
28.7: Patrones de Herencia
- Page ID
- 122776

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Diferenciar entre genotipo y fenotipo
- Describir cómo los alelos determinan los rasgos de una persona
- Resumir los experimentos de Mendel y relacionarlos con la genética humana
- Explicar la herencia de trastornos genéticos autosómicos dominantes y recesivos y vinculados al sexo
Hemos discutido los eventos que conducen al desarrollo de un recién nacido. Pero ¿qué hace que cada recién nacido sea único? La respuesta radica, por supuesto, en el ADN en el esperma y el ovocito que se combinaron para producir esa primera célula diploide, el cigoto humano.
Del genotipo al fenotipo
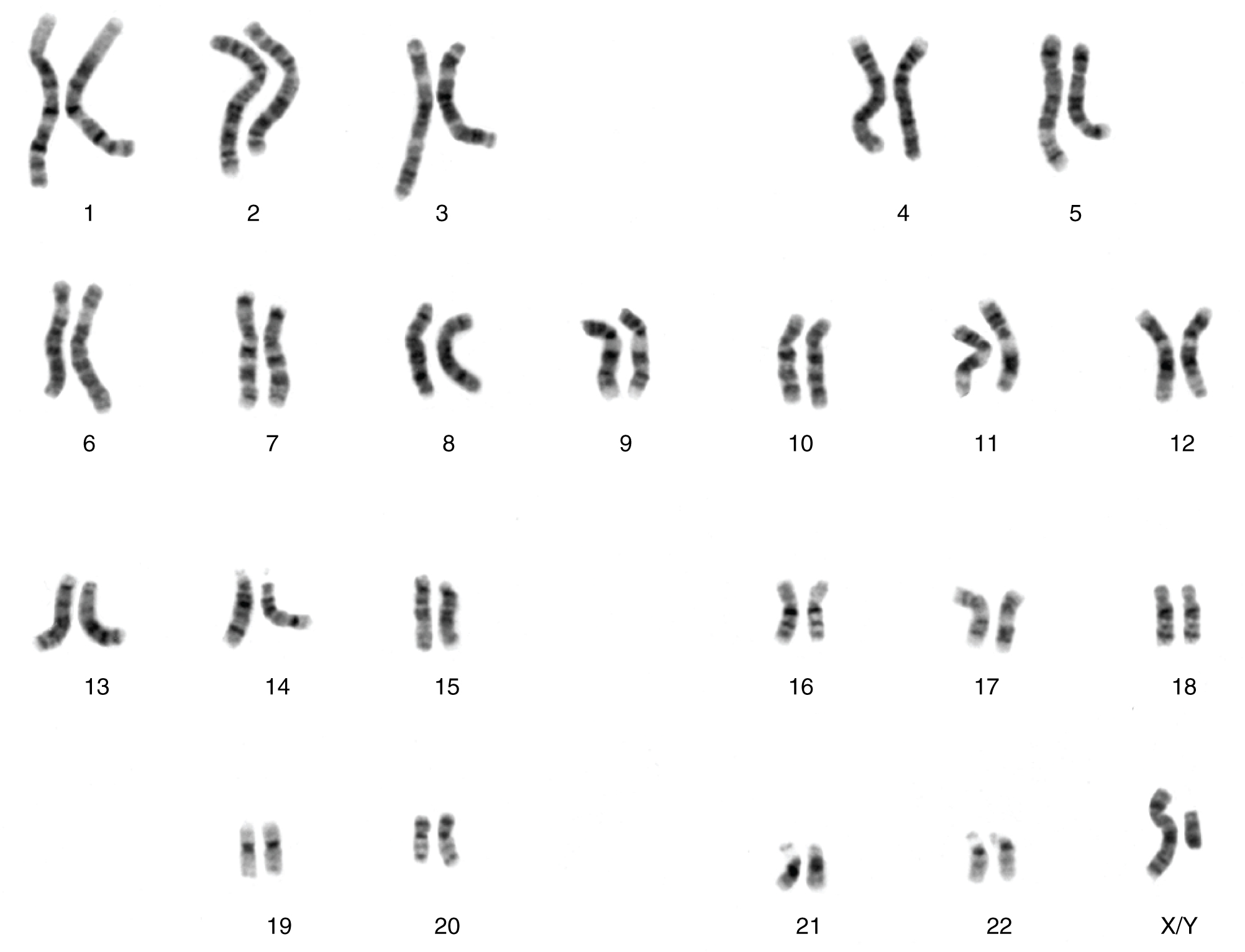
Cada célula del cuerpo humano tiene un complemento completo de ADN almacenado en 23 pares de cromosomas. La figura\(\PageIndex{1}\) muestra los pares en un arreglo sistemático llamado cariotipo. Entre estos se encuentra un par de cromosomas, llamados cromosomas sexuales, que determina el sexo del individuo (XX en hembras, XY en machos). Los 22 pares de cromosomas restantes se denominan cromosomas autosómicos. Cada uno de estos cromosomas lleva cientos o incluso miles de genes, cada uno de los cuales codifica para el ensamblaje de una proteína en particular, es decir, los genes se “expresan” como proteínas. La composición genética completa de un individuo se conoce como su genotipo. Las características que expresan los genes, ya sean físicas, conductuales o bioquímicas, son el fenotipo de una persona.
Usted hereda un cromosoma en cada par, un complemento completo de 23, de cada padre. Esto ocurre cuando el esperma y el ovocito se combinan al momento de tu concepción. Los cromosomas homólogos, los que componen un par complementario, tienen genes para las mismas características en la misma ubicación en el cromosoma. Debido a que una copia de un gen, un alelo, se hereda de cada progenitor, los alelos en estos pares complementarios pueden variar. Tomemos por ejemplo un alelo que codifica para hoyuelos. Un niño puede heredar el alelo que codifica hoyuelos en el cromosoma del padre y el alelo que codifica para piel lisa (sin hoyuelos) en el cromosoma de la madre.
Si bien una persona puede tener dos alelos idénticos para un solo gen (un estado homocigótico), también es posible que una persona tenga dos alelos diferentes (un estado heterocigótico). Los dos alelos pueden interactuar de varias maneras diferentes. La expresión de un alelo puede ser dominante, para lo cual la actividad de este gen enmascarará la expresión de un alelo no dominante, o recesivo. A veces el dominio es completo; en otras ocasiones, es incompleto. En algunos casos, ambos alelos se expresan al mismo tiempo en una forma de expresión conocida como codominancia.
