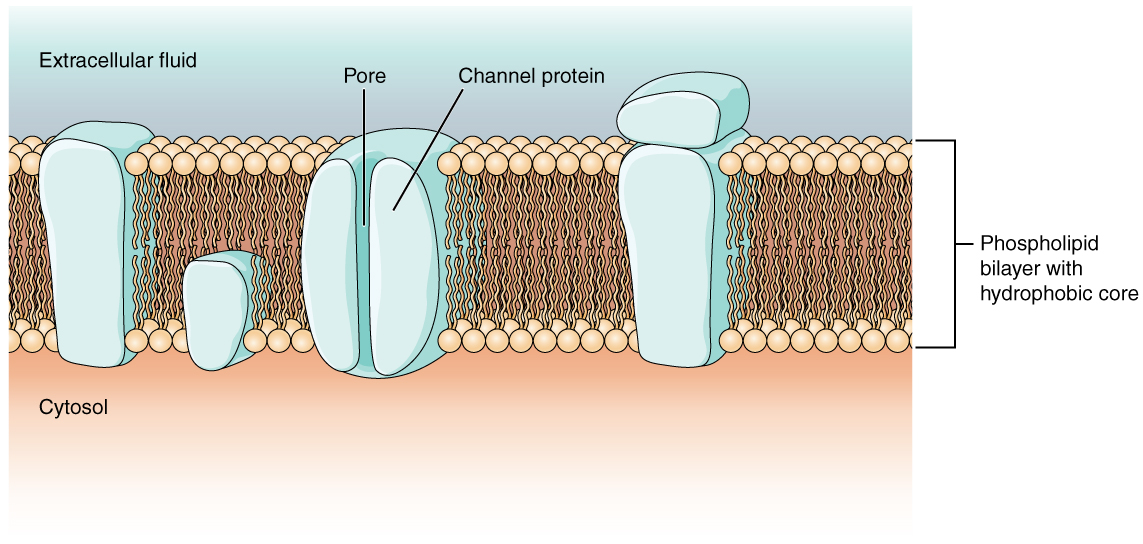
La bomba de sodio/potasio requiere energía en forma de trifosfato de adenosina (ATP), por lo que también se le conoce como ATPasa. Como se explicó en el capítulo celular, la concentración de Na + es mayor fuera de la célula que dentro, y la concentración de K + es mayor dentro de la célula que fuera. Eso significa que esta bomba está moviendo los iones contra los gradientes de concentración para sodio y potasio, razón por la cual requiere energía. De hecho, la bomba básicamente mantiene esos gradientes de concentración.
Los canales iónicos son poros que permiten que partículas cargadas específicas crucen la membrana en respuesta a un gradiente de concentración existente. Las proteínas son capaces de atravesar la membrana celular, incluyendo su núcleo hidrófobo, y pueden interactuar con la carga de iones debido a las variadas propiedades de los aminoácidos que se encuentran dentro de dominios o regiones específicos del canal proteico. Los aminoácidos hidrófobos se encuentran en los dominios que se colocan a las colas de hidrocarburos de los fosfolípidos. Los aminoácidos hidrofílicos están expuestos a los ambientes fluidos del fluido extracelular y el citosol. Adicionalmente, los iones interactuarán con los aminoácidos hidrófilos, los cuales serán selectivos para la carga del ion. Los canales para cationes (iones positivos) tendrán cadenas laterales cargadas negativamente en el poro. Los canales para aniones (iones negativos) tendrán cadenas laterales cargadas positivamente en el poro. Esto se llama exclusión electroquímica, lo que significa que el poro del canal es específico de carga.
Los canales iónicos también se pueden especificar por el diámetro del poro. La distancia entre los aminoácidos será específica para el diámetro del ion cuando se disocie de las moléculas de agua que lo rodean. Debido a las moléculas de agua circundantes, los poros más grandes no son ideales para iones más pequeños porque las moléculas de agua interactuarán, por enlaces de hidrógeno, más fácilmente que las cadenas laterales de los aminoácidos. Esto se llama exclusión por tamaño. Algunos canales iónicos son selectivos para la carga pero no necesariamente para el tamaño, y por lo tanto se denominan canales inespecíficos. Estos canales inespecíficos permiten que los cationes, particularmente Na +, K + y Ca 2+, crucen la membrana, pero excluyen aniones.
Los canales iónicos no siempre permiten que los iones se difundan libremente a través de la membrana. Algunos son abiertos por ciertos eventos, es decir, los canales están cerrados. Entonces otra forma en que los canales pueden ser categorizados es en base a cómo están cerrados. Aunque estas clases de canales iónicos se encuentran principalmente en las células del tejido nervioso o muscular, también se pueden encontrar en las células de los tejidos epiteliales y conectivos.
