
4.6: Procesamiento central táctil
- Page ID
- 124502
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Campos Receptivos e Inhibición Lateral
Los campos receptivos de las neuronas sensoriales se vuelven más complejos a medida que la información avanza por el camino. Vimos en la última lección que los mecanorreceptores tienen campos receptivos que, al tocarlos, activan la neurona. Los mecanorreceptores sinapsis en las neuronas de la columna dorsal, y esas neuronas tienen campos receptivos más complejos. Los núcleos de la columna dorsal tienen campos receptivos que se dividen en regiones centrales y envolventes. El centro del campo receptivo es resultado de la inervación directa de los mecanorreceptores. Si un estímulo toca la piel en el centro del campo receptivo de una neurona de columna dorsal, la neurona aumentará su velocidad de disparo. La estructura central/envolvente es como la de las células bipolares y ganglionares en el sistema de visión.
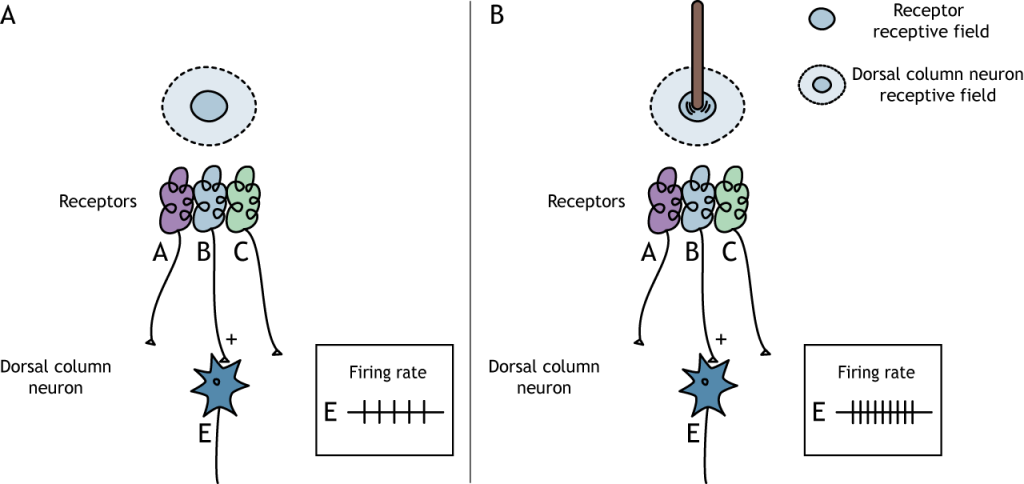