En el escenario más simple, un solo par de genes determinará una única característica heredable. Sin embargo, es bastante común que múltiples genes interactúen para conferir una característica. Por ejemplo, ocho o más genes, cada uno con sus propios alelos, determinan el color de ojos en los humanos. Además, aunque cualquier persona sólo puede tener dos alelos correspondientes a un gen dado, más de dos alelos suelen existir en una población. A este fenómeno se le llama alelos múltiples. Por ejemplo, hay tres alelos diferentes que codifican el tipo sanguíneo ABO; estos se designan I A, I B e i.
Más de 100 años de estudios genéticos teóricos y experimentales, y la secuenciación y anotación más recientes del genoma humano, han ayudado a los científicos a desarrollar una mejor comprensión de cómo se expresa el genotipo de un individuo como su fenotipo. Este conjunto de conocimientos puede ayudar a científicos y profesionales médicos a predecir, o al menos estimar, algunas de las características que heredará una descendencia examinando los genotipos o fenotipos de los padres. Una aplicación importante de este conocimiento es identificar el riesgo de ciertos trastornos genéticos heredables de un individuo. Sin embargo, la mayoría de las enfermedades tienen un patrón multigénico de herencia y también pueden verse afectadas por el ambiente, por lo que examinar los genotipos o fenotipos de los padres de una persona solo proporcionará información limitada sobre el riesgo de heredar una enfermedad. Solo para un puñado de trastornos de un solo gen pueden las pruebas genéticas permitir que los médicos calculen la probabilidad con que un niño nacido de los dos padres evaluados pueda heredar una enfermedad específica.
Teoría de la Herencia de Mendel
Nuestra comprensión contemporánea de la genética se basa en la obra de un monje del siglo XIX. Trabajando a mediados del siglo XIX, mucho antes de que alguien supiera de genes o cromosomas, Gregor Mendel descubrió que los guisantes de jardín transmiten sus características físicas a las generaciones posteriores de una manera discreta y predecible. Cuando apareó, o cruzó, dos plantas de guisante de raza pura que diferían por cierta característica, la descendencia de primera generación parecía todos uno de los padres. Por ejemplo, cuando cruzó plantas de guisante altas y enanas de raza pura, todas las crías eran altas. Mendel llamó a la altura dominante porque se expresaba en descendencia cuando estaba presente en un progenitor de raza pura. Llamó al enanismo recesivo porque estaba enmascarado en la descendencia si uno de los padres de pura raza poseía la característica dominante. Tenga en cuenta que la altura y el enanismo son variaciones sobre la característica de la altura. Mendel llamó a tal variación un rasgo. Ahora sabemos que estos rasgos son la expresión de diferentes alelos del gen que codifica la altura.
Mendel realizó miles de cruces en plantas de guisante con diferentes rasgos para una variedad de características. Y en repetidas ocasiones se le ocurrieron los mismos resultados —entre los rasgos que estudió, uno siempre fue dominante y el otro siempre fue recesivo. (Recuerde, sin embargo, que esta relación dominante-recesiva entre alelos no siempre es así; algunos alelos son codominantes, y a veces la dominación es incompleta).
Utilizando su comprensión de los rasgos dominantes y recesivos, Mendel probó si un rasgo recesivo podría perderse por completo en un linaje de guisantes o si resurgiría en una generación posterior. Al cruzar las crías de segunda generación de padres de raza pura entre sí, demostró que esta última era cierta: los rasgos recesivos reaparecieron en plantas de tercera generación en una proporción de 3:1 (tres crías que tenían el rasgo dominante y una con el rasgo recesivo). Mendel propuso entonces que características como la altura fueron determinadas por “factores” heredables que fueron transmitidos, uno de cada progenitor, y heredados en parejas por descendencia.
En el lenguaje de la genética, la teoría de Mendel aplicada a los humanos dice que si un individuo recibe dos alelos dominantes, uno de cada progenitor, el fenotipo del individuo expresará el rasgo dominante. Si un individuo recibe dos alelos recesivos, entonces el rasgo recesivo se expresará en el fenotipo. Se dice que los individuos que tienen dos alelos idénticos para un gen dado, ya sean dominantes o recesivos, son homocigotos para ese gen (homo- = “same”). Por el contrario, se dice que un individuo que tiene un alelo dominante y un alelo recesivo es heterocigoto para ese gen (hetero- = “diferente” u “otro”). En este caso, se expresará el rasgo dominante, y el individuo será fenotípicamente idéntico a un individuo que posea dos alelos dominantes para el rasgo.
Es una práctica común en genética utilizar letras mayúsculas y minúsculas para representar alelos dominantes y recesivos. Usando como ejemplo las plantas de guisante de Mendel, si una planta de guisante alto es homocigota, poseerá dos alelos altos (TT). Una planta de guisante enano debe ser homocigótica porque su enanismo solo puede expresarse cuando están presentes dos alelos recesivos (tt). Una planta de guisante heterocigótica (Tt) sería alta y fenotípicamente indistinguible de una planta de guisante homocigótica alta debido al alelo alto dominante. Mendel dedujo que una relación 3:1 de dominante a recesiva sería producida por la segregación aleatoria de factores heredables (genes) al cruzar dos plantas de guisante heterocigóticas. En otras palabras, para cualquier gen dado, es igualmente probable que los padres transmitan cualquiera de sus alelos a su descendencia en un gameto haploide, y el resultado se expresará en un patrón dominante-recesivo si ambos padres son heterocigotos para el rasgo.