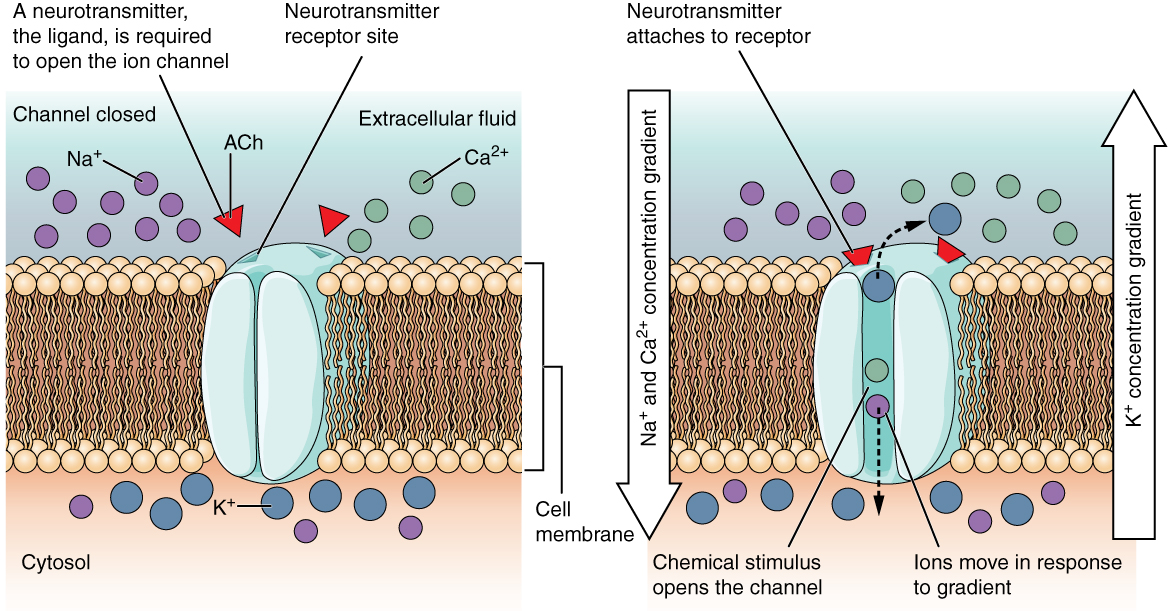
Un canal controlado por ligando se abre porque una molécula de señalización, un ligando, se une a la región extracelular del canal. Este tipo de canal también se conoce como receptor ionotrópico ya que cuando el ligando, conocido como neurotransmisor en el sistema nervioso, se une a la proteína, los iones atraviesan la membrana cambiando su carga (Figura\(\PageIndex{2}\)).
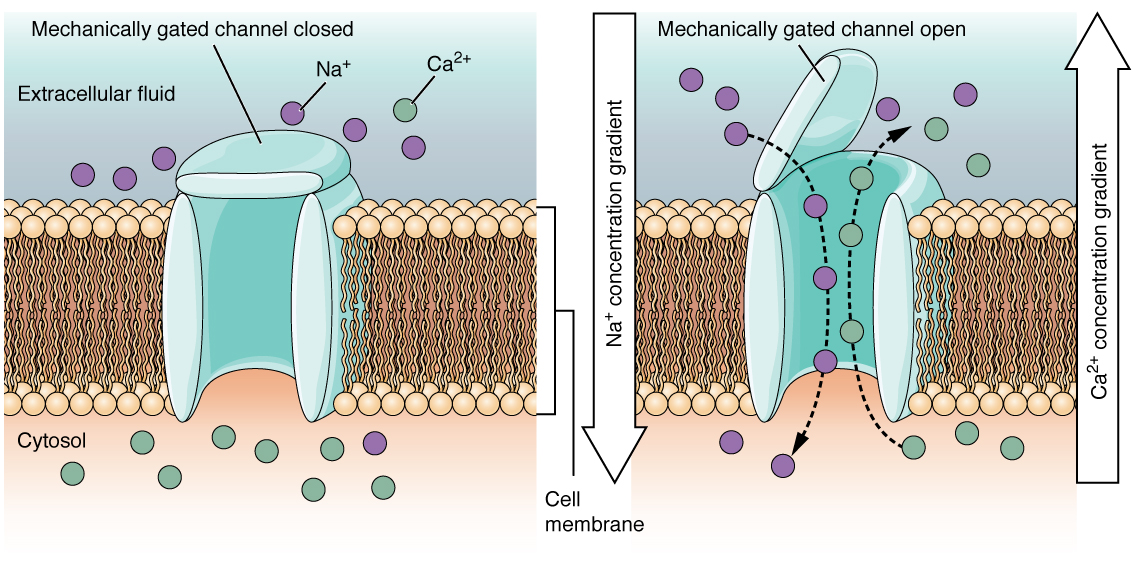
Un canal cerrado mecánicamente se abre debido a una distorsión física de la membrana celular. Muchos canales asociados con el sentido del tacto (somatosensación) están mecánicamente cerrados. Por ejemplo, a medida que se aplica presión a la piel, estos canales se abren y permiten que los iones entren en la célula. Similar a este tipo de canal sería el canal que se abre en base a los cambios de temperatura, como en probar el agua en la ducha (Figura\(\PageIndex{3}\)).
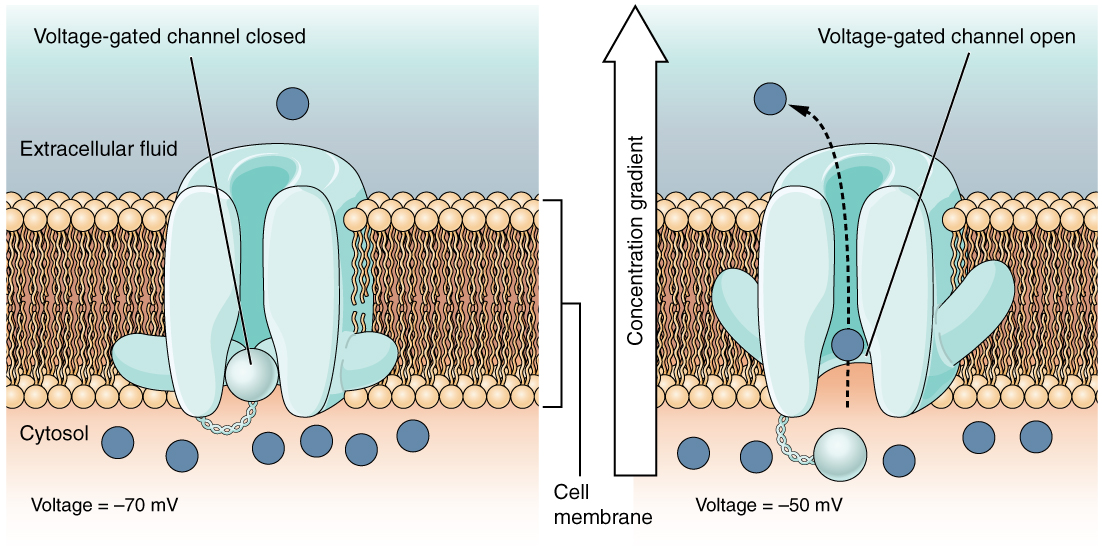
Un canal regulado por voltaje es un canal que responde a cambios en las propiedades eléctricas de la membrana en la que está incrustado. Normalmente, la porción interna de la membrana está en un voltaje negativo. Cuando ese voltaje se vuelve menos negativo, el canal comienza a permitir que los iones crucen la membrana (Figura\(\PageIndex{4}\)).
