
14.1: Percepción Sensorial
- Page ID
- 122951

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Describir diferentes tipos de receptores sensoriales
- Describir las estructuras responsables de los sentidos especiales del gusto, el olfato, el oído, el equilibrio y la visión
- Distinguir cómo se transducen los diferentes gustos
- Describir los medios de mecanorecepción para la audición y el equilibrio
- Enumere las estructuras de soporte alrededor del ojo y describa la estructura del globo ocular
- Describir los procesos de fototransducción
Un papel importante de los receptores sensoriales es ayudarnos a aprender sobre el entorno que nos rodea, o sobre el estado de nuestro entorno interno. Los estímulos de diversas fuentes, y de diferentes tipos, son recibidos y transformados en las señales electroquímicas del sistema nervioso. Esto ocurre cuando un estímulo cambia el potencial de membrana celular de una neurona sensorial. El estímulo hace que la célula sensorial produzca un potencial de acción que se transmite al sistema nervioso central (SNC), donde se integra con otra información sensorial, o a veces funciones cognitivas superiores, para convertirse en una percepción consciente de ese estímulo. La integración central puede conducir entonces a una respuesta motora.
Describir la función sensorial con el término sensación o percepción es una distinción deliberada. La sensación es la activación de las células receptoras sensoriales a nivel del estímulo. La percepción es el procesamiento central de los estímulos sensoriales en un patrón significativo. La percepción depende de la sensación, pero no todas las sensaciones son percibidas. Los receptores son las células o estructuras que detectan sensaciones. Una célula receptora es cambiada directamente por un estímulo. Un receptor de proteína transmembrana es una proteína en la membrana celular que media un cambio fisiológico en una neurona, más a menudo a través de la apertura de canales iónicos o cambios en los procesos de señalización celular. Los receptores transmembrana son activados por sustancias químicas llamadas ligandos. Por ejemplo, una molécula en los alimentos puede servir como ligando para los receptores gustativos. Otras proteínas transmembrana, que no se denominan con precisión receptores, son sensibles a los cambios mecánicos o térmicos. Los cambios físicos en estas proteínas aumentan el flujo de iones a través de la membrana y pueden generar un potencial de acción o un potencial gradual en las neuronas sensoriales.
Receptores Sensoriales
Los estímulos en el ambiente activan células receptoras especializadas en el sistema nervioso periférico. Diferentes tipos de estímulos son detectados por diferentes tipos de células receptoras. Las células receptoras pueden clasificarse en tipos sobre la base de tres criterios diferentes: tipo de célula, posición y función. Los receptores pueden clasificarse estructuralmente en función del tipo celular y su posición en relación con los estímulos que perciben. También se pueden clasificar funcionalmente sobre la base de la transducción de estímulos, o cómo el estímulo mecánico, la luz o el químico cambiaron el potencial de la membrana celular.
Tipos de receptores estructurales
Las células que interpretan información sobre el ambiente pueden ser (1) una neurona que tiene una terminación nerviosa libre, con dendritas incrustadas en tejido que recibirían una sensación; (2) una neurona que tiene una terminación encapsulada en la que las terminaciones nerviosas sensoriales están encapsuladas en tejido conectivo que potencia su sensibilidad; o (3) una célula receptora especializada, que tiene distintos componentes estructurales que interpretan un tipo específico de estímulo (Figura\(\PageIndex{1}\)). Los receptores de dolor y temperatura en la dermis de la piel son ejemplos de neuronas que tienen terminaciones nerviosas libres. También se localizan en la dermis de la piel los corpúsculos lamelados, neuronas con terminaciones nerviosas encapsuladas que responden a la presión y al tacto. Las células en la retina que responden a estímulos de luz son un ejemplo de un receptor especializado, un fotorreceptor.
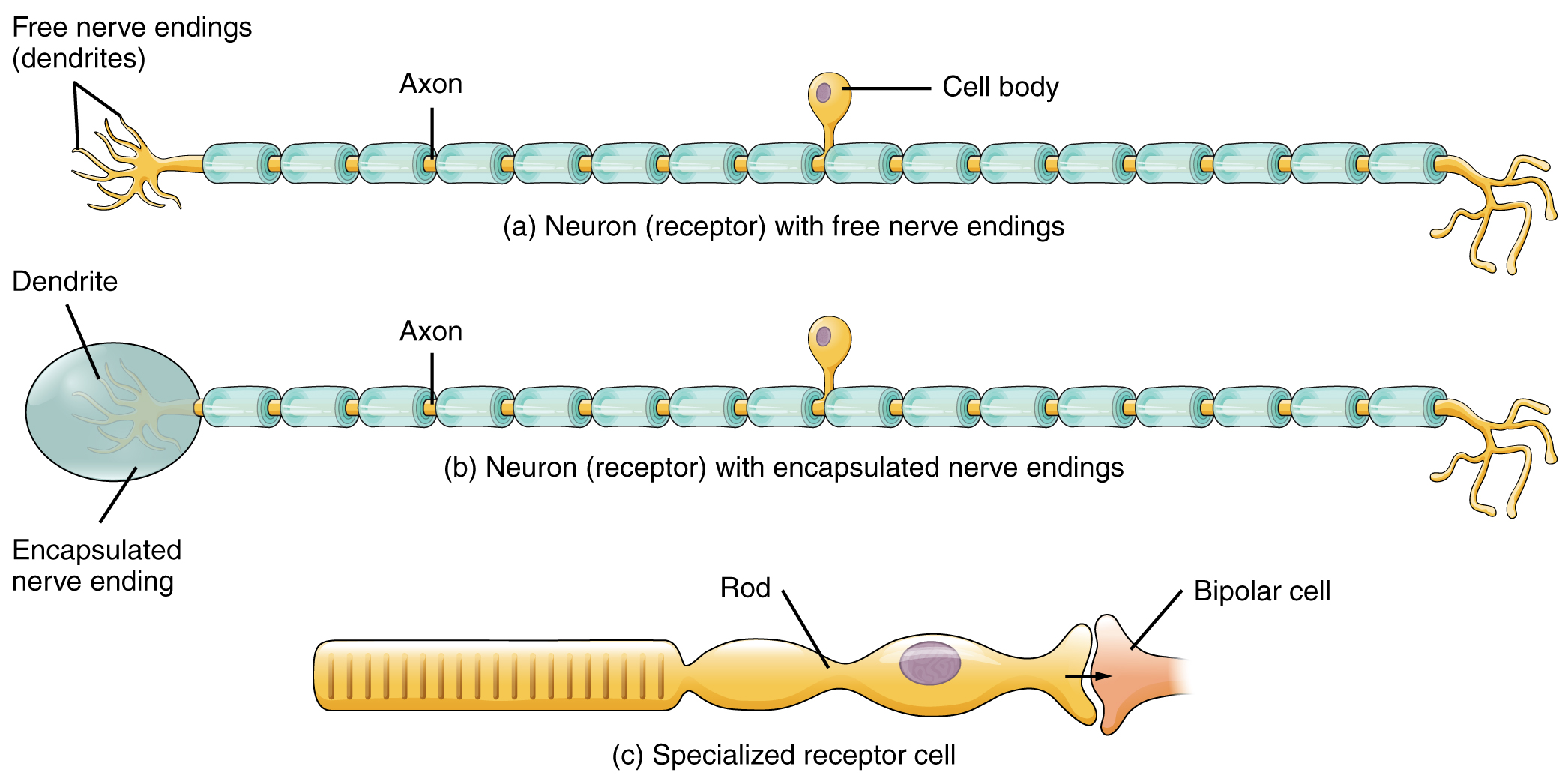
Figura\(\PageIndex{1}\): Clasificación de receptores por tipo celular. Los tipos de células receptoras pueden clasificarse en función de su estructura. Las neuronas sensoriales pueden tener (a) terminaciones nerviosas libres o (b) terminaciones encapsuladas. Los fotorreceptores en los ojos, como los bastoncillos, son ejemplos de (c) células receptoras especializadas. Estas células liberan neurotransmisores sobre una célula bipolar, que luego sinapsis con las neuronas del nervio óptico.
Otra forma en que los receptores pueden clasificarse es en función de su ubicación relativa a los estímulos. Un exteroceptor es un receptor que se localiza cerca de un estímulo en el ambiente externo, como los receptores somatosensoriales que se localizan en la piel. Un interoceptor es aquel que interpreta estímulos de órganos y tejidos internos, como los receptores que perciben el aumento de la presión arterial en la aorta o el seno carotídeo. Finalmente, un propioceptor es un receptor ubicado cerca de una parte móvil del cuerpo, como un músculo, que interpreta las posiciones de los tejidos a medida que se mueven.
Tipos de receptores funcionales
Una tercera clasificación de los receptores es por cómo el receptor transduce estímulos en cambios de potencial de membrana. Los estímulos son de tres tipos generales. Algunos estímulos son iones y macromoléculas que afectan a las proteínas receptoras transmembrana cuando estos químicos se difunden a través de la membrana celular. Algunos estímulos son variaciones físicas en el ambiente que afectan los potenciales de la membrana celular receptora. Otros estímulos incluyen la radiación electromagnética de la luz visible. Para los humanos, la única energía electromagnética que perciben nuestros ojos es la luz visible. Algunos otros organismos tienen receptores que carecen los humanos, como los sensores de calor de las serpientes, los sensores de luz ultravioleta de las abejas o los receptores magnéticos en las aves migratorias.
Las células receptoras pueden clasificarse adicionalmente en función del tipo de estímulos que transducen. Los estímulos químicos pueden ser interpretados por un quimiorreceptor que interpreta estímulos químicos, como el sabor u olor de un objeto. Los osmorreceptores responden a las concentraciones de solutos de los fluidos corporales. Adicionalmente, el dolor es principalmente un sentido químico que interpreta la presencia de sustancias químicas del daño tisular, o estímulos intensos similares, a través de un nociceptor. Los estímulos físicos, como la presión y la vibración, así como la sensación de sonido y posición corporal (equilibrio), se interpretan a través de un mecanorreceptor. Otro estímulo físico que tiene su propio tipo de receptor es la temperatura, que se detecta a través de un termorreceptor que es sensible a temperaturas por encima (calor) o por debajo (fría) de la temperatura corporal normal.
Modalidades Sensoriales
Pregúntele a cualquiera cuáles son los sentidos y es probable que enumeren los cinco sentidos principales: gusto, olfato, tacto, audición y vista. Sin embargo, estos no son todos los sentidos. La omisión más obvia de esta lista es el equilibrio. Además, lo que se conoce simplemente como tacto se puede subdividir en presión, vibración, estiramiento y posición del folículo piloso, sobre la base del tipo de mecanorreceptores que perciben estas sensaciones táctiles. Otros sentidos pasados por alto incluyen la percepción de la temperatura por termorreceptores y la percepción del dolor por los nociceptores.
Dentro del ámbito de la fisiología, los sentidos pueden clasificarse como generales o específicos. Un sentido general es aquel que se distribuye por todo el cuerpo y tiene células receptoras dentro de las estructuras de otros órganos. Los mecanorreceptores en la piel, los músculos o las paredes de los vasos sanguíneos son ejemplos de este tipo. Los sentidos generales a menudo contribuyen al sentido del tacto, como se describió anteriormente, o a la propiocepción (movimiento corporal) y cinestesia (movimiento corporal), o a un sentido visceral, que es lo más importante para las funciones autonómicas. Un sentido especial es aquel que tiene un órgano específico dedicado a él, a saber, el ojo, el oído interno, la lengua o la nariz.
Cada uno de los sentidos es referido como modalidad sensorial. Modalidad se refiere a la forma en que se codifica la información, que es similar a la idea de transducción. Las principales modalidades sensoriales se pueden describir en función de cómo se transduce cada una. Los sentidos químicos son el gusto y el olfato. El sentido general que suele denominarse tacto incluye la sensación química en forma de nocicepción, o dolor. La presión, la vibración, el estiramiento muscular y el movimiento del cabello por un estímulo externo, son percibidos por los mecanorreceptores. La audición y el equilibrio también son detectados por los mecanorreceptores. Finalmente, la visión implica la activación de fotorreceptores.
Enumerar todas las diferentes modalidades sensoriales, que pueden contar hasta 17, implica separar los cinco sentidos principales en categorías más específicas, o submodalidades, del sentido más amplio. Una modalidad sensorial individual representa la sensación de un tipo específico de estímulo. Por ejemplo, el sentido general del tacto, que se conoce como somatosensación, se puede separar en presión ligera, presión profunda, vibración, picor, dolor, temperatura o movimiento del cabello.
Gustation (Gusto)
Solo existen unas pocas submodalidades reconocidas dentro del sentido del gusto, o gusto. Hasta hace poco, sólo se reconocían cuatro sabores: dulce, salado, agrio y amargo. La investigación a principios del siglo XX condujo al reconocimiento del quinto sabor, el umami, a mediados de la década de 1980. Umami es una palabra japonesa que significa “sabor delicioso”, y a menudo se traduce para significar salado. Investigaciones muy recientes han sugerido que también puede haber un sexto sabor por las grasas, o lípidos.
