
12.5: Comunicación entre neuronas
- Page ID
- 123051

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Explicar las diferencias entre los tipos de potenciales graduados
- Categorizar los principales neurotransmisores por tipo químico y efecto
Los cambios eléctricos que se producen dentro de una neurona, como se describe en la sección anterior, son similares a que se enciende un interruptor de luz. Un estímulo inicia la despolarización, pero el potencial de acción corre por sí solo una vez que se ha alcanzado un umbral. La pregunta es ahora: “¿Qué enciende el interruptor de la luz?” Los cambios temporales en el voltaje de la membrana celular pueden ser el resultado de que las neuronas reciban información del ambiente, o de la acción de una neurona sobre otra. Estos tipos especiales de potenciales influyen en una neurona y determinan si ocurrirá un potencial de acción o no. Muchas de estas señales transitorias se originan en la sinapsis.
Potenciales graduados
Los cambios locales en el potencial de membrana se denominan potenciales graduados y generalmente se asocian con las dendritas de una neurona. La cantidad de cambio en el potencial de membrana está determinada por el tamaño del estímulo que lo provoca. En el ejemplo de probar la temperatura de la ducha, el agua ligeramente tibia solo iniciaría un pequeño cambio en un termorreceptor, mientras que el agua caliente provocaría una gran cantidad de cambio en el potencial de membrana.
Los potenciales graduados pueden ser de dos clases, ya sea despolarizantes o hiperpolarizantes (Figura\(\PageIndex{1}\)). Para una membrana en el potencial de reposo, un potencial gradual representa un cambio en ese voltaje ya sea por encima de -70 mV o por debajo de -70 mV. Los potenciales graduados despolarizantes son a menudo el resultado de la entrada de Na + o Ca 2+ en la célula. Ambos iones tienen concentraciones más altas fuera de la célula que dentro; debido a que tienen una carga positiva, se moverán hacia la célula provocando que se vuelva menos negativa en relación con el exterior. Los potenciales graduados hiperpolarizantes pueden ser causados por K + dejando la célula o Cl - entrando en la célula. Si una carga positiva sale de una celda, la celda se vuelve más negativa; si entra una carga negativa en la celda, sucede lo mismo.

Figura\(\PageIndex{1}\): Potenciales Gradados. Los potenciales graduados son cambios temporales en el voltaje de la membrana, cuyas características dependen del tamaño del estímulo. Algunos tipos de estímulos provocan despolarización de la membrana, mientras que otros causan hiperpolarización. Depende de los canales iónicos específicos que se activan en la membrana celular.
Tipos de Potenciales Gradados
Para las células unipolares de las neuronas sensoriales, tanto las que tienen terminaciones nerviosas libres como las que están dentro de las encapsulaciones, se desarrollan potenciales graduados en las dendritas que influyen en la generación de un potencial de acción en el axón de la misma célula. Esto se llama potencial generador. Para otras células receptoras sensoriales, como las células gustativas o los fotorreceptores de la retina, los potenciales graduados en sus membranas dan como resultado la liberación de neurotransmisores en las sinapsis con neuronas sensoriales. Esto se llama potencial receptor.
Un potencial postsináptico (PSP) es el potencial gradual en las dendritas de una neurona que está recibiendo sinapsis de otras células. Los potenciales postsinápticos pueden ser despolarizantes o hiperpolarizantes. La despolarización en un potencial postsináptico se denomina potencial postsináptico excitatorio (EPSP) porque hace que el potencial de la membrana se mueva hacia el umbral. La hiperpolarización en un potencial postsináptico es un potencial postsináptico inhibitorio (IPSP) porque hace que el potencial de membrana se aleje del umbral.
Suma
Todos los tipos de potenciales graduados resultarán en pequeños cambios de despolarización o hiperpolarización en el voltaje de una membrana. Estos cambios pueden llevar a que la neurona alcance el umbral si los cambios se suman, o se suman. Los efectos combinados de diferentes tipos de potenciales graduados se ilustran en la Figura\(\PageIndex{2}\). Si el cambio total de voltaje en la membrana es positivo de 15 mV, lo que significa que la membrana se despolariza de -70 mV a -55 mV, entonces los potenciales graduados resultarán en que la membrana alcance el umbral.