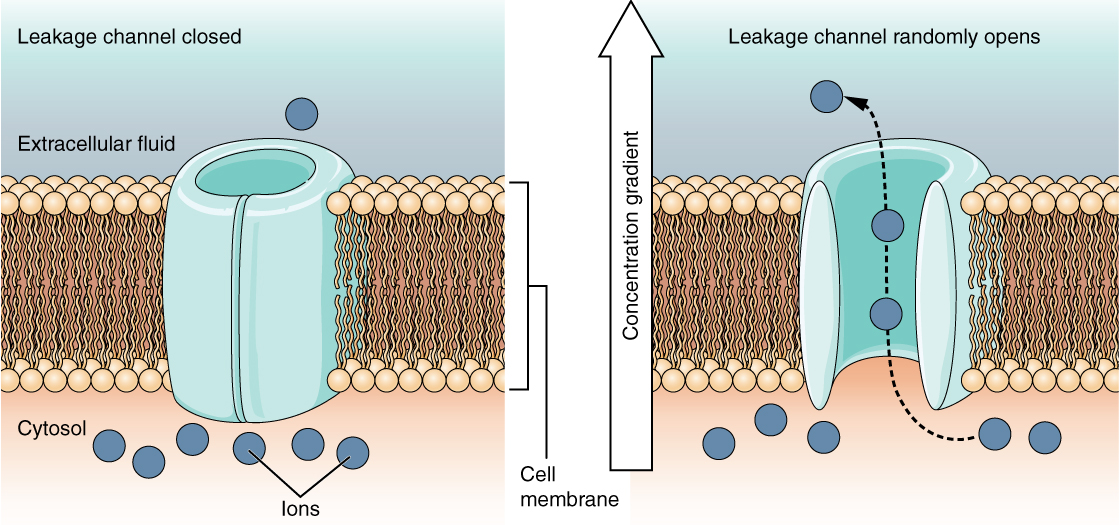
Un canal de fuga se cierra aleatoriamente, lo que significa que se abre y cierra al azar, de ahí la referencia a fugas. No hay ningún evento real que abra el canal; en cambio, tiene una tasa intrínseca de conmutación entre los estados abierto y cerrado. Los canales de fuga contribuyen al voltaje transmembrana en reposo de la membrana excitable (Figura\(\PageIndex{5}\)).
El potencial de membrana
El estado eléctrico de la membrana celular puede tener varias variaciones. Todas estas son variaciones en el potencial de membrana. Un potencial es una distribución de carga a través de la membrana celular, medida en milivoltios (mV). El estándar es comparar el interior de la célula con respecto al exterior, por lo que el potencial de membrana es un valor que representa la carga en el lado intracelular de la membrana basado en que el exterior sea cero, relativamente hablando (Figura\(\PageIndex{6}\)).
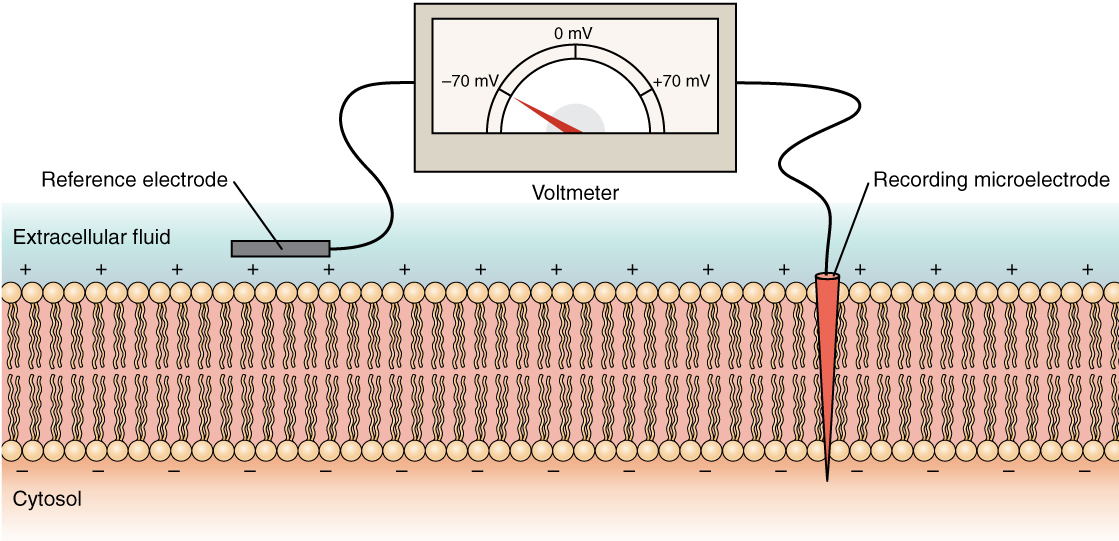
La concentración de iones en los fluidos extracelulares e intracelulares está ampliamente equilibrada, con una carga neta neutra. Sin embargo, se produce una ligera diferencia de carga justo en la superficie de la membrana, tanto interna como externamente. Es la diferencia en esta región muy limitada la que tiene todo el poder en las neuronas (y las células musculares) para generar señales eléctricas, incluyendo potenciales de acción.