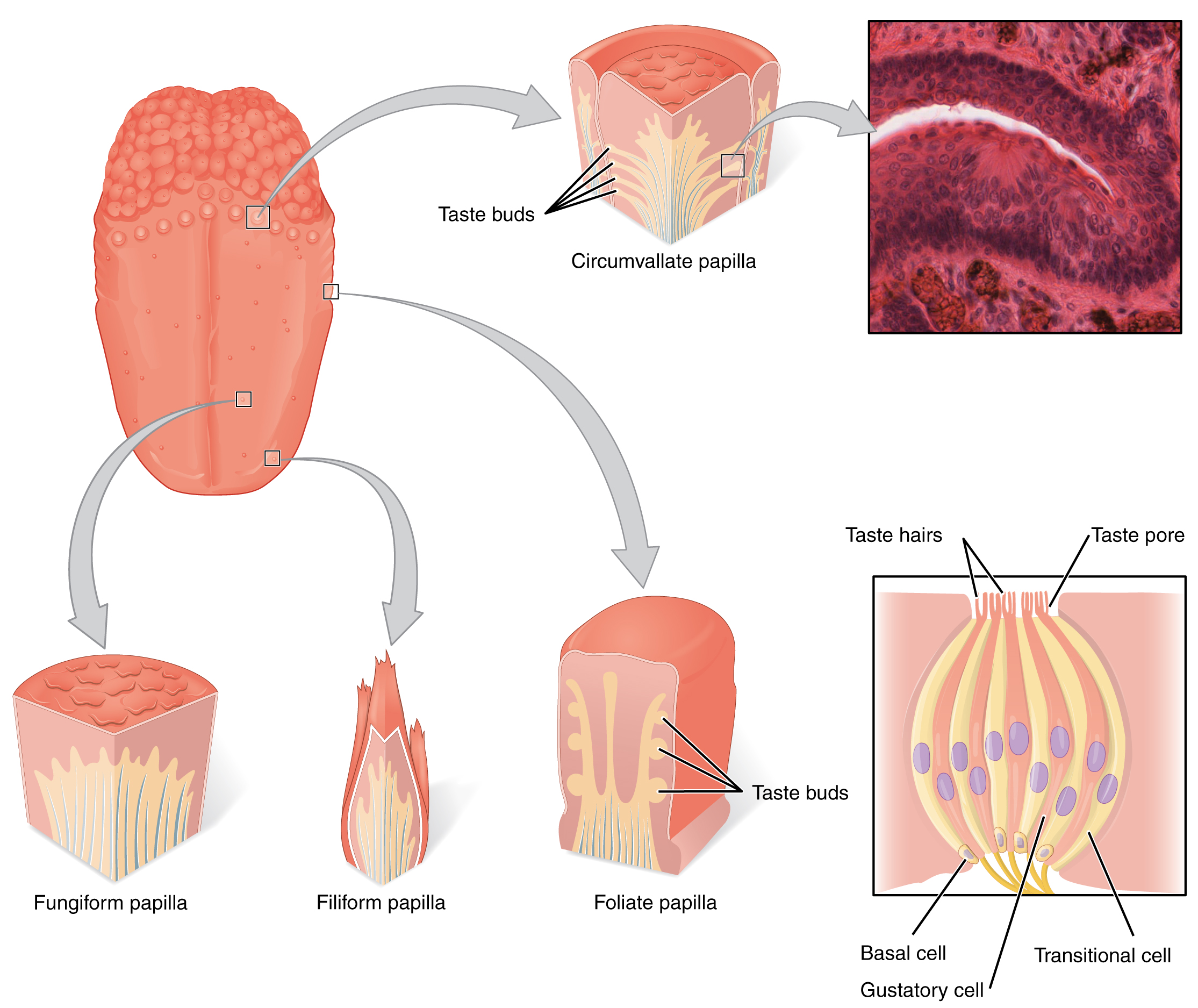
Gustation es el sentido especial asociado a la lengua. La superficie de la lengua, junto con el resto de la cavidad oral, está revestida por un epitelio escamoso estratificado. Las protuberancias elevadas llamadas papilas (singular = papila) contienen las estructuras para la transducción gustativa. Existen cuatro tipos de papilas, según su apariencia (Figura\(\PageIndex{2}\)): circunvaladas, foliadas, filiformes y fungiformes. Dentro de la estructura de las papilas se encuentran las papilas gustativas que contienen células receptoras gustativas especializadas para la transducción de estímulos gustativos. Estas células receptoras son sensibles a los químicos contenidos dentro de los alimentos que se ingieren, y liberan neurotransmisores en función de la cantidad del químico en los alimentos. Los neurotransmisores de las células gustativas pueden activar neuronas sensoriales en los nervios facial, glosofaríngeo y craneal vago.
El sabor salado es simplemente la percepción de iones de sodio (Na +) en la saliva. Cuando comes algo salado, los cristales de sal se disocian en los iones componentes Na + y Cl —, que se disuelven en la saliva de tu boca. La concentración de Na + se vuelve alta fuera de las células gustativas, creando un fuerte gradiente de concentración que impulsa la difusión del ion hacia las células. La entrada de Na + en estas células da como resultado la despolarización de la membrana celular y la generación de un potencial receptor.
El sabor agrio es la percepción de la concentración de H +. Al igual que con los iones de sodio en sabores salados, estos iones de hidrógeno ingresan a la célula y desencadenan la despolarización Los sabores agrios son, esencialmente, la percepción de los ácidos en nuestros alimentos. El aumento de las concentraciones de iones de hidrógeno en la saliva (disminución del pH de la saliva) desencadena potenciales graduados progresivamente más fuertes en las células gustativas. Por ejemplo, el jugo de naranja, que contiene ácido cítrico, tendrá un sabor agrio porque tiene un valor de pH de aproximadamente 3. Por supuesto, a menudo se endulza para que el sabor agrio quede enmascarado.
Los dos primeros sabores (salado y agrio) son desencadenados por los cationes Na + y H +. Los otros sabores son el resultado de moléculas de alimentos que se unen a un receptor acoplado a proteína G. Un sistema de transducción de señales de proteína G finalmente conduce a la despolarización de la célula gustativa. El sabor dulce es la sensibilidad de las células gustativas a la presencia de glucosa disuelta en la saliva. Otros monosacáridos como la fructosa, o edulcorantes artificiales como el aspartamo (NutraSweet™), la sacarina o la sucralosa (Splenda™) también activan los receptores dulces. La afinidad por cada una de estas moléculas varía, y algunas tendrán un sabor más dulce que la glucosa porque se unen al receptor acoplado a la proteína G de manera diferente.
El sabor amargo es similar al dulce en que las moléculas de los alimentos se unen a los receptores acoplados a proteínas G. Sin embargo, hay una serie de formas diferentes en las que esto puede suceder porque hay una gran diversidad de moléculas de sabor amargo. Algunas moléculas amargas despolarizan las células gustativas, mientras que otras hiperpolarizan las células gustativas. Asimismo, algunas moléculas amargas aumentan la activación de la proteína G dentro de las células gustativas, mientras que otras moléculas amargas disminuyen la activación de la proteína G. La respuesta específica depende de qué molécula se une al receptor.
Un grupo importante de moléculas de sabor amargo son los alcaloides. Los alcaloides son moléculas que contienen nitrógeno que a menudo tienen un pH básico. Los alcaloides se encuentran comúnmente en productos vegetales de sabor amargo, como el café, el lúpulo (en la cerveza), los taninos (en el vino), el té y la aspirina. Al contener alcaloides tóxicos, la planta es menos susceptible a la infección por microbios y menos atractiva para los herbívoros.
Por lo tanto, la función del sabor amargo puede estar relacionada principalmente con estimular el reflejo nauseoso para evitar ingerir venenos. Debido a esto, muchos alimentos amargos que normalmente se ingieren suelen combinarse con un componente dulce para hacerlos más apetecibles (crema y azúcar en el café, por ejemplo). La mayor concentración de receptores amargos parece estar en la lengua posterior, donde un reflejo nauseoso aún podría escupir alimentos venenosos.
El sabor conocido como umami a menudo se conoce como el sabor salado. Al igual que el dulce y el amargo, se basa en la activación de receptores acoplados a proteínas G por una molécula específica. La molécula que activa este receptor es el aminoácido L-glutamato. Por lo tanto, el sabor umami a menudo se percibe mientras se comen alimentos ricos en proteínas. No es sorprendente que los platillos que contienen carne a menudo se califiquen como salados.
Una vez que las células gustativas son activadas por las moléculas gustativas, liberan neurotransmisores sobre las dendritas de las neuronas sensoriales. Estas neuronas forman parte de los nervios craneales faciales y glosofaríngeos, así como un componente dentro del nervio vago dedicado al reflejo nauseoso. El nervio facial se conecta a las papilas gustativas en el tercio anterior de la lengua. El nervio glosofaríngeo se conecta a las papilas gustativas en los dos tercios posteriores de la lengua. El nervio vago se conecta a las papilas gustativas en el extremo posterior de la lengua, al borde de la faringe, que son más sensibles a estímulos nocivos como el amargor.

Mira este video para conocer a la Dra. Danielle Reed del Monell Chemical Senses Center en Filadelfia, Pensilvania, quien se interesó por la ciencia a temprana edad debido a sus experiencias sensoriales. Reconoció que su sentido del gusto era único comparado con otras personas que conocía. Ahora, estudia las diferencias genéticas entre las personas y sus sensibilidades a los estímulos gustativos. En el video, hay una breve imagen de una persona sacando la lengua, la cual ha sido cubierta con un tinte de color. Es así como el Dr. Reed es capaz de visualizar y contar papilas en la superficie de la lengua. Las personas caen en dos grupos conocidos como “catadores” y “no catadores” en función de la densidad de papilas en su lengua, lo que también indica el número de papilas gustativas. Los no catadores pueden probar los alimentos, pero no son tan sensibles a ciertos gustos, como el amargor. La doctora Reed descubrió que no es catadora, lo que explica por qué percibió la amargura de manera diferente a otras personas que conocía. ¿Eres muy sensible a los gustos? ¿Puedes ver alguna similitud entre los miembros de tu familia?
Olfato
Al igual que el sabor, el sentido del olfato, u olfato, también responde a estímulos químicos. Las neuronas receptoras olfativas se localizan en una pequeña región dentro de la cavidad nasal superior (Figura\(\PageIndex{3}\)). Esta región se conoce como el epitelio olfativo y contiene neuronas sensoriales bipolares. Cada neurona sensorial olfativa tiene dendritas que se extienden desde la superficie apical del epitelio hasta el moco que recubre la cavidad. A medida que las moléculas transportadas en el aire son inhaladas por la nariz, pasan sobre la región epitelial olfativa y se disuelven en el moco. Estas moléculas odorantes se unen a proteínas que las mantienen disueltas en el moco y ayudan a transportarlas a las dendritas olfativas. El complejo odorante-proteína se une a una proteína receptora dentro de la membrana celular de una dendrita olfativa. Estos receptores están acoplados a proteínas G y producirán un potencial de membrana gradual en las neuronas olfativas.
El axón de una neurona olfativa se extiende desde la superficie basal del epitelio, a través de un foramen olfativo en la placa cribriforme del hueso etmoideo, hasta el cerebro. El grupo de axones llamado tracto olfativo se conecta al bulbo olfativo en la superficie ventral del lóbulo frontal. A partir de ahí, los axones se dividieron para viajar a varias regiones cerebrales. Algunos viajan al cerebro, específicamente a la corteza olfativa primaria que se encuentra en las áreas inferior y medial del lóbulo temporal. Otros proyectan estructuras dentro del sistema límbico y del hipotálamo, donde los olores se asocian con la memoria a largo plazo y las respuestas emocionales. Es así como ciertos olores desencadenan recuerdos emocionales, como el olor a comida asociado con el lugar de nacimiento de uno. El olfato es la única modalidad sensorial que no hace sinapsis en el tálamo antes de conectarse a la corteza cerebral. Esta íntima conexión entre el sistema olfativo y la corteza cerebral es una de las razones por las que el olfato puede ser un potente detonante de recuerdos y emociones.
El epitelio nasal, incluyendo las células olfativas, puede ser dañado por sustancias químicas tóxicas transportadas por el aire. Por lo tanto, las neuronas olfativas se reemplazan regularmente dentro del epitelio nasal, después de lo cual los axones de las nuevas neuronas deben encontrar sus conexiones apropiadas en el bulbo olfativo. Estos nuevos axones crecen a lo largo de los axones que ya están en su lugar en el nervio craneal.

Figura\(\PageIndex{3}\): El Sistema Olfativo. a) El sistema olfativo comienza en las estructuras periféricas de la cavidad nasal. (b) Las neuronas receptoras olfativas se encuentran dentro del epitelio olfativo. (c) Los axones de las neuronas receptoras olfativas se proyectan a través de la placa cribriforme del hueso etmoideo y sinapsis con las neuronas del bulbo olfatorio (fuente tisular: simio). LM × 812. (Micrografía proporcionada por la Facultad de Medicina Regentes de la Universidad de Michigan © 2012)
TRASTORNOS DE LA... Sistema olfativo: Anosmia
Un traumatismo contundente en el rostro, como el común en muchos accidentes automovilísticos, puede llevar a la pérdida del nervio olfativo, y posteriormente, a la pérdida del sentido del olfato. Este padecimiento se conoce como anosmia. Cuando el lóbulo frontal del cerebro se mueve en relación con el hueso etmoideo, los axones del tracto olfativo pueden ser cortados. Los luchadores profesionales suelen experimentar anosmia debido a repetidos traumas en la cara y la cabeza. Además, ciertos productos farmacéuticos, como los antibióticos, pueden causar anosmia al matar todas las neuronas olfativas a la vez. Si no hay axones en su lugar dentro del nervio olfativo, entonces los axones de las neuronas olfativas recién formadas no tienen una guía para conducirlos a sus conexiones dentro del bulbo olfativo. También hay causas temporales de anosmia, como las provocadas por respuestas inflamatorias relacionadas con infecciones respiratorias o alergias.