Para los potenciales receptores, el umbral no es un factor porque el cambio en el potencial de membrana para las células receptoras causa directamente la liberación de neurotransmisores. Sin embargo, los potenciales generadores pueden iniciar potenciales de acción en el axón neuronal sensorial, y los potenciales postsinápticos pueden iniciar un potencial de acción en el axón de otras neuronas. Los potenciales graduados se suman en una ubicación específica al inicio del axón para iniciar el potencial de acción, es decir, el segmento inicial. Para las neuronas sensoriales, que no tienen un cuerpo celular entre las dendritas y el axón, el segmento inicial es directamente adyacente a las terminaciones dendríticas. Para todas las demás neuronas, el montículo del axón es esencialmente el segmento inicial del axón, y es donde tiene lugar la suma. Estas ubicaciones tienen una alta densidad de canales Na + regulados por voltaje que inician la fase de despolarización del potencial de acción.

Figura\(\PageIndex{2}\): Suma de Potencial Postsináptico. El resultado de la suma de potenciales postsinápticos es el cambio general en el potencial de membrana. En el punto A, varios potenciales postsinápticos excitatorios diferentes se suman a una gran despolarización. En el punto B, una mezcla de potenciales postsinápticos excitatorios e inhibitorios dan como resultado un resultado final diferente para el potencial de membrana.
La suma puede ser espacial o temporal, lo que significa que puede ser el resultado de múltiples potenciales graduados en diferentes ubicaciones de la neurona, o todos en el mismo lugar pero separados en el tiempo. La suma espacial se relaciona con asociar la actividad de múltiples entradas a una neurona entre sí. La suma temporal es la relación de múltiples potenciales de acción de una sola célula dando como resultado un cambio significativo en el potencial de membrana. La suma espacial y temporal también pueden actuar juntas.
Sinapsis
Hay dos tipos de conexiones entre celdas eléctricamente activas, sinapsis químicas y sinapsis eléctricas. En una sinapsis química, una señal química, es decir, un neurotransmisor, se libera de una célula y afecta a la otra célula. En una sinapsis eléctrica, existe una conexión directa entre las dos células para que los iones puedan pasar directamente de una célula a la siguiente. Si una célula se despolariza en una sinapsis eléctrica, la célula unida también se despolariza porque los iones pasan entre las células. Las sinapsis químicas implican la transmisión de información química de una célula a otra. Esta sección se concentrará en el tipo químico de sinapsis.
Un ejemplo de sinapsis química es la unión neuromuscular (NMJ) descrita en el capítulo sobre tejido muscular. En el sistema nervioso, hay muchas más sinapsis que son esencialmente las mismas que la NMJ. Todas las sinapsis tienen características comunes, las cuales se pueden resumir en esta lista:
- elemento presináptico
- neurotransmisor (empaquetado en vesículas)
- hendidura sináptica
- proteínas receptoras
- elemento postsináptico
- eliminación o recaptación de neurotransmisores
Para el NMJ, estas características son las siguientes: el elemento presináptico son los terminales axones de la neurona motora, el neurotransmisor es acetilcolina, la hendidura sináptica es el espacio entre las células donde se difunde el neurotransmisor, la proteína receptora es el receptor nicotínico de acetilcolina, el El elemento postsináptico es el sarcolema de la célula muscular, y el neurotransmisor es eliminado por la acetilcolinesterasa. Otras sinapsis son similares a esta, y los específicos son diferentes, pero todas contienen las mismas características.
Liberación de neurotransmisores
Cuando un potencial de acción alcanza los terminales del axón, los canales de Ca 2+ regulados por voltaje en la membrana de la bombilla sináptica se abren. La concentración de Ca 2+ aumenta dentro del bulbo final, y el ion Ca 2+ se asocia con proteínas en la superficie externa de las vesículas de neurotransmisores. El Ca 2+ facilita la fusión de la vesícula con la membrana presináptica para que el neurotransmisor se libere a través de la exocitosis en el pequeño hueco entre las células, conocido como hendidura sináptica.