Antes de que se puedan describir estas señales eléctricas, se debe explicar el estado de reposo de la membrana. Cuando la célula está en reposo y los canales iónicos están cerrados (excepto los canales de fuga que se abren aleatoriamente), los iones se distribuyen a través de la membrana de una manera muy predecible. La concentración de Na + fuera de la célula es 10 veces mayor que la concentración en el interior. Además, la concentración de K + dentro de la célula es mayor que en el exterior. El citosol contiene una alta concentración de aniones, en forma de iones fosfato y proteínas cargadas negativamente. Los aniones grandes son un componente de la membrana celular interna, incluyendo fosfolípidos especializados y proteínas asociadas con la valva interna de la membrana (valva es un término usado para un lado de la membrana bicapa lipídica). La carga negativa se localiza en los aniones grandes.
Con los iones distribuidos a través de la membrana a estas concentraciones, la diferencia de carga se mide a -70 mV, el valor descrito como el potencial de membrana en reposo. El valor exacto medido para el potencial de membrana en reposo varía entre células, pero -70 mV es el más comúnmente utilizado como este valor. Este voltaje en realidad sería mucho menor a excepción de las contribuciones de algunas proteínas importantes en la membrana. Los canales de fuga permiten que Na + se mueva lentamente hacia la celda o K + se mueva lentamente hacia afuera, y la bomba Na + /K + los restaura. Esto puede parecer un desperdicio de energía, pero cada uno tiene un papel en el mantenimiento del potencial de membrana.
El potencial de acción
El potencial de membrana en reposo describe el estado estacionario de la celda, que es un proceso dinámico que se equilibra con la fuga de iones y el bombeo de iones. Sin ninguna influencia externa, no va a cambiar. Para que se inicie una señal eléctrica, el potencial de la membrana tiene que cambiar.
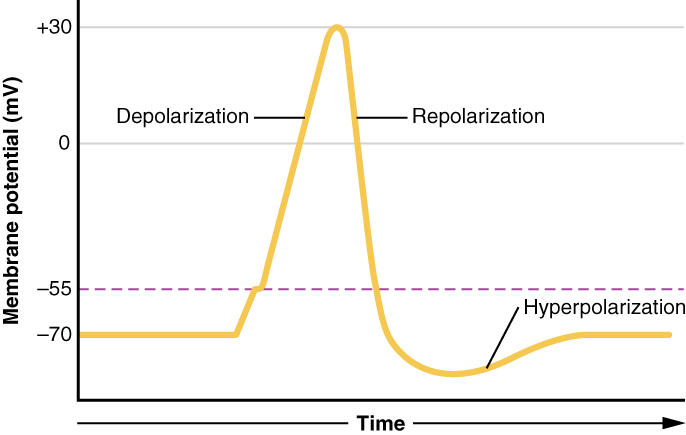
Esto comienza con una abertura de canal para Na + en la membrana. Debido a que la concentración de Na + es mayor fuera de la célula que dentro de la célula en un factor de 10, los iones se precipitarán hacia la célula que son impulsados en gran medida por el gradiente de concentración. Debido a que el sodio es un ion cargado positivamente, cambiará el voltaje relativo inmediatamente dentro de la celda en relación con inmediatamente afuera. El potencial de reposo es el estado de la membrana a un voltaje de -70 mV, por lo que el catión de sodio que ingresa a la celda provocará que se vuelva menos negativo. Esto se conoce como despolarización, lo que significa que el potencial de membrana se mueve hacia cero.
El gradiente de concentración para Na + es tan fuerte que continuará ingresando a la celda incluso después de que el potencial de membrana se haya convertido en cero, de manera que el voltaje inmediatamente alrededor del poro comienza a volverse positivo. El gradiente eléctrico también juega un papel, ya que las proteínas negativas por debajo de la membrana atraen el ion sodio. El potencial de membrana alcanzará +30 mV en el momento en que el sodio haya ingresado a la célula.
A medida que el potencial de membrana alcanza +30 mV, otros canales regulados por voltaje se abren en la membrana. Estos canales son específicos para el ion potasio. Un gradiente de concentración actúa sobre K +, también. A medida que K + comienza a salir de la celda, tomando una carga positiva con ella, el potencial de la membrana comienza a retroceder hacia su voltaje de reposo. Esto se llama repolarización, lo que significa que el voltaje de la membrana retrocede hacia el valor de -70 mV del potencial de membrana en reposo.
La repolarización devuelve el potencial de membrana al valor de -70 mV que indica el potencial de reposo, pero en realidad sobrepasa ese valor. Los iones de potasio alcanzan el equilibrio cuando el voltaje de la membrana está por debajo de -70 mV, por lo que ocurre un periodo de hiperpolarización mientras los canales K + están abiertos. Esos canales K + están ligeramente retrasados en el cierre, lo que da cuenta de este breve sobreimpulso.