La pérdida del sentido del olfato puede resultar en un sabor suave de los alimentos. Una persona con un sentido del olfato alterado puede requerir niveles adicionales de especias y condimentos para probar los alimentos. La anosmia también puede estar relacionada con algunas presentaciones de depresión leve, ya que la pérdida del disfrute de los alimentos puede llevar a una sensación general de desesperación.
La capacidad de las neuronas olfativas para reemplazarse disminuye con la edad, lo que lleva a la anosmia relacionada con la edad. Esto explica por qué algunas personas mayores salan su comida más que las personas más jóvenes. Sin embargo, este aumento de la ingesta de sodio puede aumentar el volumen sanguíneo y la presión arterial, aumentando el riesgo de enfermedades cardiovasculares en los adultos mayores.
Audición (Audición)
La audición, o audición, es la transducción de ondas sonoras en una señal neural que es posible gracias a las estructuras del oído (Figura\(\PageIndex{4}\)). La estructura grande y carnosa en la cara lateral de la cabeza se conoce como aurícula. Algunas fuentes también se referirán a esta estructura como el pabellón auricular, aunque ese término es más apropiado para una estructura que se puede mover, como el oído externo de un gato. Las curvas en forma de C de la aurícula dirigen las ondas sonoras hacia el canal auditivo. El canal ingresa al cráneo a través del meato auditivo externo del hueso temporal. Al final del canal auditivo se encuentra la membrana timpánica, o tímpano, que vibra después de ser golpeada por las ondas sonoras. La aurícula, el canal auditivo y la membrana timpánica a menudo se conocen como el oído externo. El oído medio consiste en un espacio abarcado por tres pequeños huesos llamados huesecillos. Los tres huesecillos son el malleo, el incus y el estribo, que son nombres latinos que más o menos se traducen en martillo, yunque y estribo. El maleo se une a la membrana timpánica y se articula con el incus. El incus, a su vez, se articula con el estribo. Luego, el estribo se une al oído interno, donde las ondas sonoras se transducirán en una señal neural. El oído medio está conectado a la faringe a través de la trompa de Eustaquio, lo que ayuda a equilibrar la presión del aire a través de la membrana timpánica. El tubo normalmente está cerrado pero se abrirá cuando los músculos de la faringe se contraen durante la deglución o bostezo.

Figura\(\PageIndex{4}\): Estructuras de la Oreja. El oído externo contiene la aurícula, el canal auditivo y la membrana timpánica. El oído medio contiene los huesecillos y está conectado a la faringe por la trompa de Eustaquio. El oído interno contiene la cóclea y el vestíbulo, que son responsables de la audición y el equilibrio, respectivamente.
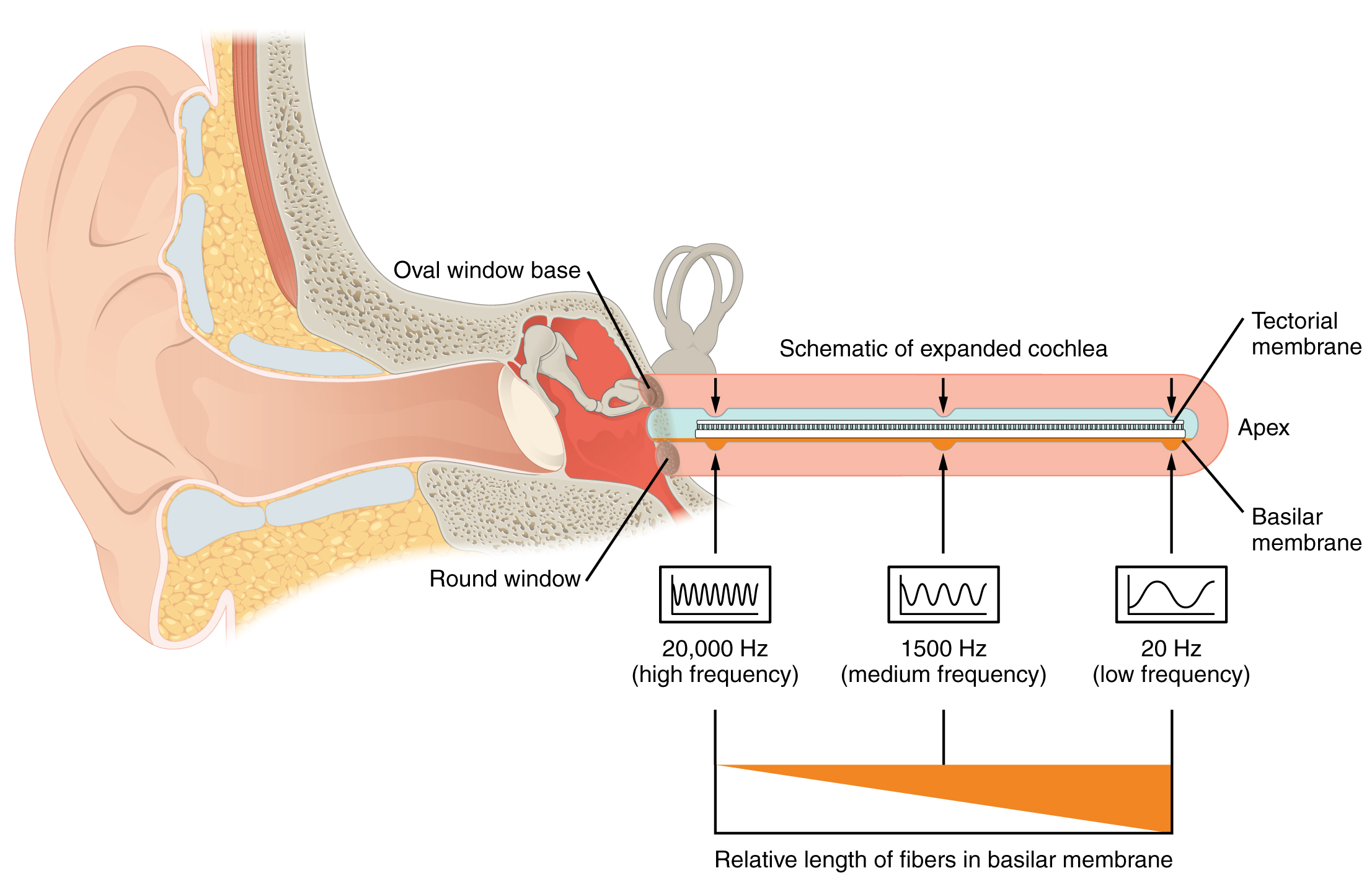
El oído interno a menudo se describe como un laberinto óseo, ya que está compuesto por una serie de canales incrustados dentro del hueso temporal. Cuenta con dos regiones separadas, la cóclea y el vestíbulo, las cuales se encargan de la audición y el equilibrio, respectivamente. Las señales neuronales de estas dos regiones se transmiten al tronco encefálico a través de haces de fibras separados. Sin embargo, estos dos haces distintos viajan juntos desde el oído interno hasta el tronco encefálico como nervio vestibulococlear. El sonido se transduce en señales neuronales dentro de la región coclear del oído interno, que contiene las neuronas sensoriales de los ganglios espirales. Estos ganglios se encuentran dentro de la cóclea en forma de espiral del oído interno. La cóclea se une al estribo a través de la ventana ovalada.
La ventana oval se ubica al inicio de un tubo lleno de líquido dentro de la cóclea llamado scala vestibuli. La scala vestibuli se extiende desde la ventana oval, viajando por encima del conducto coclear, que es la cavidad central de la cóclea que contiene las neuronas transductoras de sonido. En la punta superior de la cóclea, la scala vestibuli se curva sobre la parte superior del conducto coclear. El tubo lleno de líquido, ahora llamado el tímpano de la scala, regresa a la base de la cóclea, esta vez viajando por debajo del conducto coclear. El tímpano de la scala termina en la ventana redonda, que está cubierta por una membrana que contiene el fluido dentro de la scala. A medida que las vibraciones de los huesecillos viajan a través de la ventana ovalada, el fluido de la scala vestibuli y la scala tympani se mueve en un movimiento ondulatorio. La frecuencia de las ondas fluidas coincide con las frecuencias de las ondas sonoras (Figura\(\PageIndex{5}\)). La membrana que cubre la ventana redonda se abultará o se plegará con el movimiento del líquido dentro del tímpano de la scala.
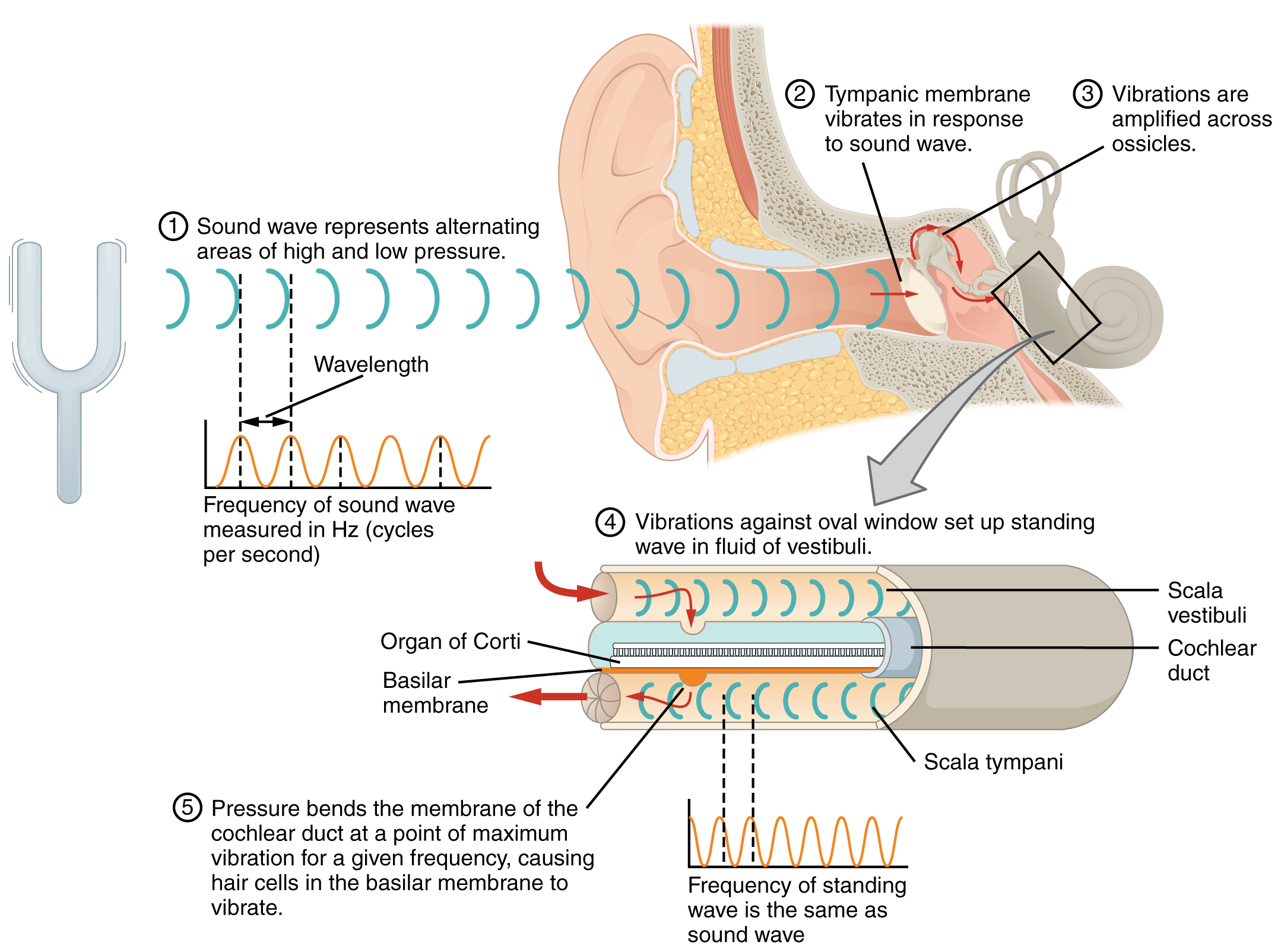
Una vista transversal de la cóclea muestra que la scala vestibuli y la scala tympani discurren a lo largo de ambos lados del conducto coclear (Figura\(\PageIndex{6}\)). El conducto coclear contiene varios órganos de Corti, que tranducen el movimiento de onda de las dos escalas en señales neuronales. Los órganos de Corti se encuentran en la parte superior de la membrana basilar, que es el lado del conducto coclear ubicado entre los órganos de Corti y la scala tympani. A medida que las ondas fluidas se mueven a través de la scala vestibuli y la scala tympani, la membrana basilar se mueve en un punto específico, dependiendo de la frecuencia de las ondas. Las ondas de mayor frecuencia mueven la región de la membrana basilar que está cerca de la base de la cóclea. Las ondas de menor frecuencia mueven la región de la membrana basilar que está cerca de la punta de la cóclea.