Una vez en la hendidura sináptica, el neurotransmisor difunde la corta distancia a la membrana postsináptica y puede interactuar con los receptores de neurotransmisores. Los receptores son específicos para el neurotransmisor, y los dos encajan como una llave y una cerradura. Un neurotransmisor se une a su receptor y no se unirá a receptores para otros neurotransmisores, haciendo de la unión un evento químico específico (Figura\(\PageIndex{3}\)).
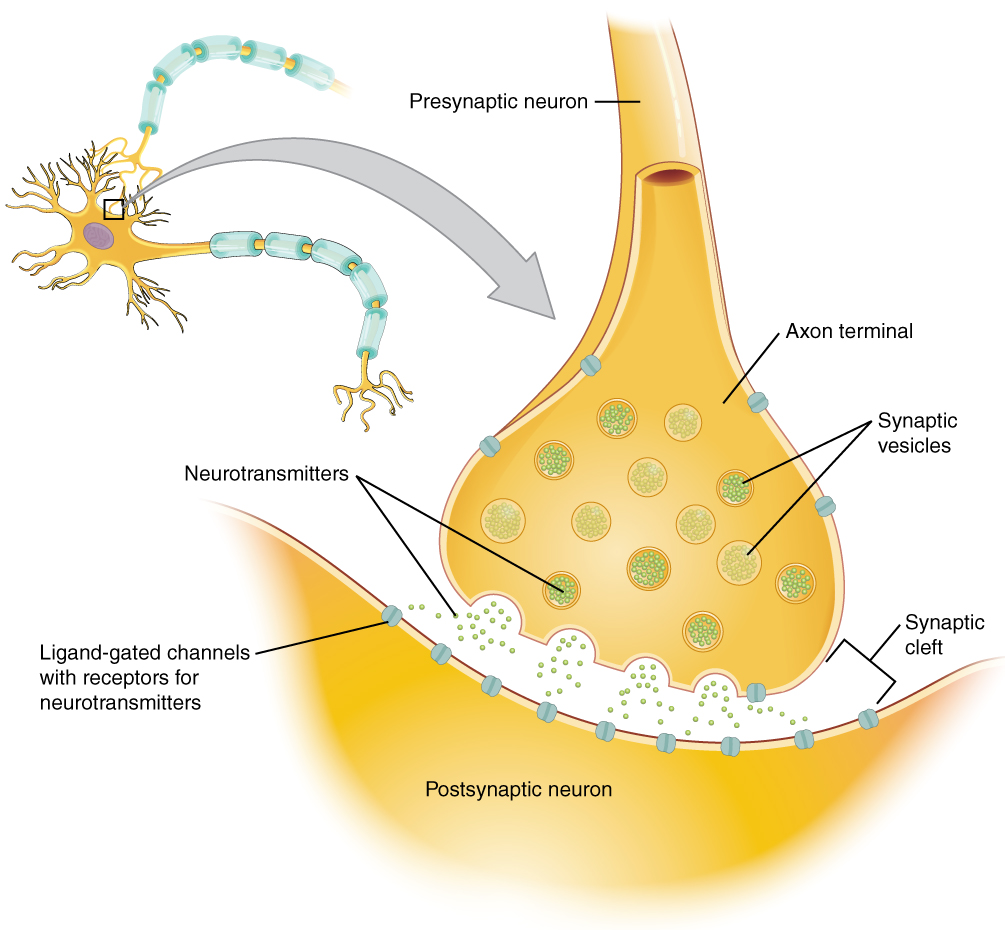
Figura\(\PageIndex{3}\): La sinapsis. La sinapsis es una conexión entre una neurona y su célula diana (que no es necesariamente una neurona). El elemento presináptico es el bulbo sináptico del axón donde Ca 2+ ingresa al bulbo para causar fusión de vesículas y liberación de neurotransmisores. El neurotransmisor se difunde a través de la hendidura sináptica para unirse a su receptor. El neurotransmisor se elimina de la sinapsis ya sea por degradación enzimática, recaptación neuronal o recaptación glial.