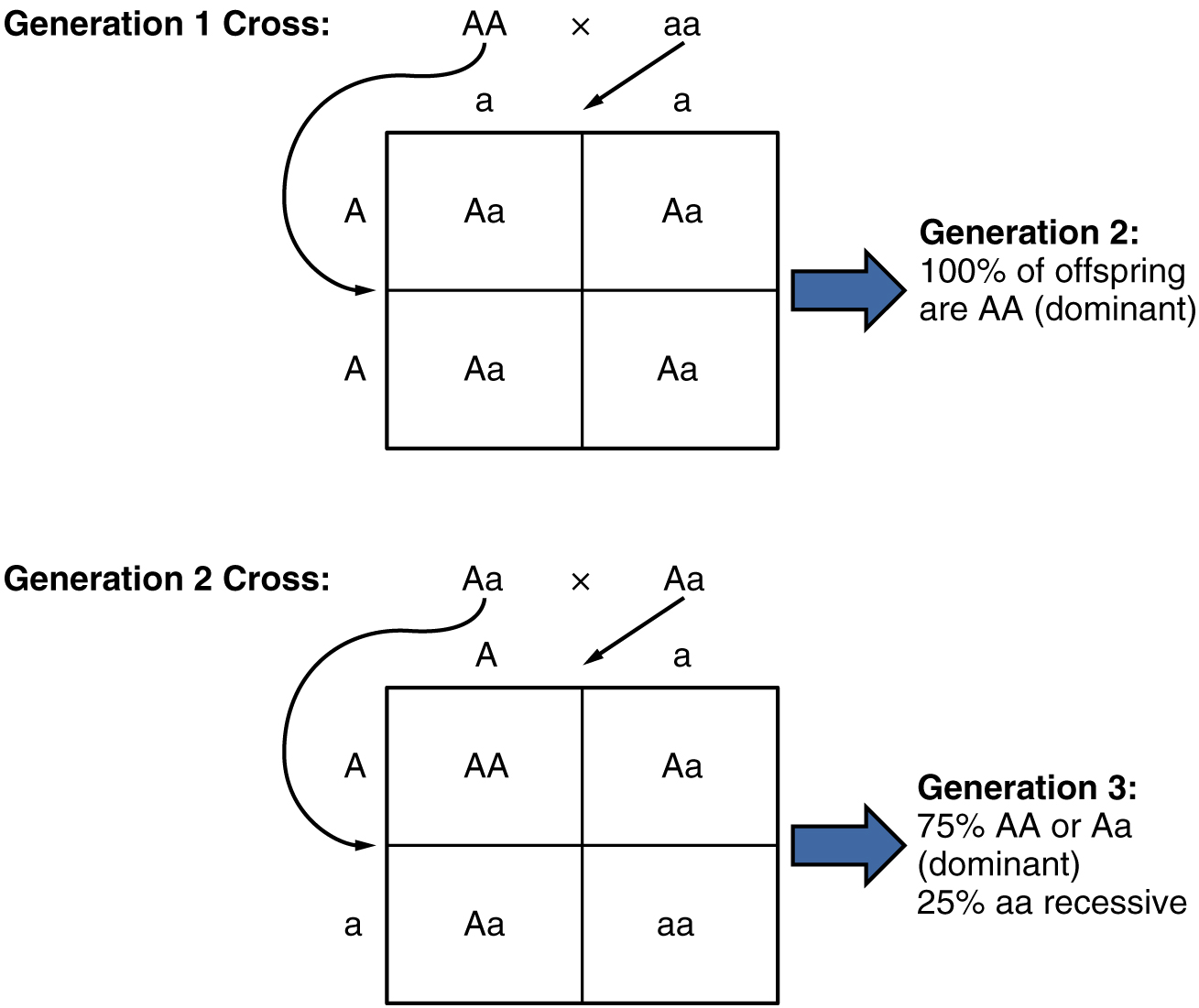
Debido a la segregación aleatoria de gametos, las leyes del azar y la probabilidad entran en juego al predecir la probabilidad de un fenotipo dado. Considera un cruce entre un individuo con dos alelos dominantes para un rasgo (AA) y un individuo con dos alelos recesivos para el mismo rasgo (aa). Todos los gametos parentales del individuo dominante serían A, y todos los gametos parentales del individuo recesivo serían a (Figura\(\PageIndex{2}\)). Todos los descendientes de esa segunda generación, heredando un alelo de cada progenitor, tendrían el genotipo Aa, y la probabilidad de expresar el fenotipo del alelo dominante sería 4 de 4, o 100 por ciento.
Esto parece bastante simple, pero el patrón de herencia se vuelve interesante cuando se cruzan los individuos Aa de segunda generación. En esta generación, el 50 por ciento de los gametos de cada padre son A y el otro 50 por ciento son a. Por el principio de segregación aleatoria de Mendel, las posibles combinaciones de gametos que la descendencia puede recibir son AA, Aa, aA (que es lo mismo que Aa) y aa. Debido a que la segregación y la fertilización son aleatorias, cada descendencia tiene un 25 por ciento de probabilidad de recibir cualquiera de estas combinaciones. Por lo tanto, si se realizara un cruce de Aa × Aa 1000 veces, aproximadamente 250 (25 por ciento) de las crías serían AA; 500 (50 por ciento) serían Aa (es decir, Aa más aA); y 250 (25 por ciento) serían aa. La relación genotípica para este patrón de herencia es 1:2:1. Sin embargo, ya hemos establecido que los individuos AA y Aa (y aA) expresan el rasgo dominante (es decir, comparten el mismo fenotipo) y, por lo tanto, pueden combinarse en un grupo. El resultado es una relación de fenotipo de tercera generación de Mendel de 3:1.
La observación de Mendel sobre las plantas de guisante también incluyó muchos cruces que involucraron múltiples rasgos, lo que le impulsó a formular el principio de surtido independiente. La ley establece que los miembros de un par de genes (alelos) de un progenitor se ordenarán independientemente de otros pares de genes durante la formación de gametos. Aplicado a las plantas de guisante, eso significa que los alelos asociados con los diferentes rasgos de la planta, como el color, la altura o el tipo de semilla, se ordenarán independientemente entre sí. Esto es cierto excepto cuando dos alelos se encuentran cerca uno del otro en el mismo cromosoma. Surtido independiente proporciona un gran grado de diversidad en la descendencia.
La genética mendeliana representa los fundamentos de la herencia, pero hay dos calificadores importantes a considerar al aplicar los hallazgos de Mendel a estudios de herencia en humanos. Primero, como ya hemos señalado, no todos los genes se heredan en un patrón dominante-recesivo. Aunque todos los individuos diploides tienen dos alelos por cada gen, los pares de alelos pueden interactuar para crear varios tipos de patrones de herencia, incluyendo dominancia incompleta y codominancia.