Los órganos de Corti contienen células ciliadas, las cuales se llaman así por la estereocilia similar al pelo que se extiende desde las superficies apicales de la célula (Figura\(\PageIndex{7}\)). La estereocilia es una matriz de estructuras similares a microvellosidades dispuestas de la más alta a la más corta. Las fibras proteicas unen los pelos adyacentes dentro de cada matriz, de tal manera que la matriz se doblará en respuesta a los movimientos de la membrana basilar. La estereocilia se extiende desde las células ciliadas hasta la membrana tectorial suprayacente, la cual está unida medialmente al órgano de Corti. Cuando las ondas de presión de la scala mueven la membrana basilar, la membrana tectorial se desliza a través de la estereocilia. Esto dobla la estereocilia hacia o lejos del miembro más alto de cada matriz. Cuando la estereocilia se dobla hacia el miembro más alto de su matriz, la tensión en los anclajes proteicos abre canales iónicos en la membrana celular pilosa. Esto despolarizará la membrana celular pilosa, desencadenando impulsos nerviosos que viajan por las fibras nerviosas aferentes unidas a las células ciliadas. Cuando la estereocilia se dobla hacia el miembro más corto de su matriz, la tensión en las ataduras se afloja y los canales iónicos se cierran. Cuando no hay sonido presente, y la estereocilia se mantiene recta, todavía existe una pequeña cantidad de tensión en las ataduras, manteniendo el potencial de membrana de la célula ciliada ligeramente despolarizado.
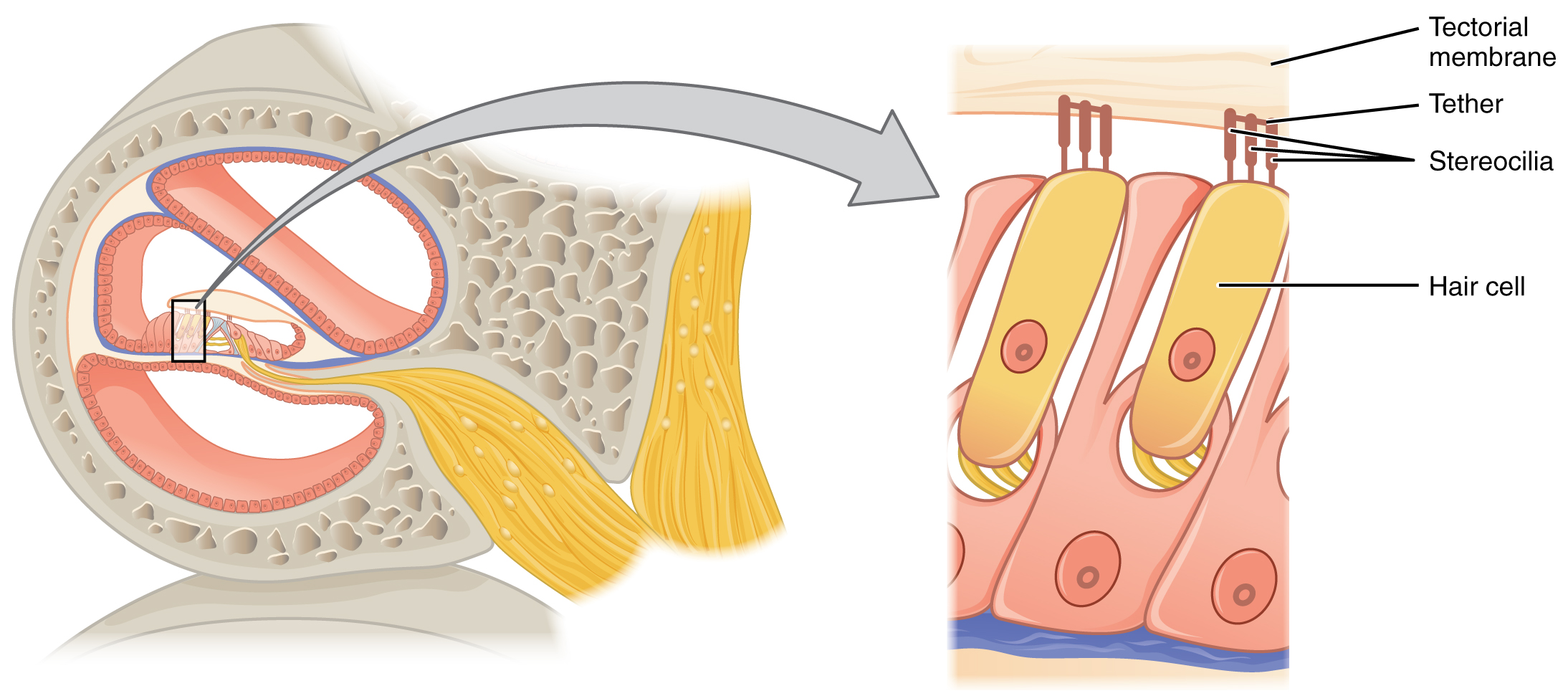


Como se indicó anteriormente, una región dada de la membrana basilar solo se moverá si el sonido entrante está a una frecuencia específica. Debido a que la membrana tectorial solo se mueve donde se mueve la membrana basilar, las células ciliadas en esta región también solo responderán a sonidos de esta frecuencia específica. Por lo tanto, a medida que cambia la frecuencia de un sonido, se activan diferentes células ciliadas a lo largo de la membrana basilar. La cóclea codifica estímulos auditivos para frecuencias entre 20 y 20,000 Hz, que es el rango de sonido que los oídos humanos pueden detectar. La unidad de Hertz mide la frecuencia de las ondas sonoras en términos de ciclos producidos por segundo. Frecuencias tan bajas como 20 Hz son detectadas por las células ciliadas en el ápice, o punta, de la cóclea. Las frecuencias en los rangos más altos de 20 kHz son codificadas por células ciliadas en la base de la cóclea, cerca de las ventanas redondas y ovales (Figura\(\PageIndex{9}\)). Most auditory stimuli contain a mixture of sounds at a variety of frequencies and intensities (represented by the amplitude of the sound wave). The hair cells along the length of the cochlear duct, which are each sensitive to a particular frequency, allow the cochlea to separate auditory stimuli by frequency, just as a prism separates visible light into its component colors.


Equilibrio (Equilibrio)
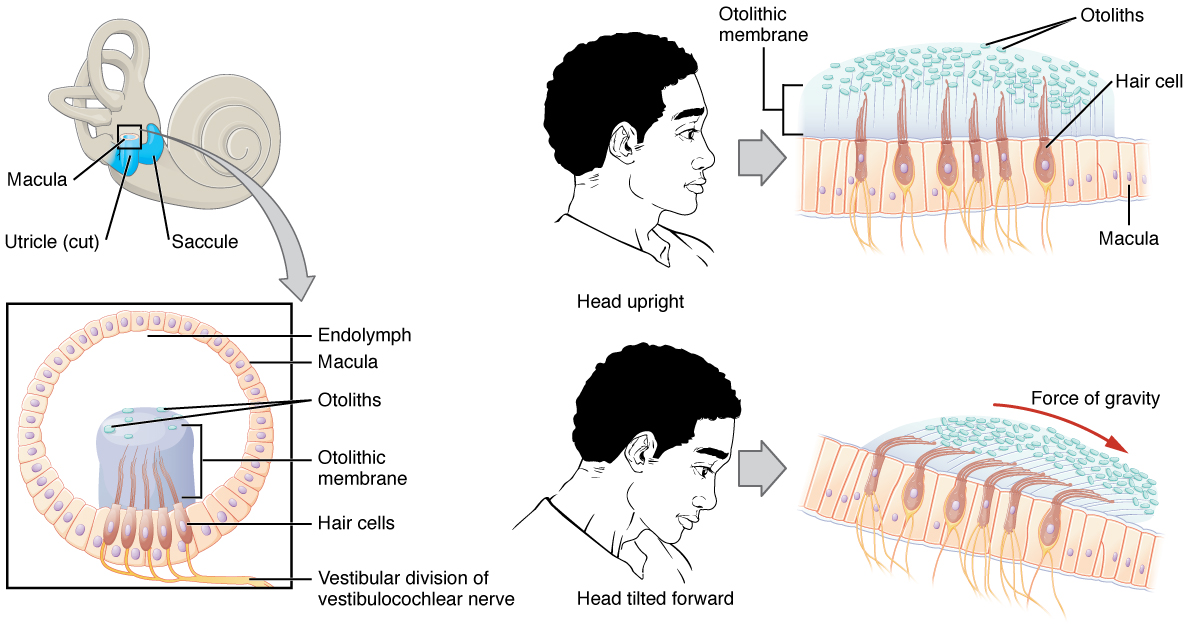
Junto con la audición, el oído interno se encarga de codificar la información sobre el equilibrio, el sentido del equilibrio. Un mecanoreceptor similar, una célula pilosa con estereocilia, detecta la posición de la cabeza, el movimiento de la cabeza y si nuestros cuerpos están en movimiento. Estas células se localizan dentro del vestíbulo del oído interno. La posición de la cabeza es detectada por el utrículo y el sáculo, mientras que el movimiento de la cabeza es detectado por los canales semicirculares. Las señales neuronales generadas en el ganglio vestibular se transmiten a través del nervio vestibulococlear al tronco encefálico y al cerebelo.
El utrículo y el sáculo están compuestos en gran parte de tejido de mácula (plural = maculae). La mácula está compuesta por células ciliadas rodeadas por células de soporte. La estereocilia de las células ciliadas se extiende en un gel viscoso llamado membrana otolítica (Figura\(\PageIndex{10}\)). Encima de la membrana otolítica se encuentra una capa de cristales de carbonato de calcio, llamados otolitos. Los otolitos esencialmente hacen que la membrana otolítica sea pesada en la parte superior. La membrana otolítica se mueve separadamente de la mácula en respuesta a los movimientos de la cabeza. La inclinación de la cabeza hace que la membrana otolítica se deslice sobre la mácula en dirección a la gravedad. La membrana otolítica móvil, a su vez, dobla la esterocilia, provocando que algunas células ciliadas se despolaricen mientras que otras hiperpolarizan. La posición exacta de la cabeza es interpretada por el cerebro basándose en el patrón de despolarización de células pilosas.
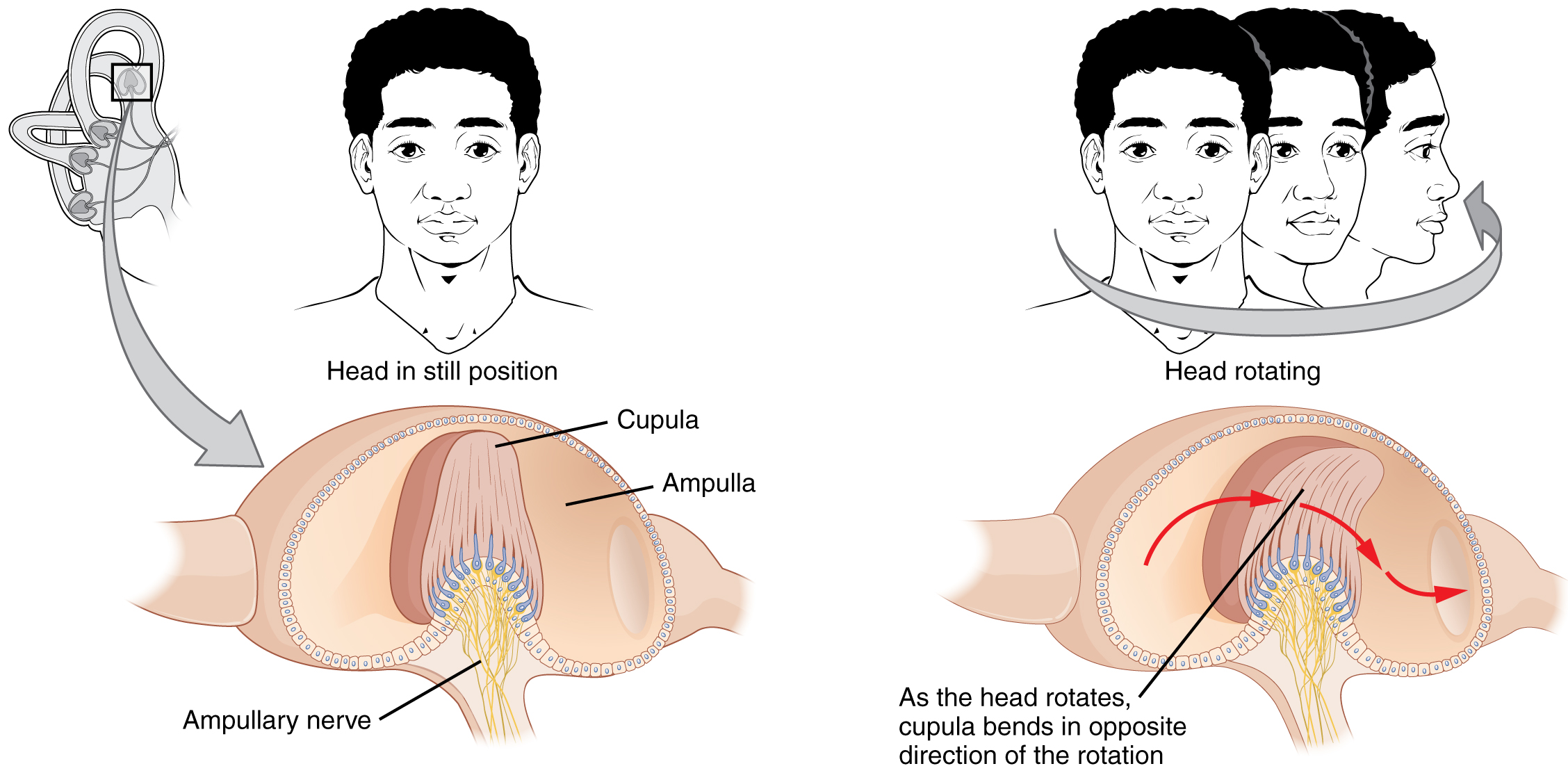
Los canales semicirculares son tres extensiones anulares del vestíbulo. Uno está orientado en el plano horizontal, mientras que los otros dos están orientados en el plano vertical. Los canales verticales anterior y posterior están orientados aproximadamente a 45 grados con respecto al plano sagital (Figura\(\PageIndex{11}\)). La base de cada canal semicircular, donde se encuentra con el vestíbulo, conecta con una región agrandada conocida como la ampolla. La ampolla contiene las células ciliadas que responden al movimiento de rotación, como girar la cabeza mientras se dice “no”. La estereocilia de estas células ciliadas se extiende hacia la cúpula, una membrana que se adhiere a la parte superior de la ampolla. A medida que la cabeza gira en un plano paralelo al canal semicircular, el fluido se desvía, desviando la cúpula en dirección opuesta al movimiento de la cabeza. Los canales semicirculares contienen varias ampullas, algunas orientadas horizontalmente y otras verticalmente. Al comparar los movimientos relativos de las ampollas horizontales y verticales, el sistema vestibular puede detectar la dirección de la mayoría de los movimientos de la cabeza dentro del espacio tridimensional (3-D).