Lo que se ha descrito aquí es el potencial de acción, el cual se presenta como un gráfico de voltaje a lo largo del tiempo en la Figura\(\PageIndex{6}\). Es la señal eléctrica que genera el tejido nervioso para la comunicación. El cambio en el voltaje de membrana de -70 mV en reposo a +30 mV al final de la despolarización es un cambio de 100 mV. Eso también se puede escribir como un cambio de 0.1-V. Para poner ese valor en perspectiva, piensa en una batería. Una batería AA que podrías encontrar en un mando a distancia de televisión tiene un voltaje de 1.5 V, o una batería de 9 V (la batería rectangular con dos postes en un extremo) es, obviamente, de 9 V. El cambio visto en el potencial de acción es uno o dos órdenes de magnitud menor que la carga en estas baterías. De hecho, el potencial de membrana puede describirse como una batería. Se almacena una carga a través de la membrana que puede liberarse en las condiciones correctas. Una batería de tu mando a distancia ha almacenado una carga que se “libera” cuando presionas un botón.
Ejercicio\(\PageIndex{1}\)
Lo que sucede a través de la membrana de una célula eléctricamente activa es un proceso dinámico que es difícil de visualizar con imágenes estáticas o a través de descripciones de texto. Vea esta animación para conocer más sobre este proceso. ¿Cuál es la diferencia entre la fuerza impulsora de Na+ y K+? ¿Y qué tiene de similar el movimiento de estos dos iones?
La pregunta es, ahora, ¿qué inicia el potencial de acción? La descripción anterior convenientemente pasa por alto ese punto. Pero es vital entender lo que está sucediendo. El potencial de membrana permanecerá en el voltaje de reposo hasta que algo cambie. La descripción anterior solo dice que se abre un canal Na +. Ahora bien, decir “se abre un canal” no significa que cambie una proteína transmembrana individual. En cambio, significa que se abre un tipo de canal. Existen algunos tipos diferentes de canales que permiten que Na + atraviese la membrana. Un canal Na + regulado por ligando se abrirá cuando un neurotransmisor se une a él y un canal Na + mecánicamente cerrado se abrirá cuando un estímulo físico afecte a un receptor sensorial (como la presión aplicada a la piel comprime un receptor táctil). Ya sea un neurotransmisor que se une a su proteína receptora o un estímulo sensorial que activa una célula receptora sensorial, algunos estímulos inician el proceso. El sodio comienza a entrar en la célula y la membrana se vuelve menos negativa.
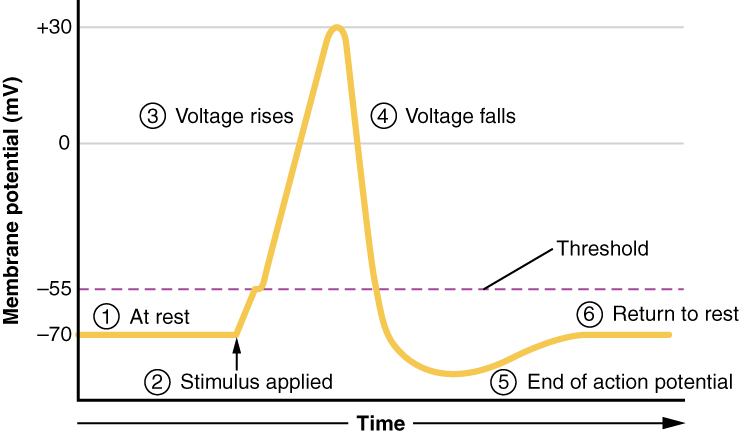
Un tercer tipo de canal que es una parte importante de la despolarización en el potencial de acción es el canal Na + regulado por voltaje. Los canales que comienzan a despolarizar la membrana debido a un estímulo ayudan a la célula a despolarizar de -70 mV a -55 mV. Una vez que la membrana alcanza ese voltaje, se abren los canales Na + regulados por voltaje. Esto es lo que se conoce como el umbral. Cualquier despolarización que no cambie el potencial de membrana a -55 mV o superior no alcanzará el umbral y por lo tanto no dará como resultado un potencial de acción. Además, cualquier estímulo que despolarice la membrana a -55 mV o más allá provocará que se abran un gran número de canales y se iniciará un potencial de acción.
Debido al umbral, el potencial de acción puede compararse con un evento digital, o sucede o no. Si no se alcanza el umbral, entonces no se produce ningún potencial de acción. Si la despolarización alcanza -55 mV, entonces el potencial de acción continúa y se extiende hasta +30 mV, a lo que K + causa repolarización, incluyendo el sobreimpulso hiperpolarizante. Además, esos cambios son los mismos para cada potencial de acción, lo que significa que una vez que se alcanza el umbral, sucede exactamente lo mismo. Un estímulo más fuerte, que podría despolarizar la membrana mucho más allá del umbral, no generará un potencial de acción “mayor”. Los potenciales de acción son “todos o ninguno”. O la membrana alcanza el umbral y todo ocurre como se describió anteriormente, o la membrana no alcanza el umbral y no pasa nada más. Todos los potenciales de acción alcanzan un pico al mismo voltaje (+30 mV), por lo que un potencial de acción no es mayor que otro. Los estímulos más fuertes iniciarán múltiples potenciales de acción más rápidamente, pero las señales individuales no son más grandes. Así, por ejemplo, no sentirás una mayor sensación de dolor, o tendrás una contracción muscular más fuerte, por el tamaño del potencial de acción porque no son de diferentes tamaños.