Sistemas de neurotransmisores
Existen varios sistemas de neurotransmisores que se encuentran en diversas sinapsis del sistema nervioso. Estos grupos se refieren a las sustancias químicas que son los neurotransmisores, y dentro de los grupos se encuentran sistemas específicos.
El primer grupo, que es un sistema neurotransmisor propio, es el sistema colinérgico. Es el sistema basado en acetilcolina. Esto incluye a la NMJ como ejemplo de sinapsis colinérgica, pero las sinapsis colinérgicas se encuentran en otras partes del sistema nervioso. Se encuentran en el sistema nervioso autónomo, además de distribuirse por todo el cerebro.
El sistema colinérgico tiene dos tipos de receptores, el receptor nicotínico se encuentra en el NMJ así como en otras sinapsis. También existe un receptor de acetilcolina conocido como el receptor muscarínico. Ambos receptores llevan el nombre de fármacos que interactúan con el receptor además de la acetilcolina. La nicotina se unirá al receptor nicotínico y lo activará de manera similar a la acetilcolina. La muscarina, producto de ciertos hongos, se unirá al receptor muscarínico. Sin embargo, la nicotina no se unirá al receptor muscarínico y la muscarina no se unirá al receptor nicotínico.
Otro grupo de neurotransmisores son los aminoácidos. Esto incluye glutamato (Glu), GABA (ácido gamma-aminobutírico, un derivado del glutamato) y glicina (Gly). Estos aminoácidos tienen un grupo amino y un grupo carboxilo en sus estructuras químicas. El glutamato es uno de los 20 aminoácidos que se utilizan para elaborar proteínas. Cada neurotransmisor de aminoácidos formaría parte de su propio sistema, a saber, los sistemas glutamatérgico, GABAérgico y glicinérgico. Cada uno tiene sus propios receptores y no interactúan entre sí. Los neurotransmisores de aminoácidos se eliminan de la sinapsis por recaptación. Una bomba en la membrana celular del elemento presináptico, o a veces una célula glial vecina, limpiará el aminoácido de la hendidura sináptica para que pueda ser reciclado, reempaquetado en vesículas y liberado nuevamente.
Otra clase de neurotransmisores es la amina biogénica, un grupo de neurotransmisores que están hechos enzimáticamente a partir de aminoácidos. Tienen grupos amino en ellos, pero ya no tienen grupos carboxilo y por lo tanto ya no se clasifican como aminoácidos. La serotonina está hecha de triptófano. Es la base del sistema serotoninérgico, que tiene sus propios receptores específicos. La serotonina se transporta de nuevo a la célula presináptica para su reempaquetado.
Otras aminas biogénicas están hechas de tirosina, e incluyen dopamina, norepinefrina y epinefrina. La dopamina es parte de su propio sistema, el sistema dopaminérgico, que tiene receptores de dopamina. La dopamina se elimina de la sinapsis mediante el transporte de proteínas en la membrana celular presináptica. La norepinefrina y la epinefrina pertenecen al sistema neurotransmisor adrenérgico. Las dos moléculas son muy similares y se unen a los mismos receptores, a los que se hace referencia como receptores alfa y beta. La norepinefrina y la epinefrina también se transportan de regreso a la célula presináptica. La epinefrina química (epi- = “on”; “-nefrina” = riñón) también se conoce como adrenalina (renal = “riñón”), y la norepinefrina a veces se conoce como noradrenalina. La glándula suprarrenal produce epinefrina y norepinefrina para ser liberadas en el torrente sanguíneo como hormonas.
Un neuropéptido es una molécula neurotransmisora compuesta por cadenas de aminoácidos conectadas por enlaces peptídicos. Esto es lo que es una proteína, pero el término proteína implica una cierta longitud a la molécula. Algunos neuropéptidos son bastante cortos, como la met-encefalina, que tiene cinco aminoácidos de longitud. Otros son largos, como la beta-endorfina, que tiene 31 aminoácidos de longitud. Los neuropéptidos a menudo se liberan en las sinapsis en combinación con otro neurotransmisor, y a menudo actúan como hormonas en otros sistemas del cuerpo, como el péptido intestinal vasoactivo (VIP) o la sustancia P.
