
4.2: Visión- La Retina
- Page ID
- 124503
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Anatomía de la Retina
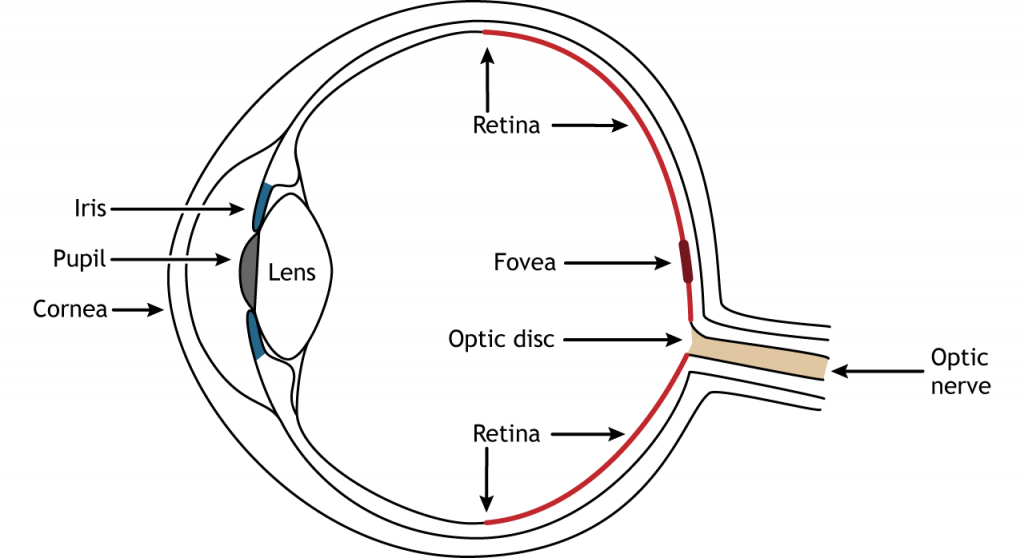
La parte frontal del ojo consiste en la córnea, la pupila, el iris y el cristalino. La córnea es la parte externa transparente del ojo. Cubre la pupila y el iris y es el primer lugar de refracción de la luz. La pupila es la abertura en el iris que permite que la luz entre en el ojo. El iris es la porción coloreada del ojo que rodea a la pupila y junto con los músculos locales pueden controlar el tamaño de la pupila para permitir que una cantidad apropiada de luz ingrese al ojo. La lente se encuentra detrás de la pupila y el iris. El cristalino refracta la luz para enfocar las imágenes en la retina. El enfoque adecuado requiere que la lente se estire o se relaje, un proceso llamado acomodación.
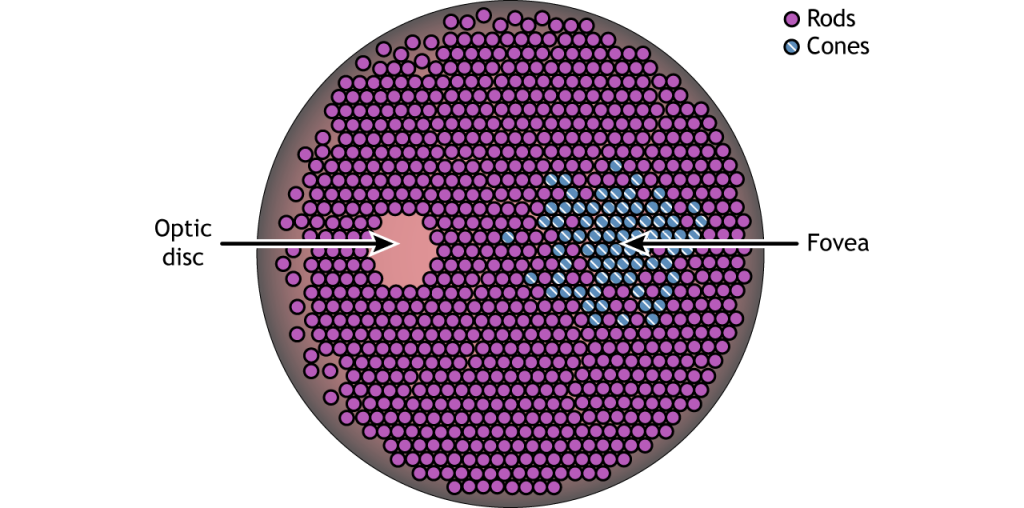
La retina es la región sensible a la luz en la parte posterior del ojo donde se encuentran los fotorreceptores, las células especializadas que responden a la luz. La retina cubre toda la parte posterior del ojo, por lo que tiene forma de cuenco. En medio del cuenco se encuentra la fóvea, la región de mayor agudeza visual, es decir, el área que puede formar las imágenes más nítidas. El nervio óptico se proyecta hacia el cerebro desde la parte posterior del ojo, transportando información de las células retinianas. Donde sale el nervio óptico, no hay fotorreceptores ya que los axones de las neuronas se están uniendo. Esta región se llama disco óptico y es la ubicación del punto ciego en nuestro campo visual.