La región envolvente del campo receptivo es el resultado de la comunicación indirecta entre las neuronas receptoras y las neuronas de la columna dorsal a través de interneuronas inhibitorias. El contorno tiene un efecto inhibitorio sobre la neurona de la columna dorsal. Si un estímulo toca la piel en el entorno del campo receptivo de una neurona de columna dorsal, la neurona disminuirá su velocidad de disparo.
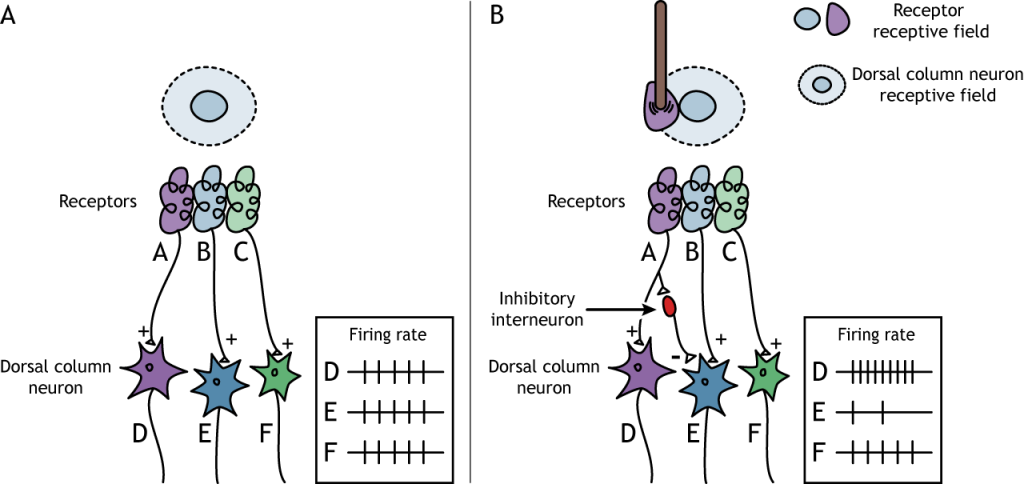
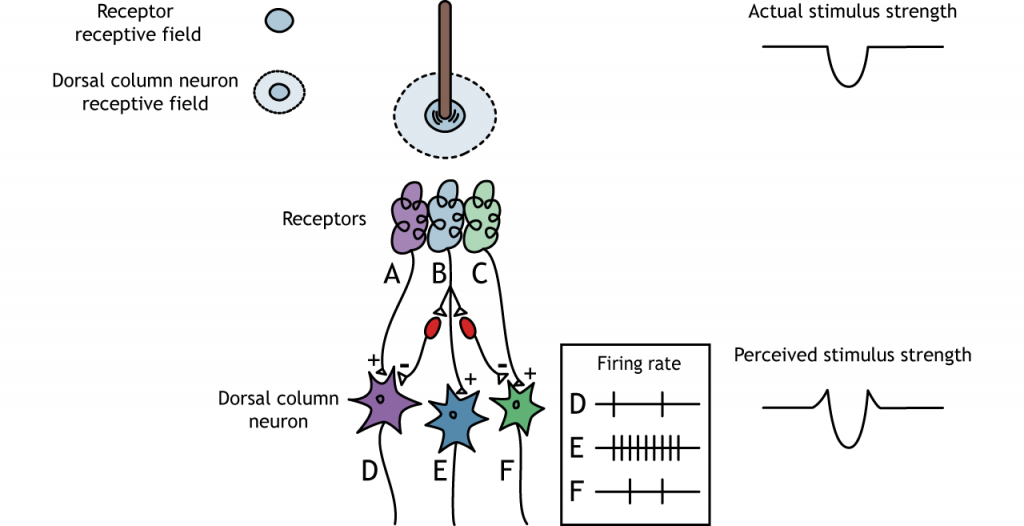
Inhibición Lateral
La estructura centro-envolvente del campo receptivo es crítica para que se produzca la inhibición lateral. La inhibición lateral es la capacidad de los sistemas sensoriales para mejorar la percepción de los bordes de los estímulos. En un punto o borde de un estímulo, debido a las interneuronas inhibidoras, la fuerza del estímulo percibida se potenciará en comparación con la fuerza del estímulo real.
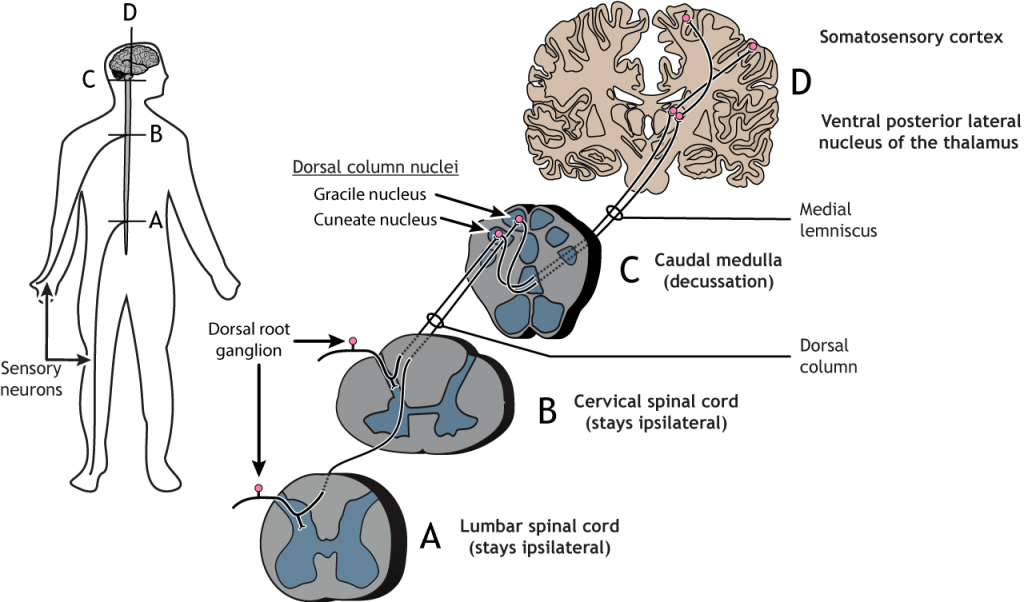
Camino al Cerebro
Vía Columna Dorsal-Lemniscus Medial
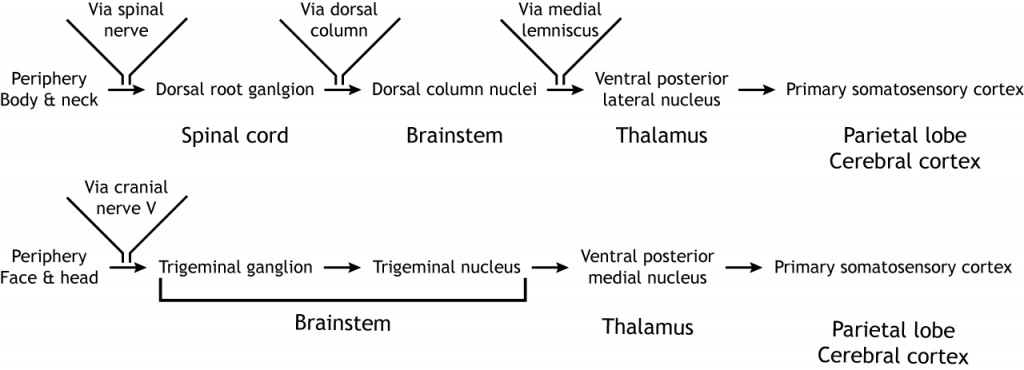
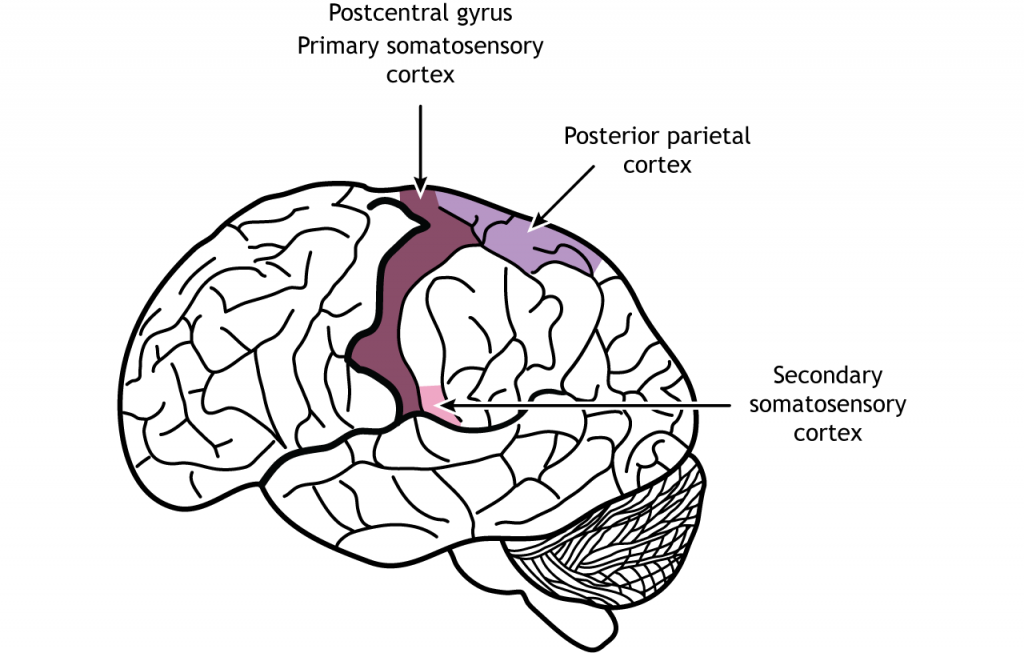
Las fibras sensoriales aferentes primarias tienen sus cuerpos celulares localizados en el ganglio de la raíz dorsal, una estructura que se encuentra justo fuera de la médula espinal. Los axones de estas neuronas de primer orden ingresan al lado dorsal ipsilateral de la médula espinal. Algunas colaterales axónicas terminan en la médula espinal y son importantes para los reflejos. La rama principal del axón asciende a la médula espinal hacia el cerebro, a través de la columna dorsal, terminando en los núcleos de la columna dorsal ubicados en el tronco encefálico. Los axones de las neuronas sensoriales en la parte inferior del cuerpo permanecen separados de los axones de las neuronas sensoriales en la parte superior del cuerpo a lo largo del camino. Estas dos poblaciones de neuronas sinapsis en diferentes regiones del tronco encefálico. Los axones de la parte inferior del cuerpo terminan en el núcleo gracile, mientras que los axones superiores del cuerpo terminan en el núcleo cuneado. Las proyecciones de las neuronas de segundo orden en los núcleos de la columna dorsal cruzan la línea media, o se decusan, y ascienden a través de un tracto de materia blanca llamado lemniscus medial. Los axones terminan en el núcleo lateral posterior ventral del tálamo. Las neuronas talámicas luego se proyectan hacia la corteza somatosensorial primaria ubicada en la circunvolución postcentral en el lóbulo parietal.