En segundo lugar, Mendel realizó sus estudios utilizando miles de plantas de guisante. Fue capaz de identificar una relación fenotípica 3:1 en crías de segunda generación debido a que su gran tamaño muestral superó la influencia de la variabilidad resultante del azar. En contraste, ninguna pareja humana ha tenido jamás miles de hijos. Si sabemos que un hombre y una mujer son heterocigotos por un trastorno genético recesivo, predeciríamos que uno de cada cuatro de sus hijos se vería afectado por la enfermedad. En la vida real, sin embargo, la influencia del azar podría cambiar esa proporción significativamente. Por ejemplo, si un hombre y una mujer son heterocigotos para la fibrosis quística, un trastorno genético recesivo que se expresa sólo cuando el individuo tiene dos alelos defectuosos, esperaríamos que uno de cada cuatro de sus hijos tuviera fibrosis quística. No obstante, es totalmente posible que tengan siete hijos, ninguno de los cuales se vea afectado, o que tengan dos hijos, ambos afectados. Para cada niño individual, la presencia o ausencia de un solo trastorno genético depende de qué alelos herede ese niño de sus padres.
Herencia Autosómica Dominante
En el caso de la fibrosis quística, el trastorno es recesivo al fenotipo normal. Sin embargo, una anomalía genética puede ser dominante en el fenotipo normal. Cuando el alelo dominante se localiza en uno de los 22 pares de autosomas (cromosomas no sexuales), nos referimos a su patrón de herencia como autosómico dominante. Un ejemplo de un trastorno autosómico dominante es la neurofibromatosis tipo I, una enfermedad que induce la formación de tumores dentro del sistema nervioso que conduce a deformidades cutáneas y esqueléticas. Considera una pareja en la que uno de los padres es heterocigoto para este trastorno (y que por lo tanto tiene neurofibromatosis), Nn, y uno de los padres es homocigótico para el gen normal, nn. El progenitor heterocigótico tendría una probabilidad del 50 por ciento de pasar el alelo dominante para este trastorno a su descendencia, y el progenitor homocigótico pasaría siempre al alelo normal. Por lo tanto, es igualmente probable que ocurran cuatro posibles genotipos de descendencia: Nn, Nn, nn y nn. Es decir, cada hijo de esta pareja tendría un 50 por ciento de posibilidades de heredar la neurofibromatosis. Este patrón de herencia se muestra en la Figura\(\PageIndex{3}\), en una forma llamada cuadrado de Punnett, que lleva el nombre de su creador, el genetista británico Reginald Punnett.
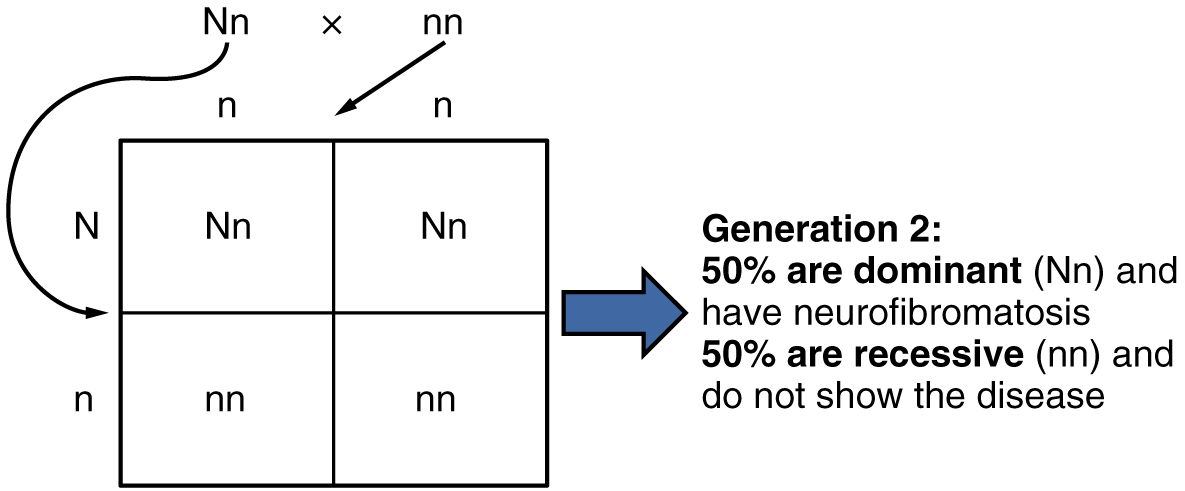
Otras enfermedades genéticas que se heredan en este patrón son el enanismo acondroplásico, el síndrome de Marfan y la enfermedad de Huntington. Debido a que los trastornos autosómicos dominantes se expresan por la presencia de un solo gen, un individuo con el trastorno sabrá que tiene al menos un gen defectuoso. La expresión de la enfermedad puede manifestarse más adelante en la vida, después de los años fértiles, lo que es el caso en la enfermedad de Huntington (discutido con más detalle más adelante en esta sección).
Herencia Autosómica Recesiva
Cuando un trastorno genético se hereda en un patrón autosómico recesivo, el trastorno corresponde al fenotipo recesivo. Los individuos heterocigotos no mostrarán síntomas de este trastorno, ya que su gen no afectado compensará. Tal individuo se llama portador. Es posible que los portadores de un trastorno autosómico recesivo nunca conozcan su genotipo a menos que tengan un hijo con el trastorno.
Un ejemplo de un trastorno autosómico recesivo es la fibrosis quística (FQ), que presentamos anteriormente. La FQ se caracteriza por la acumulación crónica de un moco espeso y tenaz en los pulmones y el tracto digestivo. Hace décadas, los niños con FQ rara vez vivían hasta la edad adulta. Con los avances en la tecnología médica, la vida media en los países desarrollados ha aumentado hasta la edad adulta media. La FQ es un trastorno relativamente común que se presenta en aproximadamente 1 en 2000 caucásicos. Un niño nacido de dos portadores de FQ tendría un 25 por ciento de probabilidades de heredar la enfermedad. Esta es la misma relación 3:1 dominante:recesiva que Mendel observó en sus plantas de guisante aplicaría aquí. El patrón se muestra en la Figura\(\PageIndex{4}\), utilizando un diagrama que rastrea la probable incidencia de un trastorno autosómico recesivo a partir de genotipos parentales.
Por otro lado, un niño nacido de un portador de CF y alguien con dos alelos no afectados tendría una probabilidad de 0 por ciento de heredar CF, pero tendría un 50 por ciento de probabilidad de ser portador. Otros ejemplos de enfermedades genéticas autosómicas recesivas incluyen el trastorno sanguíneo anemia falciforme, el trastorno neurológico fatal enfermedad de Tay-Sachs y el trastorno metabólico fenilcetonuria.