Como hemos visto, la despolarización y repolarización de un potencial de acción dependen de dos tipos de canales (el canal Na + regulado por voltaje y el canal K + regulado por voltaje). El canal Na + con regulación de voltaje en realidad tiene dos puertas. Una es la puerta de activación, que se abre cuando el potencial de membrana cruza -55 mV. La otra puerta es la puerta de inactivación, que se cierra después de un período de tiempo específico, del orden de una fracción de milisegundo. Cuando una celda está en reposo, la puerta de activación se cierra y la puerta de inactivación está abierta. Sin embargo, cuando se alcanza el umbral, se abre la puerta de activación, permitiendo que Na + se apresure hacia la célula. Cronometrada con el pico de despolarización, la puerta de inactivación se cierra. Durante la repolarización, no puede ingresar más sodio a la célula. Cuando el potencial de membrana vuelve a pasar -55 mV, la puerta de activación se cierra. Después de eso, la puerta de inactivación se vuelve a abrir, haciendo que el canal esté listo para comenzar de nuevo todo el proceso.
El canal K + regulado por voltaje tiene solo una puerta, que es sensible a un voltaje de membrana de -50 mV. Sin embargo, no se abre tan rápido como lo hace el canal Na + regulado por voltaje. Podría tomar una fracción de milisegundo para que el canal se abra una vez que se haya alcanzado ese voltaje. El tiempo de esto coincide exactamente con cuando el flujo de Na + alcanza su pico, por lo que los canales K + regulados por voltaje se abren justo cuando se están inactivando los canales Na + regulados por voltaje. A medida que el potencial de membrana se repolariza y el voltaje vuelve a pasar -50 mV, el canal se cierra, nuevamente, con un poco de retraso. El potasio continúa saliendo de la célula por un corto tiempo y el potencial de membrana se vuelve más negativo, lo que resulta en el sobreimpulso hiperpolarizante. Luego el canal se cierra nuevamente y la membrana puede regresar al potencial de reposo debido a la actividad continua de los canales no controlados y la bomba de Na + /K +.
Todo esto se lleva a cabo en aproximadamente 2 milisegundos (Figura\(\PageIndex{7}\)). Mientras se encuentra en curso un potencial de acción, no se puede iniciar otro. A ese efecto se le conoce como el periodo refractario. Hay dos fases del período refractario: el período refractario absoluto y el período refractario relativo. Durante la fase absoluta, no arrancará otro potencial de acción. Esto se debe a la puerta de inactivación del canal Na + regulado por voltaje. Una vez que ese canal vuelva a su conformación en reposo (menos de -55 mV), se podría iniciar un nuevo potencial de acción, pero sólo por un estímulo más fuerte que el que inició el potencial de acción actual. Esto se debe al flujo de K + fuera de la celda. Debido a que ese ion se está precipitando, cualquier Na + que intente entrar no despolarizará la célula, sino que sólo impedirá que la célula se hiperpolarice.
Propagación del Potencial de Acción
El potencial de acción se inicia al inicio del axón, en lo que se denomina segmento inicial. Hay una alta densidad de canales Na + regulados por voltaje para que se pueda llevar a cabo una rápida despolarización aquí. Bajando la longitud del axón, el potencial de acción se propaga porque se abren más canales Na + activados por voltaje a medida que se propaga la despolarización. Esta propagación se produce porque Na + entra a través del canal y se mueve a lo largo del interior de la membrana celular. A medida que el Na + se mueve, o fluye, una corta distancia a lo largo de la membrana celular, su carga positiva despolariza un poco más de la membrana celular. A medida que esa despolarización se propaga, se abren nuevos canales Na + activados por voltaje y más iones se precipitan hacia la célula, extendiendo la despolarización un poco más lejos.
Debido a que los canales Na + regulados por voltaje se inactivan en el pico de la despolarización, no se pueden abrir nuevamente por un breve tiempo, el período refractario absoluto. Debido a esto, la despolarización que se propaga hacia canales previamente abiertos no tiene ningún efecto. El potencial de acción debe propagarse hacia los terminales axónicos; como resultado, se mantiene la polaridad de la neurona, como se mencionó anteriormente.
La propagación, como se describió anteriormente, se aplica a los axones no mielinizados. Cuando la mielinización está presente, el potencial de acción se propaga de manera diferente. Los iones de sodio que ingresan a la célula en el segmento inicial comienzan a extenderse a lo largo del segmento axónico, pero no hay canales Na + regulados por voltaje hasta el primer nodo de Ranvier. Debido a que no hay apertura constante de estos canales a lo largo del segmento axónico, la despolarización se extiende a una velocidad óptima. La distancia entre nodos es la distancia óptima para mantener la membrana aún despolarizada por encima del umbral en el siguiente nodo. A medida que Na + se extiende a lo largo del interior de la membrana del segmento axónico, la carga comienza a disiparse. Si el nodo estuviera más abajo del axón, esa despolarización se habría caído demasiado para que los canales Na + regulados por voltaje se activaran en el siguiente nodo de Ranvier. Si los nodos estuvieran más cerca entre sí, la velocidad de propagación sería más lenta.
La propagación a lo largo de un axón no mielinizado se conoce como conducción continua; a lo largo de la longitud de un axón mielinado, es conducción salatoria. La conducción continua es lenta porque siempre hay apertura de canales Na + regulados por voltaje, y cada vez más Na + se precipita hacia la celda. La conducción saltatoria es más rápida porque el potencial de acción básicamente salta de un nodo a otro (sal = “saltar”), y la nueva afluencia de Na + renueva la membrana despolarizada. Junto con la mielinización del axón, el diámetro del axón puede influir en la velocidad de conducción. Al igual que el agua corre más rápido en un río ancho que en un arroyo estrecho, la despolarización basada en Na + se extiende más rápido por un axón ancho que por uno estrecho. Este concepto se conoce como resistencia y generalmente es cierto para cables eléctricos o plomería, así como lo es para los axones, aunque las condiciones específicas son diferentes en las escalas de electrones o iones frente al agua en un río.