El efecto de un neurotransmisor sobre el elemento postsináptico depende completamente de la proteína receptora. Primero, si no hay proteína receptora en la membrana del elemento postsináptico, entonces el neurotransmisor no tiene ningún efecto. El efecto despolarizante o hiperpolarizante también depende del receptor. Cuando la acetilcolina se une al receptor nicotínico, la célula postsináptica se despolariza. Esto se debe a que el receptor es un canal catiónico y Na + cargado positivamente se precipitará hacia la célula. Sin embargo, cuando la acetilcolina se une al receptor muscarínico, del cual hay varias variantes, podría provocar despolarización o hiperpolarización de la célula diana.
Los neurotransmisores de aminoácidos, glutamato, glicina y GABA, están asociados casi exclusivamente con un solo efecto. El glutamato se considera un aminoácido excitatorio, pero sólo porque los receptores Glu en el adulto provocan despolarización de la célula postsináptica. La glicina y el GABA son considerados aminoácidos inhibitorios, nuevamente porque sus receptores causan hiperpolarización.
Las aminas biogénicas tienen efectos mixtos. Por ejemplo, los receptores de dopamina que se clasifican como receptores D1 son excitadores mientras que los receptores de tipo D2 son inhibidores. Los receptores biogénicos de amina y los receptores neuropeptídicos pueden tener efectos aún más complejos porque algunos pueden no afectar directamente el potencial de membrana, sino que tienen un efecto sobre la transcripción génica u otros procesos metabólicos en la neurona. Las características de los diversos sistemas neurotransmisores que se presentan en esta sección se organizan en la Tabla\(\PageIndex{1}\).
Lo importante a recordar sobre los neurotransmisores, y los químicos de señalización en general, es que el efecto es totalmente dependiente del receptor. Los neurotransmisores se unen a una de dos clases de receptores en la superficie celular, ionotrópicos o metabotrópicos (Figura\(\PageIndex{4}\)). Los receptores ionotrópicos son canales iónicos activados por ligandos, tales como el receptor nicotínico para acetilcolina o el receptor de glicina. Un receptor metabotrópico involucra un complejo de proteínas que resultan en cambios metabólicos dentro de la célula. El complejo receptor incluye la proteína receptora transmembrana, una proteína G y una proteína efectora. El neurotransmisor, referido como el primer mensajero, se une a la proteína receptora en la superficie extracelular de la célula, y el lado intracelular de la proteína inicia la actividad de la proteína G. La proteína G es una hidrolasa de guanosina trifosfato (GTP) que se mueve físicamente de la proteína receptora a la proteína efectora para activar esta última. Una proteína efectora es una enzima que cataliza la generación de una nueva molécula, que actúa como el mediador intracelular de la señal que se une al receptor. A este mediador intracelular se le llama el segundo mensajero.
Diferentes receptores utilizan diferentes segundos mensajeros. Dos ejemplos comunes de segundos mensajeros son adenosina monofosfato cíclico (AMPc) y trifosfato de inositol (IP 3). La enzima adenilato ciclasa (un ejemplo de una proteína efectora) produce AMPc, y la fosfolipasa C es la enzima que produce IP 3. Segundos mensajeros, después de que son producidos por la proteína efectora, provocan cambios metabólicos dentro de la célula. Estos cambios son muy probablemente la activación de otras enzimas en la célula. En las neuronas, a menudo modifican los canales iónicos, ya sea abriéndolos o cerrándolos. Estas enzimas también pueden provocar cambios en la célula, como la activación de genes en el núcleo, y por lo tanto el aumento de la síntesis de proteínas. En las neuronas, este tipo de cambios suelen ser la base de conexiones más fuertes entre las células en la sinapsis y pueden ser la base del aprendizaje y la memoria.