Herencia dominante o recesiva ligada al X
Un patrón de transmisión ligado al X involucra genes localizados en el cromosoma X del par 23 (Figura\(\PageIndex{5}\)). Recordemos que un macho tiene un cromosoma X y un cromosoma Y. Cuando un padre transmite un cromosoma Y, el niño es masculino, y cuando transmite un cromosoma X, el niño es femenino. Una madre sólo puede transmitir un cromosoma X, ya que ambos cromosomas sexuales son cromosomas X.
Cuando un alelo anormal para un gen que se presenta en el cromosoma X es dominante sobre el alelo normal, el patrón se describe como dominante ligado al cromosoma X. Este es el caso del raquitismo resistente a la vitamina D: un padre afectado pasaría el gen de la enfermedad a todas sus hijas, pero a ninguno de sus hijos, porque dona únicamente el cromosoma Y a sus hijos (ver Figura 28.7.5.a). Si es la madre la que se ve afectada, todos sus hijos, hombres o mujeres, tendrían un 50 por ciento de probabilidad de heredar el trastorno porque solo puede pasar un cromosoma X a sus hijos (ver Figura\(\PageIndex{5}\) .b). Para una hembra afectada, el patrón de herencia sería idéntico al de un patrón de herencia autosómico dominante en el que uno de los progenitores es heterocigoto y el otro es homocigótico para el gen normal.
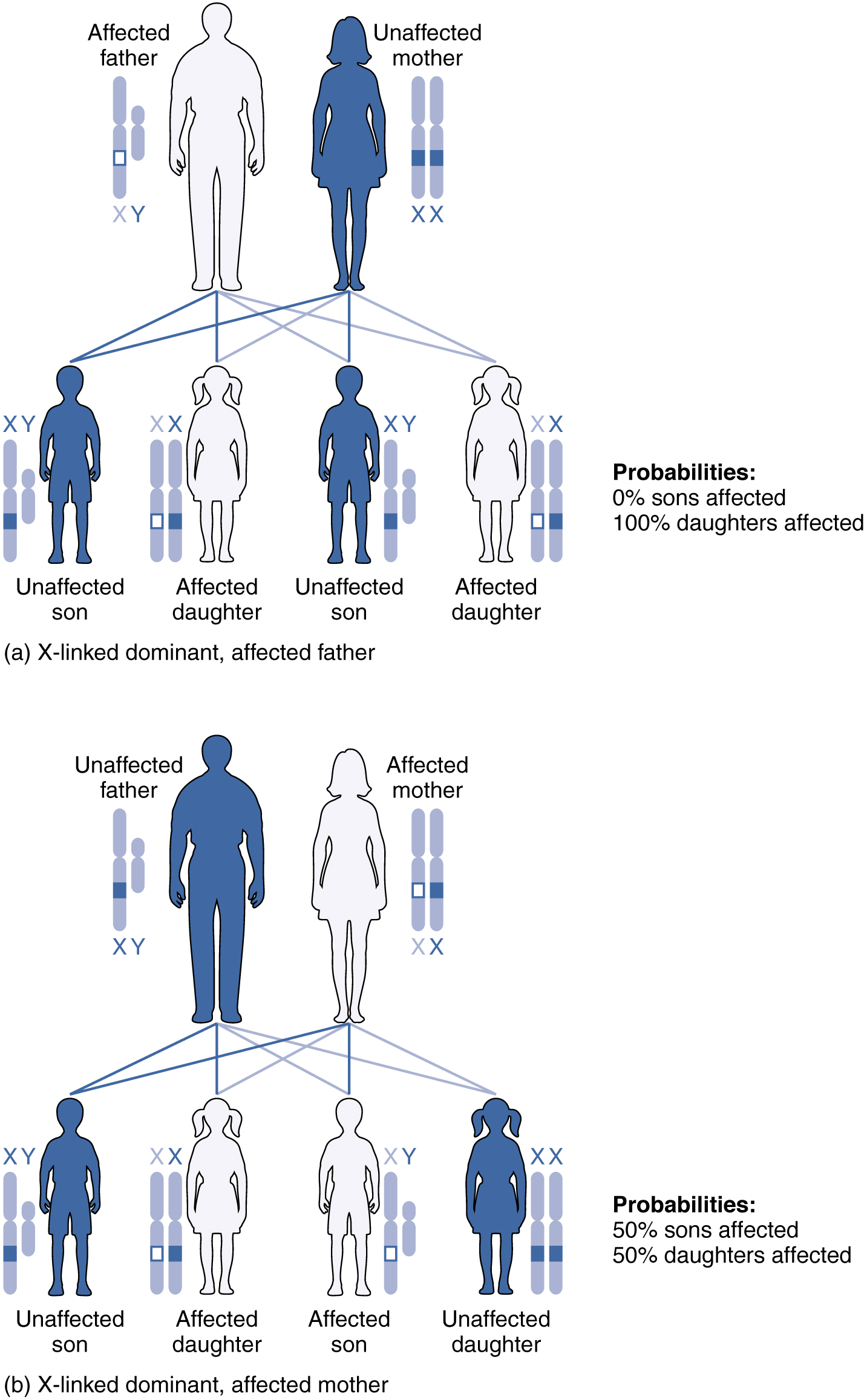
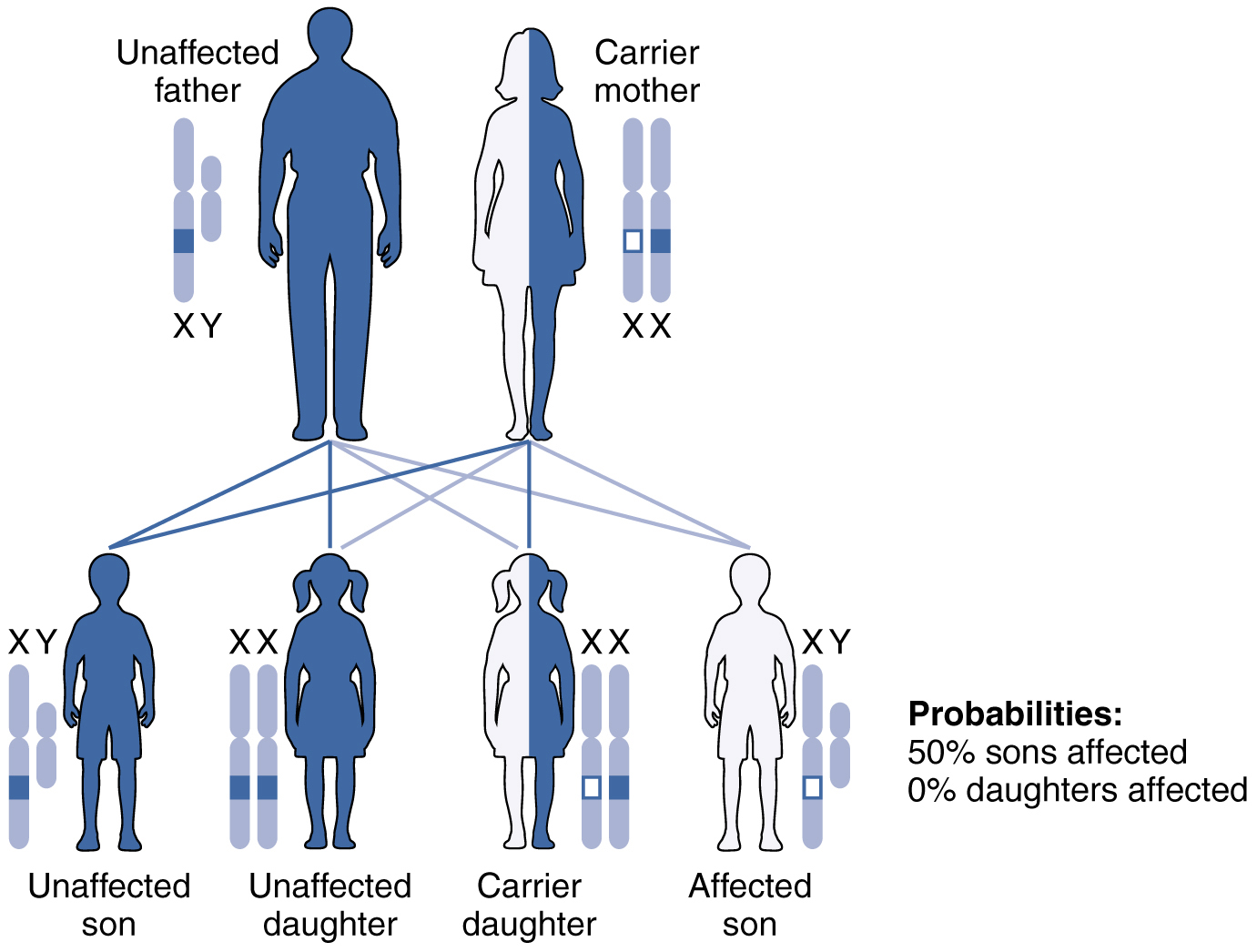
La herencia recesiva ligada al X es mucho más común porque las hembras pueden ser portadoras de la enfermedad pero aún tienen un fenotipo normal. Las enfermedades transmitidas por herencia recesiva ligada al cromosoma X incluyen daltonismo, el trastorno de coagulación sanguínea, hemofilia y algunas formas de distrofia muscular. Para un ejemplo de herencia recesiva ligada al X, considere a los padres en los que la madre es portadora no afectada y el padre es normal. Ninguna de las hijas tendría la enfermedad porque reciben un gen normal de su padre. No obstante, tienen un 50 por ciento de probabilidad de recibir el gen de la enfermedad de su madre y convertirse en portadora. En contraste, el 50 por ciento de los hijos se verían afectados (Figura\(\PageIndex{6}\)).
Con enfermedades recesivas ligadas al cromosoma X, los varones tienen la enfermedad o son genotípicamente normales, no pueden ser portadores. Las hembras, sin embargo, pueden ser genotípicamente normales, portadoras fenotípicamente normales o afectadas con la enfermedad. Una hija puede heredar el gen de una enfermedad recesiva ligada al cromosoma X cuando su madre es portadora o afectada, o su padre está afectado. La hija solo se verá afectada por la enfermedad si hereda un gen recesivo ligado al cromosoma X de ambos padres. Como puedes imaginar, los trastornos recesivos ligados al cromosoma X afectan a muchos más hombres que a mujeres. Por ejemplo, el daltonismo afecta al menos a 1 de cada 20 machos, pero solo alrededor de 1 de cada 400 hembras.