Somatosensación (Tacto)
La somatosensación se considera un sentido general, a diferencia de los sentidos especiales discutidos en esta sección. La somatosensación es el grupo de modalidades sensoriales que se asocian con el tacto, la propiocepción y la interocepción. Estas modalidades incluyen presión, vibración, tacto ligero, cosquillas, picor, temperatura, dolor, propiocepción y cinestesia. Esto significa que sus receptores no están asociados con un órgano especializado, sino que se encuentran diseminados por todo el cuerpo en una variedad de órganos. Muchos de los receptores somatosensoriales se encuentran en la piel, pero los receptores también se encuentran en los músculos, tendones, cápsulas articulares, ligamentos, y en las paredes de los órganos viscerales.
Dos tipos de señales somatosensoriales que son transducidas por terminaciones nerviosas libres son el dolor y la temperatura. Estas dos modalidades utilizan termorreceptores y nociceptores para transducir estímulos de temperatura y dolor, respectivamente. Los receptores de temperatura se estimulan cuando las temperaturas locales difieren de la temperatura corporal. Algunos termorreceptores son sensibles solo al frío y otros solo al calor. La nocicepción es la sensación de estímulos potencialmente dañinos. Los estímulos mecánicos, químicos o térmicos más allá de un umbral establecido provocarán sensaciones dolorosas. Los tejidos estresados o dañados liberan sustancias químicas que activan las proteínas receptoras en los nociceptores. Por ejemplo, la sensación de calor asociada a los alimentos picantes involucra a la capsaicina, la molécula activa en los pimientos picantes. Las moléculas de capsaicina se unen a un canal iónico transmembrana en nociceptores sensibles a temperaturas superiores a 37°C.La dinámica de la unión de la capsaicina con este canal iónico transmembrana es inusual ya que la molécula permanece unida durante mucho tiempo. Debido a esto, disminuirá la capacidad de otros estímulos para provocar sensaciones de dolor a través del nociceptor activado. Por esta razón, la capsaicina se puede utilizar como analgésico tópico, como en productos como Icy Hot™.
Si arrastras tu dedo por una superficie texturizada, la piel de tu dedo vibrará. Estas vibraciones de baja frecuencia son detectadas por mecanorreceptores llamados células de Merkel, también conocidos como mecanorreceptores cutáneos tipo I. Las células de Merkel se localizan en el estrato basal de la epidermis. La presión profunda y la vibración son transducidas por los corpúsculos lamelados (Pacinianos), que son receptores con terminaciones encapsuladas que se encuentran profundamente en la dermis, o tejido subcutáneo. El tacto ligero es transducido por las terminaciones encapsuladas conocidas como corpúsculos táctiles (Meissner). Los folículos también están envueltos en un plexo de terminaciones nerviosas conocido como plexo del folículo piloso. Estas terminaciones nerviosas detectan el movimiento del pelo en la superficie de la piel, como cuando un insecto puede estar caminando a lo largo de la piel. El estiramiento de la piel es transducido por receptores de estiramiento conocidos como corpúsculos bulbosos. Los corpúsculos bulbosos también se conocen como corpúsculos Ruffini, o mecanorreceptores cutáneos tipo II.
Otros receptores somatosensoriales se encuentran en las articulaciones y músculos. Los receptores de estiramiento monitorean el estiramiento de tendones, músculos y los componentes de las articulaciones. Por ejemplo, ¿alguna vez has estirado tus músculos antes o después del ejercicio y has notado que solo puedes estirarte hasta ahora antes de que tus músculos vuelvan a tener un estado menos estirado? Este espasmo es un reflejo que es iniciado por receptores de estiramiento para evitar el desgarro muscular. Dichos receptores de estiramiento también pueden prevenir la sobrecontracción de un músculo. En el tejido muscular esquelético, estos receptores de estiramiento se denominan husillos musculares. Los órganos del tendón de Golgi transducen de manera similar los niveles de estiramiento de los tendones. Los corpúsculos bulbosos también están presentes en las cápsulas articulares, donde miden el estiramiento en los componentes del sistema esquelético dentro de la articulación. Los tipos de terminaciones nerviosas, sus ubicaciones y los estímulos que transducen se presentan en la Tabla\(\PageIndex{1}\).
| Mecanorreceptores de Somatosensación | |||
|---|---|---|---|
| Nombre | Nombre histórico (epónimo) | Ubicación (es) | Estímulos |
| Terminaciones nerviosas libres | * | Dermis, córnea, lengua, cápsulas articulares, órganos viscerales | Dolor, temperatura, deformación mecánica |
| Mecanorreceptores | Discos de Merkel | Unión epidérmica-dérmica, membranas mucosas | Vibración de baja frecuencia (5—15 Hz) |
| Corpúsculo bulboso | Corpúsculo de Ruffini | Dermis, cápsulas articulares | Estiramiento |
| Corpúsculo táctil | Corpúsculo de Meissner | Dermis papilar, especialmente en las yemas de los dedos y los labios | Tacto ligero, vibraciones por debajo de 50 Hz |
| Corpúsculo lamelado | Corpúsculo Paciniano | Dermis profunda, tejido subcutáneo | Presión profunda, vibración de alta frecuencia (alrededor de 250 Hz) |
| Plexo del folículo piloso | * | Envuelto alrededor de folículos pilosos en la dermis | Movimiento del cabello |
| Husillo muscular | * | En línea con las fibras del músculo esquelético | Contracción muscular y estiramiento |
| Órgano de estiramiento tendinoso | Órgano tendinoso de Golgi | En línea con los tendones | Estiramiento del tendón |
Visión
La visión es el sentido especial de la vista que se basa en la transducción de estímulos de luz recibidos a través de los ojos. Los ojos se encuentran dentro de cualquiera de las dos órbitas en el cráneo. Las órbitas óseas rodean los globos oculares, protegiéndolos y anclando los tejidos blandos del ojo (Figura\(\PageIndex{12}\)). Los párpados, con pestañas en sus bordes de ataque, ayudan a proteger el ojo de las abrasiones al bloquear las partículas que pueden caer en la superficie del ojo. La superficie interna de cada párpado es una membrana delgada conocida como conjuntiva palpebral. La conjuntiva se extiende sobre las áreas blancas del ojo (la esclerótica), conectando los párpados con el globo ocular. Las lágrimas son producidas por la glándula lagrimal, ubicada debajo de los bordes laterales de la nariz. Las lágrimas producidas por esta glándula fluyen a través del conducto lagrimal hasta la esquina medial del ojo, donde las lágrimas fluyen sobre la conjuntiva, lavando las partículas extrañas.

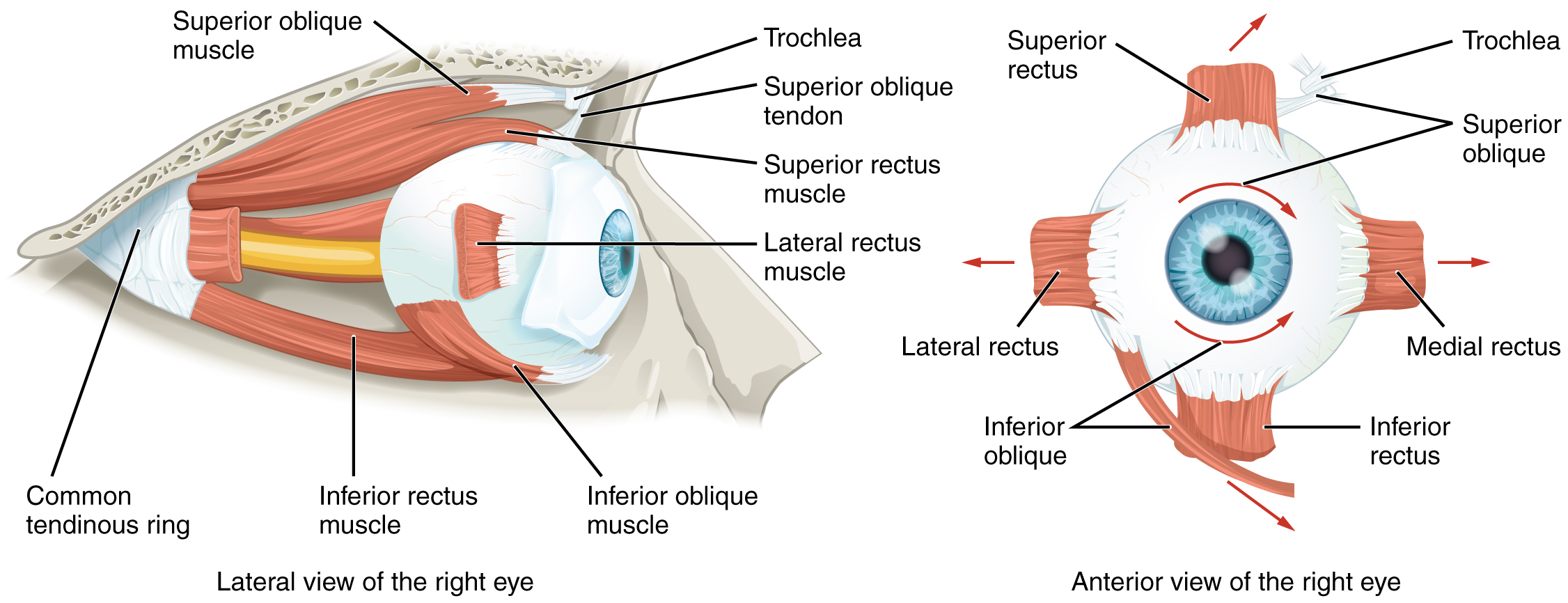
El movimiento del ojo dentro de la órbita se logra mediante la contracción de seis músculos extraoculares que se originan a partir de los huesos de la órbita y se insertan en la superficie del globo ocular (Figura\(\PageIndex{13}\)). Cuatro de los músculos están dispuestos en los puntos cardinales alrededor del ojo y llevan el nombre de esas ubicaciones. Son el recto superior, recto medial, recto inferior y recto lateral. Cuando cada uno de estos músculos se contrae, el ojo a se mueve hacia el músculo que se contrae. Por ejemplo, cuando el recto superior se contrae, el ojo gira para mirar hacia arriba. El oblicuo superior se origina en la órbita posterior, cerca del origen de los cuatro músculos rectos. Sin embargo, el tendón de los músculos oblicuos se enrosca a través de una pieza de cartílago similar a una polea conocida como la troclea. El tendón se inserta oblicuamente en la superficie superior del ojo. El ángulo del tendón a través de la troclea significa que la contracción del oblicuo superior gira el ojo medialmente. El músculo oblicuo inferior se origina en el suelo de la órbita y se inserta en la superficie inferolateral del ojo. Cuando se contrae, gira lateralmente el ojo, en oposición al oblicuo superior. La rotación del ojo por los dos músculos oblicuos es necesaria porque el ojo no está perfectamente alineado en el plano sagital. Cuando el ojo mira hacia arriba o hacia abajo, el ojo también debe girar ligeramente para compensar el recto superior tirando en un ángulo de aproximadamente 20 grados, en lugar de hacerlo recto hacia arriba. Lo mismo ocurre con el recto inferior, que se compensa con la contracción del oblicuo inferior. Un séptimo músculo en la órbita es el elevador palpebrae superioris, el cual se encarga de elevar y retraer el párpado superior, movimiento que suele ocurrir en concierto con la elevación del ojo por el recto superior (ver Figura\(\PageIndex{12}\)).