Ver el tálamo usando el BrainFacts.org 3D Brain
Ver la corteza somatosensorial primaria usando BrainFacts.org 3D Brain
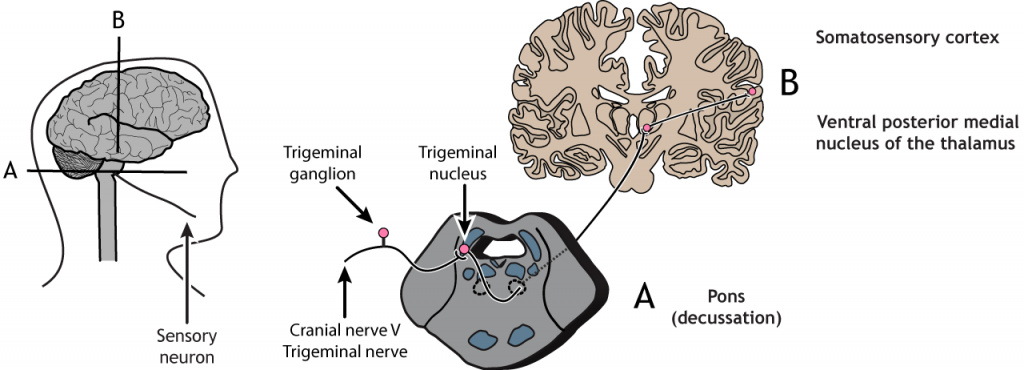
Vía del Trigémino
Los receptores sensoriales en la cara y la cabeza envían información al cerebro a través del nervio craneal V, el nervio trigémino. Las neuronas de primer orden tienen sus cuerpos celulares en el ganglio trigémino, ubicados justo fuera del tronco encefálico, y se proyectan hacia el núcleo trigémino ipsilateral en los pones. Las neuronas de segundo orden cruzan la línea media y se proyectan hasta el núcleo medial posterior ventral del tálamo. Estas neuronas luego envían proyecciones a la región de la cara de la corteza somatosensorial.
Corteza somatosensorial primaria
Anatomía
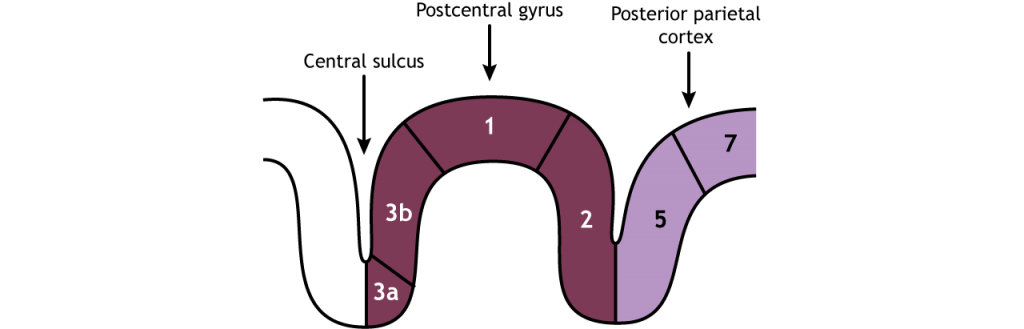
La corteza somatosensorial primaria se divide en cuatro regiones, cada una con su propia entrada y función: áreas 3a, 3b, 1 y 2. La mayoría de la información táctil de los mecanorreceptores ingresa a la región 3b, mientras que la mayoría de la información propioceptiva de los músculos ingresa a la región 3a. Estas regiones luego envían y reciben información de las áreas 1 y 2. A medida que continúa el procesamiento de la información somatosensorial, los estímulos requeridos para activar las neuronas se vuelven más complejos. Por ejemplo, el área 1 está involucrada en la detección de textura, y el área 2 está involucrada en la detección del tamaño y la forma de un objeto. La corteza parietal posterior, una región de salida importante de la corteza somatosensorial, se encuentra caudal a la circunvolución postcentral; las áreas 5 y 7 son estructuras aguas abajo que continúan procesando el tacto.