Otros patrones de herencia: dominancia incompleta, codominancia y alelos letales
No todos los trastornos genéticos se heredan en un patrón dominante-recesivo. En dominancia incompleta, la descendencia expresa un fenotipo heterocigótico que es intermedio entre el rasgo dominante homocigótico de uno de los padres y el rasgo homocigótico recesivo del otro progenitor. Un ejemplo de esto se puede ver en los dragones cuando se cruzan plantas de flores rojas y plantas de flores blancas para producir plantas de flores rosadas. En los humanos, la dominancia incompleta ocurre con uno de los genes para la textura del cabello. Cuando uno de los padres pasa un alelo de pelo rizado (el alelo incompletamente dominante) y el otro progenitor pasa un alelo de pelo liso, el efecto sobre la descendencia será intermedio, dando como resultado un cabello ondulado.
La codominancia se caracteriza por la expresión igual, distinta y simultánea de los diferentes alelos de ambos padres. Este patrón difiere de las características intermedias mezcladas que se ven en un dominio incompleto. Un ejemplo clásico de codominancia en humanos es el tipo sanguíneo ABO. Las personas son de tipo sanguíneo A si tienen un alelo para una enzima que facilita la producción de antígeno de superficie A en sus eritrocitos. Este alelo se designa I A. De la misma manera, las personas son de tipo sanguíneo B si expresan una enzima para la producción de antígeno de superficie B. Las personas que tienen alelos para ambas enzimas (I A y I B) producen ambos antígenos de superficie A y B. Como resultado, son de tipo sanguíneo AB. Debido a que se observa el efecto de ambos alelos (o enzimas), decimos que los alelos I A y I B son codominantes. También hay un tercer alelo que determina el tipo de sangre. Este alelo (i) produce una enzima no funcional. Las personas que tienen dos alelos i no producen antígenos de superficie A ni B: tienen sangre tipo O. Si una persona tiene los alelos I A e i, la persona tendrá el tipo de sangre A. Observe que no hace ninguna diferencia si una persona tiene dos alelos I A o uno I A y uno i alelo. En ambos casos, la persona es de tipo sanguíneo A. Porque I A enmascara i, decimos que I A es dominante a i. Cuadro\(\PageIndex{1}\) resume la expresión del tipo de sangre.