Los músculos extraoculares están inervados por tres nervios craneales. El recto lateral, que provoca la abducción del ojo, está inervado por el nervio abducens. El oblicuo superior está inervado por el nervio troclear. Todos los demás músculos están inervados por el nervio oculomotor, al igual que el elevador palpebrae superioris. Los núcleos motores de estos nervios craneales se conectan al tronco encefálico, que coordina los movimientos oculares.
El ojo en sí es una esfera hueca compuesta por tres capas de tejido. La capa más externa es la túnica fibrosa, que incluye la esclerótica blanca y la córnea transparente. La esclerótica representa cinco sextas partes de la superficie del ojo, la mayoría de las cuales no es visible, aunque los humanos son únicos comparados con muchas otras especies al tener tanto del “blanco del ojo” visible (Figura\(\PageIndex{14}\)). La córnea transparente cubre la punta anterior del ojo y permite que la luz entre en el ojo. La capa media del ojo es la túnica vascular, que está compuesta principalmente por la coroides, el cuerpo ciliar y el iris. La coroides es una capa de tejido conectivo altamente vascularizado que proporciona un suministro de sangre al globo ocular. La coroides es posterior al cuerpo ciliar, una estructura muscular que está unida al cristalino por fibras de zónulas. Estas dos estructuras doblan la lente, lo que le permite enfocar la luz en la parte posterior del ojo. Superpuesto al cuerpo ciliar, y visible en el ojo anterior, está el iris, la parte coloreada del ojo. El iris es un músculo liso que abre o cierra la pupila, que es el orificio en el centro del ojo que permite la entrada de luz. El iris contrae la pupila en respuesta a la luz brillante y dilata la pupila en respuesta a la luz tenue. La capa más interna del ojo es la túnica neural, o retina, que contiene el tejido nervioso responsable de la fotorrecepción.
El ojo también se divide en dos cavidades: la cavidad anterior y la cavidad posterior. La cavidad anterior es el espacio entre la córnea y el cristalino, incluyendo el iris y el cuerpo ciliar. Se llena de un fluido acuoso llamado humor acuoso. La cavidad posterior es el espacio detrás del cristalino que se extiende hasta el lado posterior del globo ocular interior, donde se encuentra la retina. La cavidad posterior se llena con un líquido más viscoso llamado humor vítreo.
La retina está compuesta por varias capas y contiene células especializadas para el procesamiento inicial de estímulos visuales. Los fotorreceptores (bastones y conos) cambian su potencial de membrana cuando son estimulados por la energía de la luz. El cambio en el potencial de membrana altera la cantidad de neurotransmisor que las células fotorreceptoras liberan sobre las células bipolares en la capa sináptica externa. Es la célula bipolar en la retina la que conecta un fotorreceptor a una célula ganglionar retiniana (RGC) en la capa sináptica interna. Allí, las células amacrinas contribuyen adicionalmente al procesamiento de la retina antes de que el RGC produzca un potencial de acción. Los axones de las RGC, que se encuentran en la capa más interna de la retina, se recogen en el disco óptico y dejan el ojo como nervio óptico (ver Figura\(\PageIndex{14}\)). Debido a que estos axones pasan a través de la retina, no hay fotorreceptores en la parte posterior del ojo, donde comienza el nervio óptico. Esto crea un “punto ciego” en la retina, y un punto ciego correspondiente en nuestro campo visual.

Tenga en cuenta que los fotorreceptores en la retina (bastones y conos) se encuentran detrás de los axones, RGC, células bipolares y vasos sanguíneos retinianos. Una cantidad significativa de luz es absorbida por estas estructuras antes de que la luz llegue a las células fotorreceptoras. Sin embargo, en el centro exacto de la retina hay una pequeña área conocida como la fóvea. En la fóvea, la retina carece de las células de soporte y los vasos sanguíneos, y solo contiene fotorreceptores. Por lo tanto, la agudeza visual, o la nitidez de la visión, es mayor en la fóvea. Esto se debe a que la fóvea es donde la menor cantidad de luz entrante es absorbida por otras estructuras retinianas (ver Figura\(\PageIndex{14}\)). A medida que uno se mueve en cualquier dirección desde este punto central de la retina, la agudeza visual disminuye significativamente. Además, cada célula fotorreceptora de la fóvea está conectada a un solo RGC. Por lo tanto, este RGC no tiene que integrar entradas de múltiples fotorreceptores, lo que reduce la precisión de la transducción visual. Hacia los bordes de la retina, varios fotorreceptores convergen en las RGC (a través de las células bipolares) hasta una proporción de 50 a 1. La diferencia en la agudeza visual entre la fóvea y la retina periférica se evidencia fácilmente al mirar directamente una palabra en medio de este párrafo. El estímulo visual en medio del campo de visión cae sobre la fóvea y se encuentra en el foco más agudo. Sin apartar los ojos de esa palabra, observe que las palabras al principio o al final del párrafo no están enfocadas. Las imágenes en tu visión periférica son enfocadas por la retina periférica, y tienen bordes vagos, borrosos y palabras que no están tan claramente identificadas. Como resultado, gran parte de la función neural de los ojos se ocupa de mover los ojos y la cabeza para que importantes estímulos visuales se centren en la fóvea.
La luz que cae sobre la retina provoca cambios químicos en las moléculas pigmentarias en los fotorreceptores, lo que finalmente conduce a un cambio en la actividad de los RGC. Las células fotorreceptoras tienen dos partes, el segmento interno y el segmento externo (Figura\(\PageIndex{15}\)). El segmento interno contiene el núcleo y otros orgánulos comunes de una célula, mientras que el segmento externo es una región especializada en la que tiene lugar la fotorrecepción. Hay dos tipos de fotorreceptores, varillas y conos, que difieren en la forma de su segmento exterior. Los segmentos externos en forma de varilla del fotorreceptor contienen una pila de discos unidos a membrana que contienen el pigmento fotosensible rodopsina. Los segmentos externos en forma de cono del fotorreceptor de cono contienen sus pigmentos fotosensibles en las incrustaciones de la membrana celular. Hay tres fotopigmentos de cono, llamados opsinas, cada uno de los cuales es sensible a una determinada longitud de onda de luz. La longitud de onda de la luz visible determina su color. Los pigmentos en los ojos humanos están especializados en percibir tres colores primarios diferentes: rojo, verde y azul.

A nivel molecular, los estímulos visuales provocan cambios en la molécula fotopigmentaria que conducen a cambios en el potencial de membrana de la célula fotorreceptora. Una sola unidad de luz se llama fotón, el cual se describe en la física como un paquete de energía con propiedades tanto de una partícula como de una onda. La energía de un fotón está representada por su longitud de onda, correspondiendo cada longitud de onda de luz visible a un color particular. La luz visible es radiación electromagnética con una longitud de onda entre 380 y 720 nm. Las longitudes de onda de radiación electromagnética mayores a 720 nm caen en el rango infrarrojo, mientras que las longitudes de onda más cortas que 380 nm caen en el rango ultravioleta. La luz con una longitud de onda de 380 nm es azul mientras que la luz con una longitud de onda de 720 nm es roja oscura. Todos los demás colores caen entre rojo y azul en varios puntos a lo largo de la escala de longitud de onda.
Los pigmentos opsina son en realidad proteínas transmembrana que contienen un cofactor conocido como retinal. La retina es una molécula de hidrocarburo relacionada con la vitamina A. Cuando un fotón golpea la retina, la larga cadena hidrocarbonada de la molécula se altera bioquímicamente. Específicamente, los fotones hacen que algunos de los carbonos de doble enlace dentro de la cadena cambien de una conformación cis a una trans. Este proceso se llama fotoisomerización. Antes de interactuar con un fotón, los carbonos de doble enlace flexibles de la retina están en la conformación cis. Esta molécula es referida como 11-cis-retinal. Un fotón que interactúa con la molécula hace que los carbonos flexibles de doble enlace cambien a la conformación trans, formando todo-trans-retinal, que tiene una cadena hidrocarbonada lineal (Figura\(\PageIndex{16}\)).
El cambio de forma de la retina en los fotorreceptores inicia la transducción visual en la retina. La activación de las proteínas retinianas y opsina resulta en la activación de una proteína G. La proteína G cambia el potencial de membrana de la célula fotorreceptora, que luego libera menos neurotransmisor en la capa sináptica externa de la retina. Hasta que la molécula retiniana vuelva a cambiar a la forma 11-cis-retiniana, la opsina no puede responder a la energía de la luz, que se llama blanqueamiento. Cuando se blanquea un gran grupo de fotopigmentos, la retina enviará información como si se estuviera percibiendo información visual opuesta. Después de un brillante destello de luz, las imágenes posteriores suelen verse en negativo. La fotoisomerización es revertida por una serie de cambios enzimáticos para que la retina responda a más energía lumínica.
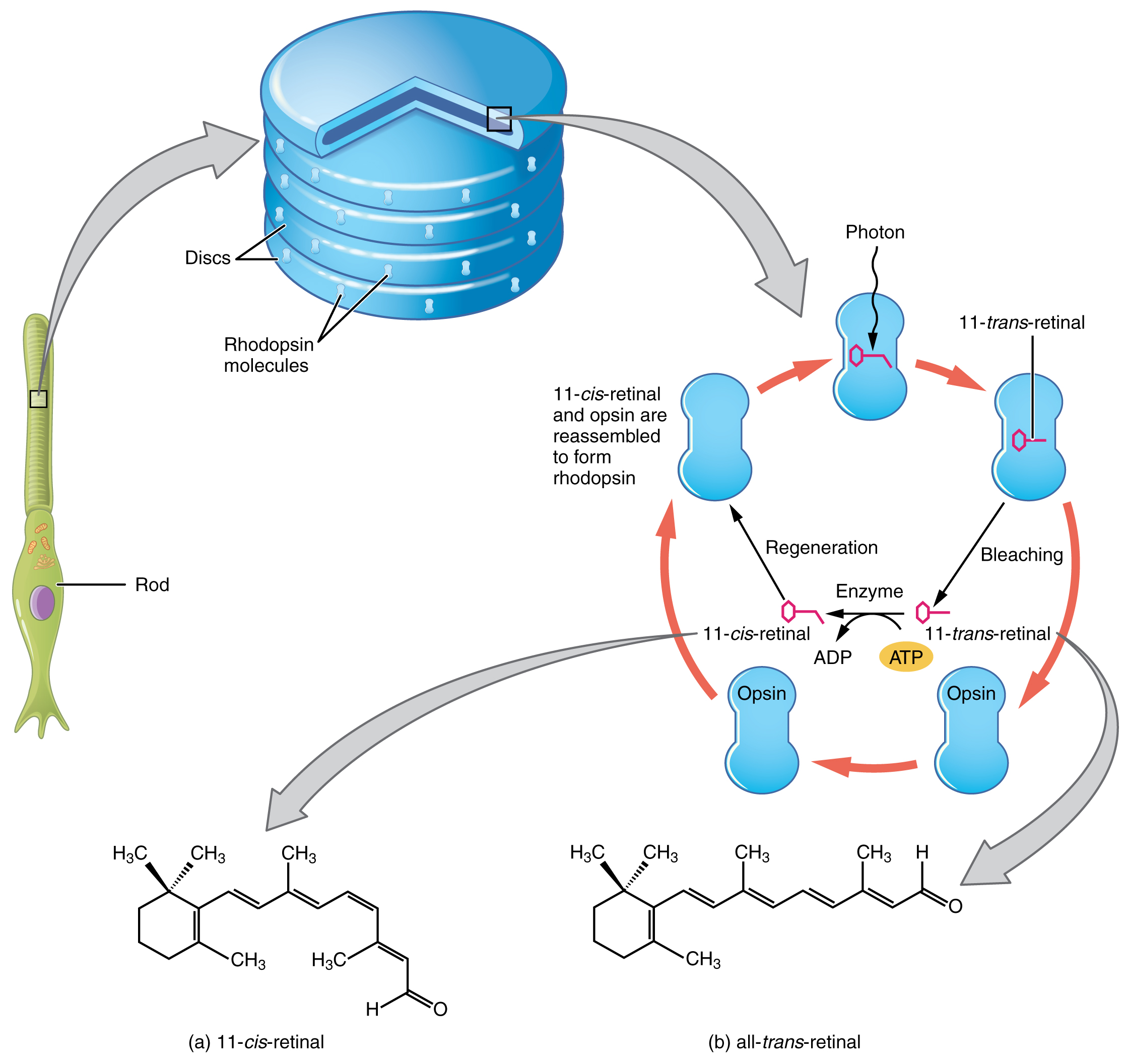
Las opsinas son sensibles a longitudes de onda limitadas de luz. La rodopsina, el fotopigmento en varillas, es más sensible a la luz a una longitud de onda de 498 nm. Las opsinas de tres colores tienen sensibilidades pico de 564 nm, 534 nm y 420 nm correspondientes aproximadamente a los colores primarios de rojo, verde y azul (Figura\(\PageIndex{17}\)). La absorbancia de la rodopsina en las varillas es mucho más sensible que en las opsinas cónicas; específicamente, las varillas son sensibles a la visión en condiciones de poca luz, y los conos son sensibles a condiciones más brillantes. En la luz solar normal, la rodopsina se blanqueará constantemente mientras los conos estén activos. En una habitación oscura, no hay suficiente luz para activar las opsinas de cono, y la visión depende completamente de las varillas. Las varillas son tan sensibles a la luz que un solo fotón puede resultar en un potencial de acción del RGC correspondiente de una varilla.