Figura\(\PageIndex{4}\): Tipos de receptores. (a) Un receptor ionotrópico es un canal que se abre cuando el neurotransmisor se une a él. (b) Un receptor metabotrópico es un complejo que provoca cambios metabólicos en la célula cuando el neurotransmisor se une a ella (1). Después de la unión, la proteína G hidroliza GTP y se mueve a la proteína efectora (2). Cuando la proteína G entra en contacto con la proteína efectora, se genera un segundo mensajero, como AMPc (3). El segundo mensajero puede entonces provocar cambios en la neurona, como abrir o cerrar canales iónicos, cambios metabólicos y cambios en la transcripción génica.
| Sistema | Colinérgico | Aminoácidos | Aminas biogénicas | Neuropéptidos |
|---|---|---|---|---|
| Neurotransmisores | Acetilcolina | Glutamato, glicina, GABA | Serotonina (5-HT), dopamina, norepinefrina, (epinefrina) | Met-encefalina, beta-endorfina, VIP, Sustancia P, etc. |
| Receptores | Receptores nicotínicos y muscarínicos | Receptores Glu, receptores gly, receptores GABA | Receptores 5-HT, receptores D1 y D2, receptores α-adrenérgicos y β-adrenérgicos | Los receptores son demasiado numerosos para enumerarlos, pero son específicos de los péptidos. |
| Eliminación | Degradación por acetilcolinesterasa | Recaptación por neuronas o glía | Recaptación por neuronas | Degradación por enzimas llamadas peptidasas |
| Efecto postsináptico | El receptor nicotínico provoca despolarización. Los receptores muscarínicos pueden causar tanto despolarización como hiperpolarización dependiendo del subtipo. | Los receptores Glu provocan despolarización. Los receptores Gly y GABA causan hiperpolarización. | La despolarización o hiperpolarización depende del receptor específico. Por ejemplo, los receptores D1 causan despolarización y los receptores D2 causan hiperpolarización. | La despolarización o hiperpolarización depende del receptor específico. |
TRASTORNOS DE LA... Sistema Nervioso
La causa subyacente de algunas enfermedades neurodegenerativas, como el Alzheimer y el Parkinson, parece estar relacionada con las proteínas, específicamente, con las proteínas que se comportan mal. Una de las teorías más fuertes de lo que causa la enfermedad de Alzheimer se basa en la acumulación de placas beta-amiloides, conglomerados densos de una proteína que no está funcionando correctamente. La enfermedad de Parkinson está ligada a un aumento de una proteína conocida como alfa-sinucleína que es tóxica para las células del núcleo de la sustancia nigra en el mesencéfalo.
Para que las proteínas funcionen correctamente, dependen de su forma tridimensional. La secuencia lineal de aminoácidos se pliega en una forma tridimensional que se basa en las interacciones entre esos aminoácidos. Cuando se altera el plegamiento, y las proteínas adquieren una forma diferente, dejan de funcionar correctamente. Pero la enfermedad no es necesariamente el resultado de la pérdida funcional de estas proteínas; más bien, estas proteínas alteradas comienzan a acumularse y pueden volverse tóxicas. Por ejemplo, en el Alzheimer, el sello distintivo de la enfermedad es la acumulación de estas placas amiloides en la corteza cerebral. El término acuñado para describir este tipo de enfermedades es “proteopatía” e incluye otras enfermedades. La enfermedad de Creutzfeld-Jacob, la variante humana de la enfermedad priónica conocida como enfermedad de las vacas locas en el bovino, también implica la acumulación de placas amiloides, similares a la enfermedad de Alzheimer, así como enfermedades de otros sistemas orgánicos, como la fibrosis quística o la diabetes tipo 2. El reconocimiento de la relación entre estas enfermedades ha sugerido nuevas posibilidades terapéuticas. Interferir con la acumulación de las proteínas, y posiblemente tan pronto como su producción original dentro de la célula, puede desbloquear nuevas formas de aliviar estas devastadoras enfermedades.
Revisión del Capítulo
La base de la señal eléctrica dentro de una neurona es el potencial de acción que se propaga por el axón. Para que una neurona genere un potencial de acción, necesita recibir aportes de otra fuente, ya sea otra neurona o un estímulo sensorial. Esa entrada dará como resultado la apertura de canales iónicos en la neurona, resultando en un potencial gradual basado en la fuerza del estímulo. Los potenciales graduados pueden ser despolarizantes o hiperpolarizantes y pueden sumarse para afectar la probabilidad de que la neurona alcance el umbral.