Mesa\(\PageIndex{1}\)
| Expresión de Tipos de Sangre | ||
|---|---|---|
| Tipo de sangre | Genotipo | Patrón de herencia |
| A | I A I A o I A i | I A es dominante a i |
| B | I B I B o I B i | I B es dominante a i |
| AB | I A I B | I A es codominante a I B |
| O | ii | Dos alelos recesivos |
Ciertas combinaciones de alelos pueden ser letales, lo que significa que impiden que el individuo se desarrolle en el útero, o causar una vida más corta. En los patrones de herencia letal recesiva, un niño que nace de dos padres heterocigotos (portadores) y que heredó el alelo defectuoso de ambos no sobreviviría. Un ejemplo de ello es Tay—Sachs, un trastorno fatal del sistema nervioso. En este trastorno, los padres con una copia del alelo para el trastorno son portadores. Si ambos transmiten su alelo anormal, su descendencia desarrollará la enfermedad y morirá en la infancia, generalmente antes de los 5 años.
Los patrones dominantes de herencia letal son mucho más raros porque ni los heterocigotos ni los homocigotos sobreviven. Por supuesto, los alelos letales dominantes que surgen de forma natural a través de la mutación y provocan abortos espontáneos o mortinatos nunca se transmiten a las generaciones posteriores. Sin embargo, algunos alelos letales dominantes, como el alelo para la enfermedad de Huntington, provocan una vida más corta pero pueden no identificarse hasta después de que la persona alcance la edad reproductiva y tenga hijos. La enfermedad de Huntington causa degeneración irreversible de las células nerviosas y muerte en el 100 por ciento de los individuos afectados, pero es posible que no se exprese hasta que el individuo alcance la mediana edad. De esta manera, los alelos letales dominantes pueden mantenerse en la población humana. A las personas con antecedentes familiares de la enfermedad de Huntington se les suele ofrecer asesoramiento genético, lo que puede ayudarles a decidir si desean o no hacerse la prueba del gen defectuoso.
Mutaciones
Una mutación es un cambio en la secuencia de nucleótidos de ADN que puede afectar o no el fenotipo de una persona. Las mutaciones pueden surgir espontáneamente de errores durante la replicación del ADN, o pueden ser el resultado de agresiones ambientales como radiación, ciertos virus o exposición al humo del tabaco u otros químicos tóxicos. Debido a que los genes codifican para el ensamblaje de proteínas, una mutación en la secuencia de nucleótidos de un gen puede cambiar la secuencia de aminoácidos y, en consecuencia, la estructura y función de una proteína. Se cree que las mutaciones espontáneas que ocurren durante la meiosis explican muchos abortos espontáneos (abortos espontáneos).
Trastornos cromosómicos
En ocasiones una enfermedad genética no es causada por una mutación en un gen, sino por la presencia de un número incorrecto de cromosomas. Por ejemplo, el síndrome de Down es causado por tener tres copias del cromosoma 21. Esto se conoce como trisomía 21. La causa más común de trisomía 21 es la no disyunción cromosómica durante la meiosis. La frecuencia de eventos de no disyunción parece aumentar con la edad, por lo que la frecuencia de tener un hijo con síndrome de Down aumenta en mujeres mayores de 36 años. La edad del padre importa menos porque la no disyunción es mucho menos probable que ocurra en un espermatozoide que en un óvulo.
Mientras que el síndrome de Down es causado por tener tres copias de un cromosoma, el síndrome de Turner es causado por tener solo una copia del cromosoma X. Esto se conoce como monosomía. El niño afectado es siempre femenino. Las mujeres con síndrome de Turner son estériles porque sus órganos sexuales no maduran.
CONEXIONES DE CARRERA
Consejero Genético
Dada la intrincada orquestación de la expresión génica, la migración celular y la diferenciación celular durante el desarrollo prenatal, es sorprendente que la gran mayoría de los recién nacidos estén sanos y libres de defectos congénitos importantes. Cuando una mujer mayor de 35 años está embarazada o tiene la intención de quedar embarazada, o su pareja tiene más de 55 años, o si hay antecedentes familiares de un trastorno genético, es posible que ella y su pareja quieran hablar con un consejero genético para discutir la probabilidad de que su hijo pueda verse afectado por un trastorno genético o cromosómico. Un consejero genético puede interpretar los antecedentes familiares de una pareja y estimar los riesgos para su futura descendencia.
Para muchas enfermedades genéticas, una prueba de ADN puede determinar si una persona es portadora. Por ejemplo, el estado del portador para X frágil, un trastorno ligado al X asociado con retraso mental, o para la fibrosis quística se puede determinar con una simple extracción de sangre para obtener ADN para la prueba. Un consejero genético puede educar a una pareja sobre las implicaciones de dicha prueba y ayudarles a decidir si se someten a pruebas. Para los trastornos cromosómicos, las opciones de prueba disponibles incluyen un análisis de sangre, amniocentesis (en la que se analiza el líquido amniótico) y muestreo de vellosidades coriónicas (en el que se analiza el tejido de la placenta). Cada uno de estos tiene ventajas e inconvenientes. Un consejero genético también puede ayudar a una pareja a sobrellevar la noticia de que uno o ambos socios son portadores de una enfermedad genética, o que su hijo por nacer ha sido diagnosticado con un trastorno cromosómico u otro defecto de nacimiento.
Para convertirse en consejero genético, se necesita completar un programa de pregrado de 4 años y luego obtener una Maestría en Ciencias en Consejería Genética de una universidad acreditada. La certificación de la Junta se obtiene después de aprobar los exámenes por la Junta Americana de Consejería Genética. Los consejeros genéticos son profesionales esenciales en muchas ramas de la medicina, pero existe una demanda particular de consejeros genéticos preconceptuales y prenatales.

Visite el sitio web de la Sociedad Nacional de Consejeros Genéticos para obtener más información sobre consejeros genéticos.

Visite el sitio web de la Junta Americana de Consejeros Genéticos, Inc., para obtener más información sobre consejeros genéticos.