Los tres tipos de opsinas de cono, al ser sensibles a diferentes longitudes de onda de luz, nos proporcionan visión de color. Al comparar la actividad de los tres conos diferentes, el cerebro puede extraer información de color a partir de estímulos visuales. Por ejemplo, una luz azul brillante que tenga una longitud de onda de aproximadamente 450 nm activaría los conos “rojos” mínimamente, los conos “verdes” marginalmente, y los conos “azules” predominantemente. La activación relativa de los tres conos diferentes es calculada por el cerebro, que percibe el color como azul. Sin embargo, los conos no pueden reaccionar a la luz de baja intensidad y las varillas no perciben el color de la luz. Por lo tanto, nuestra visión con poca luz está, en esencia, en escala de grises. Es decir, en una habitación oscura, todo aparece como una tonalidad de gris. Si piensas que puedes ver colores en la oscuridad, lo más probable es que sea porque tu cerebro sabe de qué color es algo y está confiando en ese recuerdo.


Nervios Sensoriales
Una vez que cualquier célula sensorial transduce un estímulo en un impulso nervioso, ese impulso tiene que viajar a lo largo de los axones para llegar al SNC. En muchos de los sentidos especiales, los axones que salen de los receptores sensoriales tienen una disposición topográfica, lo que significa que la ubicación del receptor sensorial se relaciona con la ubicación del axón en el nervio. Por ejemplo, en la retina, los axones de las RGC en la fóvea se localizan en el centro del nervio óptico, donde están rodeados por axones de las RGC más periféricas.
Nervios Espinales
Generalmente, los nervios espinales contienen axones aferentes de receptores sensoriales en la periferia, como de la piel, mezclados con axones eferentes que viajan a los músculos u otros órganos efectores. A medida que el nervio espinal se acerca a la médula espinal, se divide en raíces dorsales y ventrales. La raíz dorsal contiene solo los axones de las neuronas sensoriales, mientras que las raíces ventrales contienen solo los axones de las neuronas motoras. Algunas de las ramas sinapsificarán con neuronas locales en el ganglio de la raíz dorsal, el cuerno posterior (dorsal), o incluso el cuerno anterior (ventral), al nivel de la médula espinal donde ingresan. Otras ramas recorrerán una corta distancia arriba o abajo de la columna vertebral para interactuar con las neuronas en otros niveles de la médula espinal. Una rama también puede convertirse en la columna posterior (dorsal) de la materia blanca para conectarse con el cerebro. Por conveniencia, utilizaremos los términos ventral y dorsal en referencia a las estructuras dentro de la médula espinal que forman parte de estas vías. Esto ayudará a subrayar las relaciones entre los diferentes componentes. Por lo general, los sistemas nerviosos espinales que se conectan al cerebro son contralaterales, en que el lado derecho del cuerpo está conectado al lado izquierdo del cerebro y el lado izquierdo del cuerpo al lado derecho del cerebro.
Nervios craneales
Los nervios craneales transmiten información sensorial específica desde la cabeza y el cuello directamente al cerebro. Para las sensaciones debajo del cuello, el lado derecho del cuerpo está conectado al lado izquierdo del cerebro y el lado izquierdo del cuerpo al lado derecho del cerebro. Mientras que la información espinal es contralateral, los sistemas nerviosos craneales son en su mayoría ipsilaterales, lo que significa que un nervio craneal en el lado derecho de la cabeza está conectado al lado derecho del cerebro. Algunos nervios craneales contienen solo axones sensoriales, como los nervios olfativos, ópticos y vestibulococleares. Otros nervios craneales contienen axones tanto sensoriales como motores, incluidos los nervios trigémino, facial, glosofaríngeo y vago (sin embargo, el nervio vago no está asociado con el sistema nervioso somático). Los sentidos generales de la somatosensación para el rostro viajan a través del sistema trigémino.
Revisión del Capítulo
Los sentidos son olfato (olfato), gusto (gusto), somatosensación (sensaciones asociadas con la piel y el cuerpo), audición (audición), equilibrio (equilibrio) y visión. A excepción de la somatosensación, esta lista representa los sentidos especiales, o aquellos sistemas del cuerpo que están asociados con órganos específicos como la lengua o el ojo. La somatosensación pertenece a los sentidos generales, que son aquellas estructuras sensoriales que se distribuyen por todo el cuerpo y en las paredes de diversos órganos. Los sentidos especiales son todos principalmente parte del sistema nervioso somático en el sentido de que son percibidos conscientemente a través de procesos cerebrales, aunque algunos sentidos especiales contribuyen a la función autonómica. Los sentidos generales se pueden dividir en somatosensación, que comúnmente se considera táctil, pero incluye táctil, presión, vibración, temperatura y percepción del dolor. Los sentidos generales también incluyen los sentidos viscerales, que están separados de la función del sistema nervioso somático en que normalmente no se elevan al nivel de percepción consciente.
Las células que transducen estímulos sensoriales en las señales electroquímicas del sistema nervioso se clasifican en función de los aspectos estructurales o funcionales de las células. Las clasificaciones estructurales se basan en la anatomía de la célula que interactúa con el estímulo (terminaciones nerviosas libres, terminaciones encapsuladas o célula receptora especializada), o donde la célula se encuentra en relación con el estímulo (interoceptor, exteroceptor, propioceptor). En tercer lugar, la clasificación funcional se basa en cómo la célula transduce el estímulo en una señal neural. Los quimiorreceptores responden a estímulos químicos y son la base para el olfato y la gustación. Los quimiorreceptores están relacionados con los osmorreceptores y nociceptores para el equilibrio de líquidos y la recepción del dolor, respectivamente. Los mecanorreceptores responden a estímulos mecánicos y son la base para la mayoría de los aspectos de la somatosensación, además de ser la base de la audición y el equilibrio en el oído interno. Los termorreceptores son sensibles a los cambios de temperatura y los fotorreceptores son sensibles a la energía lumínica.
Los nervios que transmiten información sensorial desde la periferia al SNC son nervios espinales, conectados a la médula espinal, o nervios craneales, conectados al cerebro. Los nervios espinales tienen poblaciones mixtas de fibras; algunas son fibras motoras y otras sensoriales. Las fibras sensoriales se conectan a la médula espinal a través de la raíz dorsal, que está unida al ganglio de la raíz dorsal. La información sensorial del cuerpo que se transmite a través de los nervios espinales se proyectará hacia el lado opuesto del cerebro para ser procesada por la corteza cerebral. Los nervios craneales pueden ser fibras estrictamente sensoriales, como los nervios olfativos, ópticos y vestibulococleares, o nervios sensoriales y motores mixtos, como los nervios trigémino, facial, glosofaríngeo y vago. Los nervios craneales están conectados al mismo lado del cerebro del que se origina la información sensorial.
Preguntas de Enlace Interactivo
Mira este video para conocer a la Dra. Danielle Reed del Monell Chemical Senses Center en Filadelfia, PA, quien se interesó por la ciencia a temprana edad debido a sus experiencias sensoriales. Reconoció que su sentido del gusto era único comparado con otras personas que conocía. Ahora, estudia las diferencias genéticas entre las personas y sus sensibilidades a los estímulos gustativos. En el video, hay una breve imagen de una persona sacando la lengua, la cual ha sido cubierta con un tinte de color. Es así como el Dr. Reed es capaz de visualizar y contar papilas en la superficie de la lengua. Las personas caen en dos grandes grupos conocidos como “catadores” y “no catadores” en base a la densidad de papilas en su lengua, lo que también indica el número de papilas gustativas. Los no catadores pueden probar los alimentos, pero no son tan sensibles a ciertos gustos, como el amargor. La doctora Reed descubrió que no es catadora, lo que explica por qué percibió la amargura de manera diferente a otras personas que conocía. ¿Eres muy sensible a los gustos? ¿Puedes ver alguna similitud entre los miembros de tu familia?
Respuesta: Las respuestas variarán, pero una respuesta típica podría ser: Puedo comer casi cualquier cosa (¡excepto hongos!) , así que no creo que sea tan sensible a los gustos. A toda mi familia le gusta comer una variedad de alimentos, así que parece que todos tenemos el mismo nivel de sensibilidad.
Figura\(\PageIndex{8}\) La membrana basilar es la membrana delgada que se extiende desde el núcleo central de la cóclea hasta el borde. ¿Qué se ancla a esta membrana para que puedan ser activadas por el movimiento de los fluidos dentro de la cóclea?
Respuesta: Figura\(\PageIndex{8}\) Las células ciliadas se encuentran en el órgano de Corti, el cual se localiza sobre la membrana basilar. La estereocilia de esas células normalmente estaría unida a la membrana tectorial (aunque se desprenden en la micrografía debido al procesamiento del tejido).
Mira este video para conocer más sobre cómo las estructuras del oído convierten las ondas sonoras en una señal neural al mover los “pelos”, o estereocilia, del conducto coclear. Las ubicaciones específicas a lo largo de la longitud del conducto codifican frecuencias o pasos específicos. El cerebro interpreta el significado de los sonidos que escuchamos como música, habla, ruido, etc. ¿Qué estructuras auditivas son responsables de la amplificación y transferencia del sonido del oído externo al oído interno?
Respuesta: Los huesos pequeños del oído medio, los huesecillos, amplifican y transfieren el sonido entre la membrana timpánica del oído externo y la ventana ovalada del oído interno.
Mira esta animación para conocer más sobre el oído interno y ver cómo se desenrolla la cóclea, con la base en la parte posterior de la imagen y el ápice en la parte delantera. Las longitudes de onda específicas del sonido hacen vibrar regiones específicas de la membrana basilar, al igual que las teclas de un piano producen sonido a diferentes frecuencias. Con base en la animación, ¿dónde las frecuencias, desde tonos altos hasta bajos, causan actividad en las células ciliadas dentro del conducto coclear?
Respuesta: Las frecuencias altas activan las células ciliadas hacia la base de la cóclea, y las bajas frecuencias activan las células ciliadas hacia el ápice de la cóclea.
Mire este video para obtener más información sobre una sección transversal a través del cerebro que representa la vía visual desde el ojo hasta la corteza occipital. La primera mitad de la vía es la proyección desde las RGC a través del nervio óptico hasta el núcleo geniculado lateral en el tálamo a ambos lados. Esta primera fibra en la vía sinapsis sobre una célula talámica que luego se proyecta hacia la corteza visual en el lóbulo occipital donde tiene lugar la “visión” o percepción visual. Este video da una visión general abreviada del sistema visual concentrándose en el camino desde los ojos hasta el lóbulo occipital. El video hace la afirmación (a 0:45) de que “las células especializadas en la retina llamadas células ganglionares convierten los rayos de luz en señales eléctricas”. ¿Qué aspecto del procesamiento retiniano se simplifica con esa afirmación? Explica tu respuesta.
Respuesta: Los fotorreceptores convierten la energía luminosa, o fotones, en una señal electroquímica. La retina contiene células bipolares y las RGC que finalmente la convierten en potenciales de acción que se envían desde la retina al SNC. Es importante reconocer cuándo los medios populares y las fuentes en línea simplifican demasiado procesos fisiológicos complejos para que no se generen malentendidos. Este video fue creado por un fabricante de dispositivos médicos que podría estar tratando de resaltar otros aspectos del sistema visual además del procesamiento de retina. La afirmación que hacen no es incorrecta, simplemente agrupa varios pasos, lo que hace que suene como que los RGC son los transductores, más que los fotorreceptores.
Preguntas de revisión
P. ¿Qué tipo de célula receptora es responsable de transducir los estímulos del dolor?
A. mecanorreceptor
B. nociceptor
C. osmorreceptor
D. fotorreceptor
Respuesta: B
P. ¿Cuál de estos nervios craneales forma parte del sistema gustativo?
A. olfativo
B. troclear
C. trigémino
D. facial
Respuesta: D
P. ¿Qué submodalidad de sabor es sensible al pH de la saliva?
A. umami
B. agrio
C. amargo
D. dulce
Respuesta: B
P. ¿Los axones a partir de los cuales neurona en la retina conforman el nervio óptico?
A. células amacrinas
B. fotorreceptores
C. células bipolares
D. células ganglionares retinianas
Respuesta: D
P. ¿Qué tipo de célula receptora está involucrada en las sensaciones de sonido y equilibrio?
A. fotorreceptor
B. quimiorreceptor
C. mecanorreceptor
D. nociceptor
Respuesta: C
Preguntas de Pensamiento Crítico
P. El edulcorante conocido como stevia puede reemplazar la glucosa en los alimentos. ¿Qué significa la similitud molecular de la stevia con la glucosa para el sentido gustativo?