Mapa somatópico
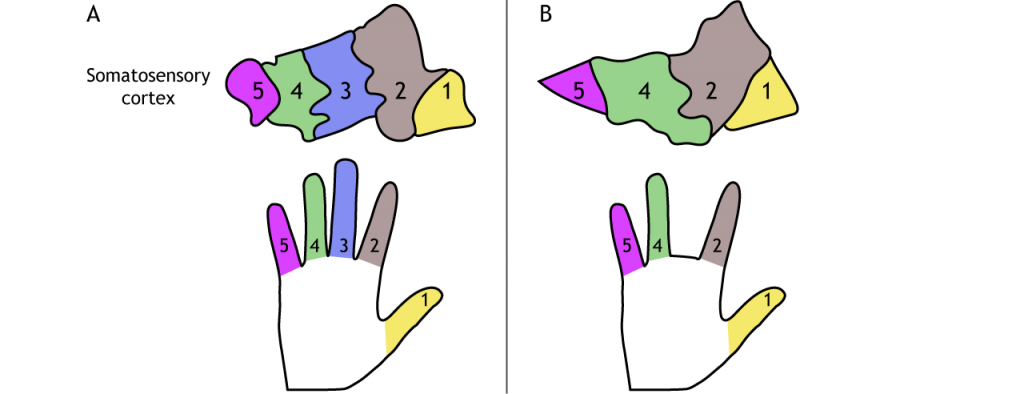
Los campos receptivos de cada neurona de orden superior aumentan de tamaño y complejidad, pero incluso las neuronas corticales están asociadas con una región específica del cuerpo. Las neuronas corticales están organizadas por la región del cuerpo que representan, por lo que las neuronas que responden a la sensación en los dedos se localizan cerca de las neuronas que responden a la sensación en la mano. Recuerda desde arriba que los axones en la columna dorsal de la parte inferior del cuerpo corren junto a, pero permanecen separados de, los axones de la parte superior del cuerpo. Esta separación, que ocurre para todas las regiones del cuerpo y en todos los niveles de la vía, crea un mapa somatotópico del cuerpo en la corteza somatosensorial primaria. Cada área de la corteza somatosensorial (Figura 23.7) tiene su propio, pero similar, mapa del cuerpo.
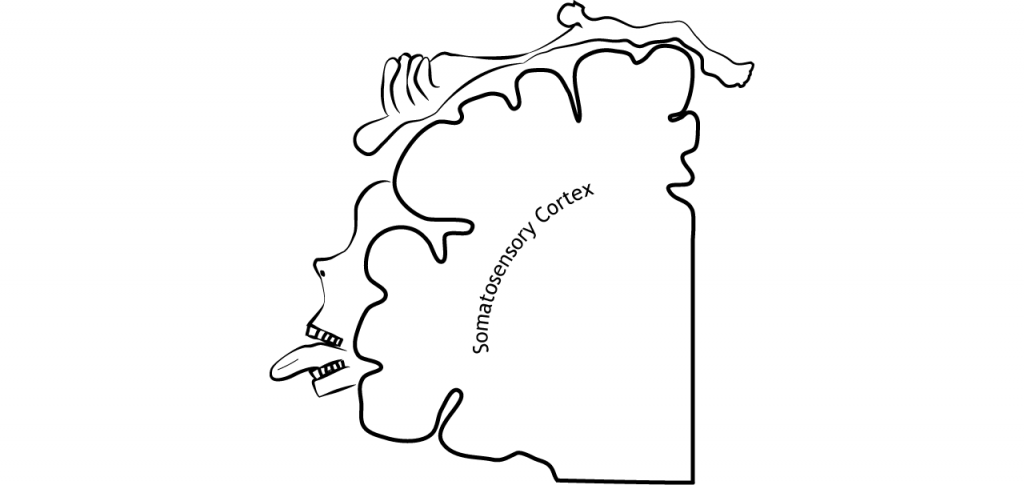
Las regiones con alta densidad de receptores en la piel y, por lo tanto, discriminación fina de dos puntos, tienen más espacio cortical dedicado a ellas. Esto significa que la representación cortical del cuerpo no es fiel a las proporciones físicas reales. Un homúnculo es una representación caricatura de cómo sería un cuerpo si el tamaño real del cuerpo fuera proporcional a la representación cortical. Las manos y los labios serían excesivamente grandes mientras que el torso, brazos y piernas, serían relativamente pequeños.