Revisión del Capítulo
Hay dos aspectos en la composición genética de una persona. Su genotipo se refiere a la composición genética de los cromosomas que se encuentran en todas sus células y los alelos que se transmiten de sus padres. Su fenotipo es la expresión de ese genotipo, basado en la interacción de los alelos emparejados, así como en cómo las condiciones ambientales afectan esa expresión.
Trabajando con plantas de guisante, Mendel descubrió que los factores que dan cuenta de diferentes rasgos en los padres se transmiten discretamente a las crías en parejas, uno de cada progenitor. Articuló los principios de segregación aleatoria y surtido independiente para dar cuenta de los patrones de herencia que observó. Los factores de Mendel son genes, con diferentes variantes que se denominan alelos y esos alelos son dominantes o recesivos en expresión. Cada progenitor pasa un alelo por cada gen a la descendencia, y es igualmente probable que la descendencia herede cualquier combinación de pares de alelos. Cuando Mendel cruzó individuos heterocigotos, encontró repetidamente una relación dominante-recesiva 3:1. Postuló correctamente que la expresión del rasgo recesivo estaba enmascarada en heterocigotos pero resurgiría en su descendencia de manera predecible.
La genética humana se enfoca en identificar diferentes alelos y comprender cómo se expresan. Los investigadores médicos están especialmente interesados en la identificación de patrones de herencia para trastornos genéticos, lo que proporciona los medios para estimar el riesgo de que la descendencia de una pareja determinada herede una enfermedad o trastorno genético. Los patrones de herencia en humanos incluyen dominancia autosómica y recesividad, dominancia ligada al X y recesividad, dominancia incompleta, codominancia y letalidad. Un cambio en la secuencia de nucleótidos del ADN, que puede manifestarse o no en un fenotipo, se denomina mutación.
Preguntas de revisión
P. El síndrome de Marfan se hereda en un patrón autosómico dominante. ¿Cuál de los siguientes es cierto?
A. Las crías femeninas tienen más probabilidades de ser portadoras de la enfermedad.
B. Las crías masculinas tienen más probabilidades de heredar la enfermedad.
C. Las crías masculinas y femeninas tienen la misma probabilidad de heredar la enfermedad.
D. Las crías femeninas tienen más probabilidades de heredar la enfermedad.
Respuesta: C
P. Además de la codominancia, los antígenos del grupo sanguíneo ABO también son un ejemplo de ________.
A. dominio incompleto
B. Herencia recesiva ligada al X
C. alelos múltiples
D. Herencia letal recesiva
Respuesta: C
P. Zoe tiene fibrosis quística. ¿Cuál de las siguientes es la explicación más probable?
A. Zoe probablemente heredó un alelo defectuoso de su padre, que es portador, y un alelo normal de su madre.
B. Zoe probablemente heredó un alelo defectuoso de su madre, quien también debe tener fibrosis quística, y un alelo normal de su padre.
C. Zoe debe haber heredado alelos defectuosos de ambos padres, ambos de los cuales también deben tener fibrosis quística.
D. Zoe debe haber heredado alelos defectuosos de ambos padres, ambos de los cuales son portadores.
Respuesta: D
Preguntas de Pensamiento Crítico
P. ¿Explicar por qué era esencial que Mendel realizara sus cruces utilizando una muestra de gran tamaño?
A. Mediante el uso de tamaños de muestra grandes, Mendel minimizó el efecto de la variabilidad aleatoria resultante del azar. Esto le permitió identificar verdaderas proporciones correspondientes a herencia dominante-recesiva.
P. ¿Cómo puede una mujer portadora de un trastorno recesivo ligado al cromosoma X tener una hija afectada?
A. La única forma en que una hija afectada podría nacer es si la portadora se apareó con un varón que resultó afectado. En este caso, el 50 por ciento de las hijas se verían afectadas. Alternativamente, pero sumamente improbable, la hija podría verse afectada por una mutación espontánea.
Glosario
- alelo
- formas alternativas de un gen que ocupan un locus específico en un gen específico
- cromosoma autosómico
- en humanos, los 22 pares de cromosomas que no son los cromosomas sexuales (XX o XY)
- autosómico dominante
- patrón de herencia dominante que corresponde a un gen en uno de los 22 cromosomas autosómicos
- autosómico recesivo
- patrón de herencia recesiva que corresponde a un gen en uno de los 22 cromosomas autosómicos
- transportista
- individuo heterocigótico que no presenta síntomas de un trastorno genético recesivo pero puede transmitir el trastorno a su descendencia
- codominancia
- patrón de herencia que corresponde a la expresión igual, distinta y simultánea de dos alelos diferentes
- dominante
- describe un rasgo que se expresa tanto en forma homocigótica como heterocigótica
- letal dominante
- patrón de herencia en el que los individuos con una o dos copias de un alelo letal no sobreviven en el útero ni tienen una vida útil más corta
- genotipo
- composición genética completa de un individuo
- heterocigótico
- que tiene dos alelos diferentes para un gen dado
- homocigotos
- que tiene dos alelos idénticos para un gen dado
- dominio incompleto
- patrón de herencia en el que un genotipo heterocigótico expresa un fenotipo intermedio entre fenotipos dominante y recesivo
- cariotipo
- disposición sistemática de imágenes de cromosomas en pares homólogos
- mutación
- cambio en la secuencia de nucleótidos del ADN
- fenotipo
- manifestación física o bioquímica del genotipo; expresión de los alelos
- Cuadrado Punnett
- cuadrícula utilizada para mostrar todas las combinaciones posibles de alelos transmitidos por los padres a la descendencia y predecir la probabilidad matemática de que la descendencia herede un genotipo dado
- recesivo
- describe un rasgo que solo se expresa en forma homocigótica y se enmascara en forma heterocigótica
- letal recesivo
- patrón de herencia en el que los individuos con dos copias de un alelo letal no sobreviven en el útero o tienen una vida más corta
- cromosomas sexuales
- par de cromosomas involucrados en la determinación del sexo; en los machos, los cromosomas XY; en las hembras, los cromosomas XX
- rasgo
- variación de una característica expresada
- Vinculado a X
- patrón de herencia en el que se porta un alelo en el cromosoma X del par 23
- Dominante ligada al X
- patrón de herencia dominante que corresponde a un gen en el cromosoma X del par 23
- Recesivo ligado al X
- patrón de herencia recesiva que corresponde a un gen en el cromosoma X del par 23