Los potenciales graduados pueden ser el resultado de estímulos sensoriales. Si el estímulo sensorial es recibido por las dendritas de una neurona sensorial unipolar, como la neurona sensorial que termina en la piel, el potencial gradual se denomina potencial generador porque puede generar directamente el potencial de acción en el segmento inicial del axón. Si el estímulo sensorial es recibido por una célula receptora sensorial especializada, el potencial clasificado se denomina potencial receptor. Los potenciales graduados producidos por las interacciones entre neuronas en las sinapsis se denominan potenciales postsinápticos (PSP). Un potencial gradual despolarizante en una sinapsis se llama PSP excitatoria, y un potencial gradual hiperpolarizante en una sinapsis se llama PSP inhibitoria.
Las sinapsis son los contactos entre neuronas, que pueden ser de naturaleza química o eléctrica. Las sinapsis químicas son mucho más comunes. En una sinapsis química, el neurotransmisor se libera del elemento presináptico y se difunde a través de la hendidura sináptica. El neurotransmisor se une a una proteína receptora y provoca un cambio en la membrana postsináptica (la PSP). El neurotransmisor debe ser inactivado o eliminado de la hendidura sináptica para que el estímulo sea limitado en el tiempo.
Las características particulares de una sinapsis varían en función del sistema neurotransmisor producido por esa neurona. El sistema colinérgico se encuentra en la unión neuromuscular y en ciertos lugares dentro del sistema nervioso. Los aminoácidos, como el glutamato, la glicina y el ácido gamma-aminobutírico (GABA) se utilizan como neurotransmisores. Otros neurotransmisores son el resultado de que los aminoácidos se cambian enzimáticamente, como en las aminas biogénicas, o que se unen covalentemente entre sí, como en los neuropéptidos.
Preguntas de Enlace Interactivo
Mira este video para conocer la suma. El proceso de conversión de señales eléctricas en señales químicas y de retorno requiere cambios sutiles que pueden resultar en aumentos transitorios o disminuciones en el voltaje de la membrana. Para provocar un cambio duradero en la celda objetivo, generalmente se suman o se suman múltiples señales. ¿La suma espacial tiene que ocurrir de una vez, o las señales separadas pueden llegar a la neurona postsináptica en momentos ligeramente diferentes? Explica tu respuesta.
Respuesta: Una segunda señal de una neurona presináptica separada puede llegar un poco más tarde, siempre y cuando llegue antes de que la primera muera, o se disipe.
Mira este video para conocer la liberación de un neurotransmisor. El potencial de acción llega al final del axón, llamado terminal del axón, y se libera una señal química para decirle a la célula diana que haga algo, ya sea que inicie un nuevo potencial de acción, o que suprima esa actividad. En un espacio muy corto, la señal eléctrica del potencial de acción se cambia a la señal química de un neurotransmisor, y luego de vuelta a los cambios eléctricos en la membrana de la célula diana. ¿Cuál es la importancia de los canales de calcio activados por voltaje en la liberación de neurotransmisores?
Respuesta: El potencial de acción despolariza la membrana celular del terminal axón, que contiene el canal Ca 2+ regulado por voltaje. Ese cambio de voltaje abre el canal para que Ca 2+ pueda ingresar al terminal axón. Los iones de calcio permiten que las vesículas sinápticas liberen su contenido a través de exocitosis.
Preguntas de revisión
P. ¿Cuánto de un cambio en el potencial de membrana es necesario para que la suma de potenciales postsinápticos resulte en la generación de un potencial de acción?
A. +30 mV
B. +15 mV
C. +10 mV
D. -15 mV
Respuesta: B
P. Se abre un canal sobre una membrana postsináptica que hace que un ion negativo entre en la célula. ¿Qué tipo de potencial calificado es este?