Procesamiento de nivel superior de información táctil
La corteza somatosensorial primaria envía proyecciones a otras regiones del lóbulo parietal para un procesamiento de mayor nivel de información táctil.
Corteza somatosensorial secundaria
La corteza somatosensorial secundaria (SII) se localiza en el lóbulo parietal inferior, justo por encima de la fisura lateral. Esta región, al igual que la corriente dorsal del procesamiento visual, es responsable del reconocimiento de objetos, la textura, la forma y el tamaño de discernimiento. El SII también cuenta con campos receptivos que representan regiones bilaterales del cuerpo, por lo que ambos hemisferios serán activados por el tacto a cada lado del cuerpo. El SII envía proyecciones a la corteza parietal posterior, la corteza premotora, la amígdala y el hipocampo.
Corteza parietal Posterior
La corteza parietal posterior reconoce características táctiles como orientación y movimiento. También es importante para combinar los componentes táctiles y motores de acciones como el agarre. La corteza parietal posterior sale a la corteza motora frontal.
Ver la corteza parietal posterior usando el BrainFacts.org 3D Brain
Plasticidad cortical
En la edad adulta, el cerebro es plástico, lo que significa que las conexiones sinápticas pueden reorganizarse bajo ciertas condiciones. La amputación o pérdida de un dedo, por ejemplo, conducirá a que el espacio cortical asociado sea remapeado funcionalmente por entrada de regiones vecinas de la mano. Las neuronas corticales no mueren, comienzan a ser activadas por una región diferente del cuerpo. Así mismo, la representación cortical puede expandirse con el uso o la práctica. El entrenamiento repetido de ciertos dedos puede llevar a un aumento en el espacio cortical mapeado a esos dígitos. Se cree que la plasticidad cortical subyace al fenómeno de la percepción de las extremidades fantasmas después de la amputación. En estos casos, los sujetos que han perdido una región de su cuerpo a veces todavía pueden “sentir” la parte faltante.
Claves para llevar
- Los campos receptivos se vuelven más complejos a medida que la información se mueve por el camino táctil
- La inhibición lateral mejora los bordes y las fronteras al afectar la fuerza percibida del estímulo
- Sinapsis aferentes mecanorreceptores en los núcleos de la columna dorsal en la médula. Información luego decusa y sinapsis en el núcleo ventral posterior del tálamo antes de viajar a la corteza somatosensorial primaria
- Axones sensoriales de las sinapsis de la parte inferior del cuerpo en el núcleo gracile en la columna dorsal
- Axones sensoriales de las sinapsis de la parte superior del cuerpo en el núcleo cuneado en la columna dorsal
- Información de la sinapsis cervical y corporal en el núcleo lateral posterior ventral del tálamo
- Información de la sinapsis de cabeza y cara en el núcleo medial posterior ventral del tálamo
- La corteza somatosensorial primaria está organizada en un mapa somatotópico
- La corteza es plástica y las conexiones pueden cambiar con la experiencia
¡Ponte a prueba!
Un elemento H5P interactivo ha sido excluido de esta versión del texto. Puedes verlo en línea aquí:
https://openbooks.lib.msu.edu/neuroscience/?p=512#h5p-22
Revisión adicional
- ¿Cuál es el nombre anatómico de la corteza somatosensorial primaria?
- Después de que la información somatosensorial sale del tronco encefálico, ¿debe retransmitirse a través de qué estructura antes de alcanzar la corteza somatosensorial primaria?
RESPUESTAS
Versión en video de la lección

Un elemento de YouTube ha sido excluido de esta versión del texto. Puedes verlo en línea aquí: https://openbooks.lib.msu.edu/neuroscience/?p=512