DESBALANCE HOMEOSTÁTICO: Concentración
Las células gliales, especialmente los astrocitos, son responsables de mantener el ambiente químico del tejido del SNC. Las concentraciones de iones en el fluido extracelular son la base de cómo se establece el potencial de membrana y los cambios en la señalización electroquímica. Si se altera el equilibrio de iones, son posibles resultados drásticos.
Normalmente la concentración de K+ es mayor dentro de la neurona que en el exterior. Después de la fase de repolarización del potencial de acción, los canales de fuga de K+ y la bomba de Na+/K+ aseguran que los iones regresen a sus ubicaciones originales. Después de un accidente cerebrovascular u otro evento isquémico, los niveles extracelulares de K+ son elevados. Los astrocitos de la zona están equipados para eliminar el exceso de K+ para ayudar a la bomba. Pero cuando el nivel está muy desbalanceado, los efectos pueden ser irreversibles.
Los astrocitos pueden volverse reactivos en casos como estos, lo que perjudica su capacidad para mantener el ambiente químico local. Las células gliales se agrandan y sus procesos se hinchan. Pierden su capacidad de amortiguación K+ y la función de la bomba se ve afectada, o incluso revertida. Uno de los primeros signos de enfermedad celular es esta “fuga” de iones de sodio en las células del cuerpo. Este desequilibrio sodio/potasio afecta negativamente la química interna de las células, impidiendo que funcionen normalmente.
Visite este sitio para ver un laboratorio virtual de neurofisiología, y observar procesos electrofisiológicos en el sistema nervioso, donde los científicos miden directamente las señales eléctricas producidas por las neuronas. A menudo, los potenciales de acción ocurren tan rápidamente que ver una pantalla para verlos ocurrir no es útil. Un altavoz es alimentado por las señales grabadas de una neurona y “estalla” cada vez que la neurona dispara un potencial de acción. Estos potenciales de acción están disparando tan rápido que suena como estático en la radio. Los electrofisiólogos pueden reconocer los patrones dentro de esa estática para entender lo que está sucediendo. ¿Por qué se utiliza el modelo de sanguijuela para medir la actividad eléctrica de las neuronas en lugar de usar humanos?
Revisar
El sistema nervioso se caracteriza por señales eléctricas que se envían de una zona a otra. Ya sea que esas áreas estén cercanas o muy alejadas, la señal debe viajar a lo largo de un axón. La base de la señal eléctrica es la distribución controlada de iones a través de la membrana. Los canales iónicos transmembrana regulan cuando los iones pueden entrar o salir de la célula, de modo que se genera una señal precisa. Esta señal es el potencial de acción que tiene una forma muy característica basada en cambios de voltaje a través de la membrana en un período de tiempo dado.
La membrana está normalmente en reposo con concentraciones establecidas de Na + y K + en cada lado. Un estímulo iniciará la despolarización de la membrana, y los canales regulados por voltaje darán como resultado una mayor despolarización seguida de la repolarización de la membrana. Un ligero sobreimpulso de la hiperpolarización marca el final del potencial de acción. Mientras se encuentra en curso un potencial de acción, no se puede generar otro bajo las mismas condiciones. Mientras que el canal Na + regulado por voltaje está inactivado, absolutamente no se pueden generar potenciales de acción. Una vez que ese canal ha vuelto a su estado de reposo, es posible un nuevo potencial de acción, pero debe ser iniciado por un estímulo relativamente más fuerte para superar el K + que sale de la célula.
El potencial de acción se desplaza hacia abajo del axón a medida que los canales iónicos regulados por voltaje se abren por la despolarización en expansión. En los axones no mielinizados, esto sucede de manera continua porque hay canales regulados por voltaje a lo largo de la membrana. En los axones mielinizados, la propagación se describe como saltatoria porque los canales regulados por voltaje solo se encuentran en los nodos de Ranvier y los eventos eléctricos parecen “saltar” de un nodo al siguiente. La conducción salatoria es más rápida que la conducción continua, lo que significa que los axones mielinizados propagan sus señales más rápido. El diámetro del axón también marca la diferencia ya que los iones que se difunden dentro de la célula tienen menos resistencia en un espacio más amplio.
Preguntas de Enlace Interactivo
Lo que sucede a través de la membrana de una célula eléctricamente activa es un proceso dinámico que es difícil de visualizar con imágenes estáticas o a través de descripciones de texto. Vea esta animación para entender realmente el proceso. ¿Cuál es la diferencia entre la fuerza impulsora para Na + y K +? ¿Y qué tiene de similar el movimiento de estos dos iones?
Respuesta: El sodio se está moviendo hacia la célula debido al inmenso gradiente de concentración, mientras que el potasio se está moviendo hacia fuera por la despolarización que causa el sodio. Sin embargo, ambos bajan sus respectivos gradientes, hacia el equilibrio.
Visite este sitio para ver un laboratorio virtual de neurofisiología, y observar procesos electrofisiológicos en el sistema nervioso, donde los científicos miden directamente las señales eléctricas producidas por las neuronas. A menudo, los potenciales de acción ocurren tan rápidamente que ver una pantalla para verlos ocurrir no es útil. Un altavoz es alimentado por las señales grabadas de una neurona y “estalla” cada vez que la neurona dispara un potencial de acción. Estos potenciales de acción están disparando tan rápido que suena como estático en la radio. Los electrofisiólogos pueden reconocer los patrones dentro de esa estática para entender lo que está sucediendo. ¿Por qué se utiliza el modelo de sanguijuela para medir la actividad eléctrica de las neuronas en lugar de usar humanos?