A. despolarización
B. repolarización
C. hiperpolarizante
D. no polarizante
Respuesta: C
P. ¿Qué neurotransmisor se libera en la unión neuromuscular?
A. norepinefrina
B. serotonina
C. dopamina
D. acetilcolina
Respuesta: D
P. ¿Qué tipo de receptor requiere una proteína efectora para iniciar una señal?
A. amina biogénica
B. receptor ionotrópico
C. sistema colinérgico
D. receptor metabotrópico
Respuesta: D
P. ¿Cuál de los siguientes neurotransmisores se asocia exclusivamente con la inhibición?
A. GABA
B. acetilcolina
C. glutamato
D. norepinefrina
Respuesta: A
Preguntas de Pensamiento Crítico
P. Si una célula postsináptica tiene sinapsis de cinco células diferentes, y tres causan EPSP y dos de ellas causan IPSP, dar un ejemplo de una serie de despolarizaciones e hiperpolarizaciones que resultarían en que la neurona alcanzara el umbral.
A. EPSP1 = +5 mV, EPSP2 = +7 mV, EPSP 3 = +10 mV, IPSP1 = -4 mV, IPSP2 = -3 mV. 5 + 7 + 10 — 4 — 3 = +15 mV.
P. ¿Por qué el receptor es el elemento importante que determina el efecto que tiene un neurotransmisor en una célula diana?
A. Diferentes neurotransmisores tienen diferentes receptores. Así, el tipo de receptor en la célula postsináptica es lo que determina qué canales iónicos se abren. La unión de acetilcolina al receptor nicotínico provoca que los cationes crucen la membrana. La unión de GABA a su receptor hace que el cloruro de anión atraviese la membrana.
Glosario
- amina biogénica
- clase de neurotransmisores derivados enzimáticamente de aminoácidos pero que ya no contienen un grupo carboxilo
- sinapsis química
- conexión entre dos neuronas, o entre una neurona y su diana, donde un neurotransmisor se difunde a través de una distancia muy corta
- sistema colinérgico
- sistema neurotransmisor de acetilcolina, que incluye sus receptores y la enzima acetilcolinesterasa
- proteína efectora
- enzima que cataliza la generación de una nueva molécula, que actúa como el mediador intracelular de la señal que se une al receptor
- sinapsis eléctrica
- conexión entre dos neuronas, o cualquiera de dos células eléctricamente activas, donde los iones fluyen directamente a través de canales que abarcan sus membranas celulares adyacentes
- Potencial postsináptico excitatorio (EPSP)
- potencial gradual en la membrana postsináptica que es el resultado de la despolarización y hace que sea más probable que ocurra un potencial de acción
- potencial generador
- potencial gradual de dendritas de una célula unipolar que genera el potencial de acción en el segmento inicial del axón de esa célula
- Proteína G
- guanosina trifosfato (GTP) hidrolasa que se mueve físicamente de la proteína receptora a la proteína efectora para activar esta última
- Potencial postsináptico inhibitorio (IPSP)
- potencial gradual en la membrana postsináptica que es el resultado de la hiperpolarización y hace que un potencial de acción sea menos probable que ocurra
- receptor metabotrópico
- receptor neurotransmisor que involucra un complejo de proteínas que causan cambios metabólicos en una célula
- receptor muscarínico
- tipo de proteína receptora de acetilcolina que se caracteriza por unirse también a muscarina y es un receptor metabotrópico
- neuropéptido
- tipo de neurotransmisor que incluye moléculas de proteína y cadenas más cortas de aminoácidos
- receptor nicotínico
- tipo de proteína receptora de acetilcolina que se caracteriza por unirse también a la nicotina y es un receptor ionotrópico
- Potencial postsináptico (PSP)
- potencial gradual en la membrana postsináptica causado por la unión de neurotransmisores a receptores proteicos
- potencial del receptor
- potencial gradual en una célula sensorial especializada que causa directamente la liberación de neurotransmisores sin un potencial de acción intermedio
- suma espacial
- combinación de potenciales graduados a través de la membrana celular neuronal causada por señales de elementos presinápticos separados que se suman para iniciar un potencial de acción
- summate
- sumar juntos, como en el cambio acumulativo en los potenciales postsinápticos hacia alcanzar el umbral en la membrana, ya sea a través de un lapso de la membrana o durante una cierta cantidad de tiempo
- hendidura sináptica
- pequeña brecha entre células en una sinapsis química donde el neurotransmisor se difunde del elemento presináptico al elemento postsináptico
- suma temporal
- combinación de potenciales graduados en la misma ubicación en una neurona dando como resultado una señal fuerte de una entrada