A. La molécula de stevia es similar a la glucosa de tal manera que se unirá al receptor de glucosa en papilas gustativas sensibles al dulce. Sin embargo, no es un sustrato para el metabolismo generador de ATP dentro de las células.
P. ¿Por qué el punto ciego del disco óptico en cualquiera de los dos ojos no resulta en un punto ciego en el campo visual?
A. El campo visual de cada ojo se proyecta sobre la retina a medida que la luz es enfocada por el cristalino. La información visual del campo visual derecho cae en el lado izquierdo de la retina y viceversa. El disco óptico en el ojo derecho está en el lado medial de la fóvea, que sería el lado izquierdo de la retina. No obstante, el disco óptico en el ojo izquierdo estaría en el lado derecho de esa fóvea, por lo que el campo visual derecho cae en el lado de la retina en el campo izquierdo donde no hay punto ciego.
Glosario
- alcaloide
- sustancia, generalmente de una fuente vegetal, que es químicamente básica con respecto al pH y estimulará los receptores amargos
- célula amacrina
- tipo de célula en la retina que se conecta a las células bipolares cerca de la capa sináptica externa y proporciona la base para el procesamiento temprano de la imagen dentro de la retina
- ampulla
- en el oído, la estructura en la base de un canal semicircular que contiene las células ciliadas y la cúpula para la transducción del movimiento rotacional de la cabeza
- anosmia
- pérdida del sentido del olfato; generalmente el resultado de la alteración física del primer nervio craneal
- humor acuoso
- líquido acuoso que llena la cámara anterior que contiene la córnea, el iris, el cuerpo ciliar y el cristalino del ojo
- audición
- sentido del oído
- aurícula
- estructura externa carnosa de la oreja
- membrana basilar
- en el oído, el piso del conducto coclear sobre el que se asienta el órgano de Corti
- célula bipolar
- tipo de célula en la retina que conecta los fotorreceptores a las RGC
- capsaicina
- molécula que activa los nociceptores al interactuar con un canal iónico sensible a la temperatura y es la base de sensaciones “calientes” en los alimentos picantes
- quimiorreceptor
- célula receptora sensorial que es sensible a estímulos químicos, como el gusto, el olfato o el dolor
- coroides
- tejido altamente vascular en la pared del ojo que suministra sangre a la retina externa
- cuerpo ciliar
- estructura muscular lisa en la superficie interior del iris que controla la forma de la lente a través de las fibras zónulas
- cóclea
- porción auditiva del oído interno que contiene estructuras para transducir estímulos sonoros
- conducto coclear
- espacio dentro de la porción auditiva del oído interno que contiene el órgano de Corti y está adyacente a la scala tympani y scala vestibuli a ambos lados
- fotorreceptor de cono
- uno de los dos tipos de células receptoras retinianas que se especializa para la visión del color mediante el uso de tres fotopigmentos distribuidos a través de tres poblaciones separadas de células
- contralateral
- palabra que significa “en el lado opuesto”, como en los axones que cruzan la línea media en un tramo de fibra
- córnea
- recubrimiento fibroso de la región anterior del ojo que es transparente para que la luz pueda pasar a través de él
- cúpula
- estructura especializada dentro de la base de un canal semicircular que dobla la estereocilia de las células ciliadas cuando la cabeza gira por medio del movimiento relativo del fluido encerrado
- terminación encapsulada
- configuración de una neurona receptora sensorial con dendritas rodeadas de estructuras especializadas para ayudar en la transducción de un tipo particular de sensación, como los corpúsculos lamelados en la dermis profunda y el tejido subcutáneo
- equilibrio
- sentido del equilibrio que incluye sensaciones de posición y movimiento de la cabeza
- oído externo
- estructuras en la superficie lateral de la cabeza, incluyendo la aurícula y el canal auditivo de vuelta a la membrana timpánica
- exteroceptor
- receptor sensorial que se posiciona para interpretar estímulos del ambiente externo, como fotorreceptores en el ojo o receptores somatosensoriales en la piel
- músculo extraocular
- uno de los seis músculos que se originan en los huesos de la órbita e insertan en la superficie del ojo los cuales son los encargados de mover el ojo
- túnica fibrosa
- capa externa del ojo compuesta principalmente por tejido conectivo conocido como esclerótica y córnea
- fóvea
- centro exacto de la retina en el que se enfocan los estímulos visuales para una agudeza máxima, donde la retina es más delgada, en la que no hay nada más que fotorreceptores
- terminación nerviosa libre
- configuración de una neurona receptora sensorial con dendritas en el tejido conectivo del órgano, como en la dermis de la piel, que suelen ser sensibles a estímulos químicos, térmicos y mecánicos
- sentido general
- cualquier sistema sensorial que se distribuya por todo el cuerpo e incorporado a órganos de múltiples otros sistemas, como las paredes de los órganos digestivos o la piel
- gustation
- sentido del gusto
- células receptoras gustativas
- células sensoriales en la yema gustativa que transducen los estímulos químicos de la gustación
- células ciliadas
- Células mecanorreceptoras que se encuentran en el oído interno que transducen estímulos para los sentidos de la audición y el equilibrio
- incus
- (también, yunque) osículo del oído medio que conecta el maleo con el estribo
- oblicuo inferior
- músculo extraocular responsable de la rotación lateral del ojo
- recto inferior
- músculo extraocular responsable de mirar hacia abajo
- oído interno
- estructura dentro del hueso temporal que contiene los aparatos sensoriales de audición y equilibrio
- segmento interior
- en el ojo, la sección de un fotorreceptor que contiene el núcleo y otros orgánulos principales para las funciones celulares normales
- capa sináptica interna
- capa en la retina donde las células bipolares se conectan a RGC
- interoceptor
- receptor sensorial que se posiciona para interpretar estímulos de órganos internos, como receptores de estiramiento en la pared de los vasos sanguíneos
- ipsilateral
- significado de palabra en el mismo lado, como en axones que no cruzan la línea media en un tracto de fibra
- iris
- porción coloreada del ojo anterior que rodea la pupila
- cinestesia
- sentido del movimiento corporal basado en sensaciones en los músculos esqueléticos, tendones, articulaciones y la piel
- conducto lagrimal
- conducto en la esquina medial de la órbita que drena lágrimas en la cavidad nasal
- glándula lagrimal
- glándula lateral a la órbita que produce lágrimas para lavar a través de la superficie del ojo
- recto lateral
- músculo extraocular responsable de la abducción del ojo
- lente
- componente del ojo que enfoca la luz en la retina
- elevador palpebrae superioris
- músculo que provoca elevación del párpado superior, controlado por fibras en el nervio oculomotor
- mácula
- agrandamiento en la base de un canal semicircular en el que se produce la transducción de estímulos de equilibrio dentro de la ampolla
- malleus
- (también, martillo) osículo que está directamente unido a la membrana timpánica
- mecanorreceptor
- célula receptora que transduce estímulos mecánicos en una señal electroquímica
- recto medial
- músculo extraocular responsable de la aducción del ojo
- oído medio
- espacio dentro del hueso temporal entre el canal auditivo y el laberinto óseo donde los huesecillos amplifican las ondas sonoras desde la membrana timpánica hasta la ventana ovalada
- túnica neural
- capa del ojo que contiene tejido nervioso, es decir, la retina
- nociceptor
- célula receptora que detecta estímulos de dolor
- moléculas odorantes
- sustancias químicas volátiles que se unen a las proteínas receptoras en las neuronas olfativas para estimular el sentido del olfato
- olfacción
- sentido del olfato
- bulbo olfativo
- objetivo central del primer nervio craneal; localizado en la superficie ventral del lóbulo frontal en el cerebro
- epitelio olfativo
- región del epitelio nasal donde se localizan las neuronas olfativas
- neurona sensorial olfativa
- célula receptora del sistema olfativo, sensible a los estímulos químicos del olfato, cuyos axones componen el primer nervio craneal
- opsin
- proteína que contiene el cofactor fotosensible retiniano para la fototransducción
- disco óptico
- mancha en la retina en la que los axones RGC salen del ojo y pasan los vasos sanguíneos de la retina interna
- nervio óptico
- segundo nervio craneal, que es responsable de la sensación visual
- órgano de Corti
- estructura en la cóclea en la que las células ciliadas transducen movimientos de ondas sonoras a señales electroquímicas
- osmorreceptor
- célula receptora que detecta diferencias en las concentraciones de fluidos corporales sobre la base de la presión osmótica
- huesecillos
- tres huesos pequeños en el oído medio
- otolito
- capa de cristales de carbonato de calcio ubicada en la parte superior de la membrana otolítica
- membrana otolítica
- sustancia gelatinosa en el utrículo y sáculo del oído interno que contiene cristales de carbonato de calcio y en la que se incrusta la estereocilia de las células ciliadas
- segmento exterior
- en el ojo, la sección de un fotorreceptor que contiene moléculas de opsina que transducen estímulos de luz
- capa sináptica externa
- capa en la retina en la que los fotorreceptores se conectan a las células bipolares
- ventana ovalada
- membrana en la base de la cóclea donde se une el estribo, marcando el inicio de la scala vestibuli
- conjuntiva palpebral
- membrana unida a la superficie interna de los párpados que cubre la superficie anterior de la córnea
- papila
- para la gustación, una proyección similar a un bache en la superficie de la lengua que contiene papilas gustativas
- fotoisomerización
- cambio químico en la molécula retiniana que altera el enlace para que cambie del isómero 11-cis-retinal al isómero todo-trans-retinal
- fotón
- “paquete” individual de luz
- fotorreceptor
- célula receptora especializada para responder a estímulos lumínicos
- propiocepción
- sentido de posición y movimiento del cuerpo
- propioceptor
- célula receptora que detecta cambios en la posición y aspectos cinestésicos del cuerpo
- pupila
- agujero abierto en el centro del iris que la luz pasa a través del ojo
- célula receptora
- célula que transduce estímulos ambientales en señales neuronales
- retina
- tejido nervioso del ojo en el que tiene lugar la fototransducción
- retiniano
- cofactor en una molécula de opsina que sufre un cambio bioquímico cuando es golpeado por un fotón (pronunciado con un énfasis en la última sílaba)
- células ganglionares retinianas (RGC)
- neurona de la retina que se proyecta a lo largo del segundo nervio craneal
- rodopsina
- molécula de fotopigmento que se encuentra en los fotorreceptores de varilla
- fotorreceptor de varilla
- uno de los dos tipos de células receptoras retinianas especializadas para la visión con poca luz
- ventana redonda
- membrana que marca el final del tímpano de la scala
- sáculo
- estructura del oído interno responsable de transducir la aceleración lineal en el plano vertical
- scala tymbani
- porción de la cóclea que se extiende desde el ápice hasta la ventana redonda
- scala vestibuli
- porción de la cóclea que se extiende desde la ventana oval hasta el ápice
- esclerótica
- blanco del ojo
- canales semicirculares
- estructuras dentro del oído interno responsables de transducir la información del movimiento de rotación
- modalidad sensorial
- un sistema particular para interpretar y percibir estímulos ambientales por el sistema nervioso
- somatosensación
- sentido general asociado con modalidades agrupadas como toque
- sentido especial
- cualquier sistema sensorial asociado con una estructura orgánica específica, es decir, olfato, gusto, vista, audición y equilibrio
- ganglio espiral
- localización de cuerpos celulares neuronales que transmiten información auditiva a lo largo del octavo nervio craneal
- estribo
- (también, estribo) osículo del oído medio que está unido al oído interno
- estereocilia
- serie de extensiones de membrana apical en una célula pilosa que transducen movimientos cuando se doblan
- submodalidad
- sentido específico dentro de un sentido mayor más amplio como el dulce como parte del sentido del gusto, o el color como parte de la visión
- oblicuo superior
- músculo extraocular responsable de la rotación medial del ojo
- recto superior
- músculo extraocular responsable de mirar hacia arriba
- papilas gustativas
- estructuras dentro de una papila en la lengua que contienen células receptoras gustativas
- membrana tectorial
- componente del órgano de Corti que se extiende sobre las células ciliadas, en las que se incrusta la estereocilia
- termorreceptor
- receptor sensorial especializado para estímulos de temperatura
- topográfico
- relativa a la información posicional
- transducción
- proceso de cambiar un estímulo ambiental en las señales electroquímicas del sistema nervioso
- troclea
- estructura cartilaginosa que actúa como una polea para el músculo oblicuo superior
- membrana timpánica
- tímpano
- umami
- sabor submodalidad para la sensibilidad a la concentración de aminoácidos; también llamado el sentido salado
- utrículo
- estructura del oído interno responsable de transducir la aceleración lineal en el plano horizontal
- túnica vascular
- capa media del ojo compuesta principalmente de tejido conectivo con un rico suministro de sangre
- ganglio vestibular
- localización de cuerpos celulares neuronales que transmiten información de equilibrio a lo largo del octavo nervio craneal
- vestíbulo
- en el oído, la porción del oído interno responsable de la sensación de equilibrio
- sentido visceral
- sentido asociado con los órganos internos
- visión
- sentido especial de la vista basado en la transducción de estímulos de luz
- agudeza visual
- propiedad de la visión relacionada con la nitidez del foco, que varía en relación con la posición de la retina
- humor vítreo
- líquido viscoso que llena la cámara posterior del ojo
- fibras de zonula
- conexiones fibrosas entre el cuerpo ciliar y el cristalino