Respuesta: Las propiedades de la electrofisiología son comunes a todos los animales, por lo que usar la sanguijuela es un enfoque más fácil y humano para estudiar las propiedades de estas células. Existen diferencias entre los sistemas nerviosos de los invertebrados (como una sanguijuela) y los vertebrados, pero no por el bien de lo que estudian estos experimentos.
Preguntas de revisión
P: ¿Qué ion entra en una neurona provocando la despolarización de la membrana celular?
A. sodio
B. cloruro
C. potasio
D. fosfato
Respuesta: A
P. ¿Los canales Na + de voltaje se abren al llegar a qué estado?
A. potencial de descanso
B. umbral
C. repolarización
D. sobreimpulso
Respuesta: B
P: ¿Qué requiere un canal con puerta de ligando para poder abrirse?
A. aumento de la concentración de iones Na +
B. unión de un neurotransmisor
C. aumento en la concentración de iones K +
D. despolarización de la membrana
Respuesta: B
P: ¿A qué responde un canal cerrado mecánicamente?
A. estímulo físico
B. estímulo químico
C. aumento de la resistencia
D. disminución de la resistencia
Respuesta: A
P. ¿Cuál de los siguientes voltajes probablemente se mediría durante el periodo refractario relativo?
A. +30 mV
B. 0 mV
C. -45 mV
D. -80 mv
Respuesta: D
P: ¿Cuál de las siguientes probablemente va a propagar un potencial de acción más rápido?
A. un axón delgado y no mielinizado
B. un axón delgado y mielinizado
C. un axón grueso y no mielinizado
D. un axón espeso y mielinizado
Respuesta: D
Preguntas de Pensamiento Crítico
P: ¿Qué significa que un potencial de acción sea un evento de “todo o ninguno”?
A. La membrana celular debe alcanzar el umbral antes de que se abran los canales Na + regulados por voltaje. Si no se alcanza el umbral, esos canales no se abren, y no se produce la fase despolarizante del potencial de acción, la membrana celular simplemente volverá a su estado de reposo.
P. La percepción consciente del dolor a menudo se retrasa debido al tiempo que tardan las sensaciones en llegar a la corteza cerebral. ¿Por qué sería este el caso basado en la propagación del potencial axónico?
A. Los axones de las neuronas sensoriales sensibles al dolor son delgados y no mielinizados, de modo que esa sensación tarda más en llegar al cerebro que otras sensaciones.
Glosario
- período refractario absoluto
- tiempo durante un período de acción cuando no se puede generar otro potencial de acción porque el canal Na + regulado por voltaje está inactivado
- puerta de activación
- parte del canal Na + regulado por voltaje que se abre cuando el voltaje de la membrana alcanza el umbral
- conducción continua
- propagación lenta de un potencial de acción a lo largo de un axón no mielinado debido a canales Na + regulados por voltaje ubicados a lo largo de toda la longitud de la membrana celular
- despolarización
- cambio en un potencial de membrana celular desde reposo hacia cero
- exclusión electroquímica
- principio de permitir selectivamente iones a través de un canal sobre la base de su carga
- membrana excitable
- membrana celular que regula el movimiento de iones para que se pueda generar una señal eléctrica
- cerrado
- propiedad de un canal que determina cómo se abre bajo condiciones específicas, como cambio de voltaje o deformación física
- puerta de inactivación
- parte de un canal Na + regulado por voltaje que se cierra cuando el potencial de membrana alcanza +30 mV
- receptor ionotrópico
- receptor del neurotransmisor que actúa como una puerta del canal iónico, y se abre por la unión del neurotransmisor
- canal de fuga
- canal iónico que se abre aleatoriamente y no se conecta a un evento específico, también conocido como un canal no regulado
- canales de ligando-cerrado
- otro nombre para un receptor ionotrópico para el que un neurotransmisor es el ligando
- canal cerrado mecánicamente
- canal iónico que se abre cuando un evento físico afecta directamente la estructura de la proteína
- potencial de membrana
- distribución de carga a través de la membrana celular, basada en las cargas de iones
- canal inespecífico
- canal que no es específico de un ion sobre otro, tal como un canal catiónico inespecífico que permite cualquier ion cargado positivamente a través de la membrana
- periodo refractario
- tiempo después del inicio de un potencial de acción cuando no se puede generar otro potencial de acción
- período refractario relativo
- tiempo durante el período refractario cuando un nuevo potencial de acción solo puede ser iniciado por un estímulo más fuerte que el potencial de acción actual porque los canales K + regulados por voltaje no están cerrados
- repolarización
- retorno del potencial de membrana a su voltaje normalmente negativo al final del potencial de acción
- resistencia
- propiedad de un axón que se relaciona con la capacidad de las partículas para difundirse a través del citoplasma; esto es inversamente proporcional al diámetro de la fibra
- potencial de membrana en reposo
- la diferencia de voltaje medida a través de una membrana celular en condiciones de estado estacionario, típicamente -70 mV
- conducción saltatoria
- propagación rápida del potencial de acción a lo largo de un axón mielinado debido a que los canales Na + regulados por voltaje están presentes solo en los nodos de Ranvier
- exclusión de tamaño
- principio de permitir selectivamente iones a través de un canal sobre la base de su tamaño relativo
- canal de voltaje
- canal iónico que se abre debido a un cambio en la carga distribuida a través de la membrana donde se encuentra