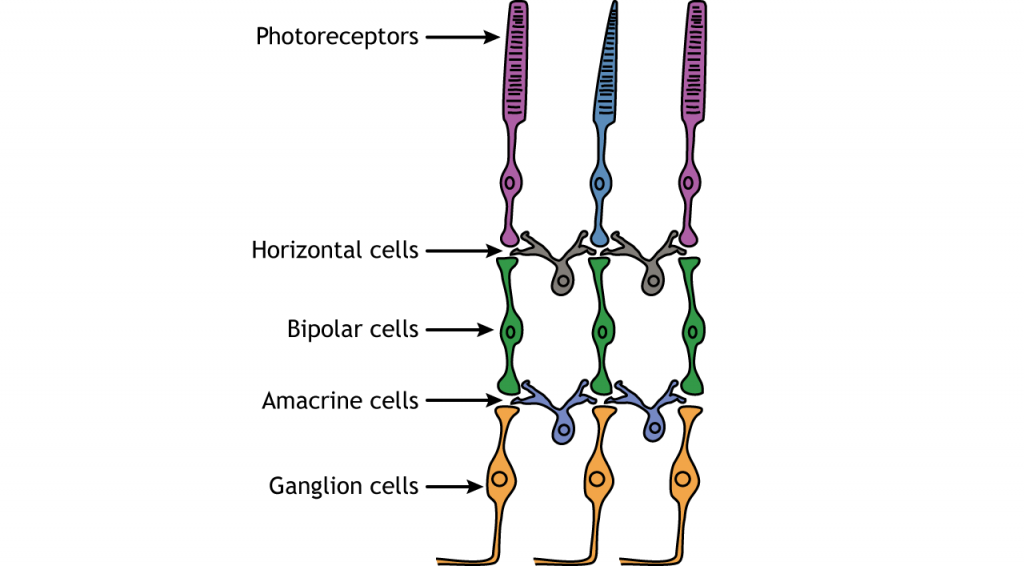
Células retinianas
Además de los fotorreceptores, hay otros cuatro tipos celulares en la retina. Los fotorreceptores sinapsis en las células bipolares y las células bipolares sinapsis en las células ganglionares. Las células horizontales y amacrinas permiten la comunicación lateral entre las neuronas.
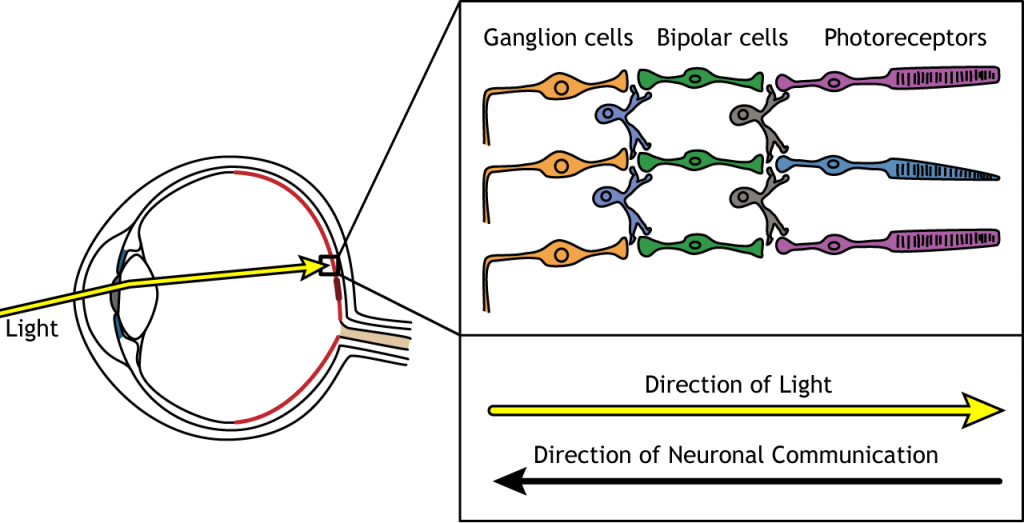
Dirección de Información
Cuando la luz entra al ojo y golpea la retina, debe pasar a través de todas las capas celulares neuronales antes de alcanzar y activar los fotorreceptores. Luego, los fotorreceptores inician la comunicación sináptica hacia las células ganglionares.
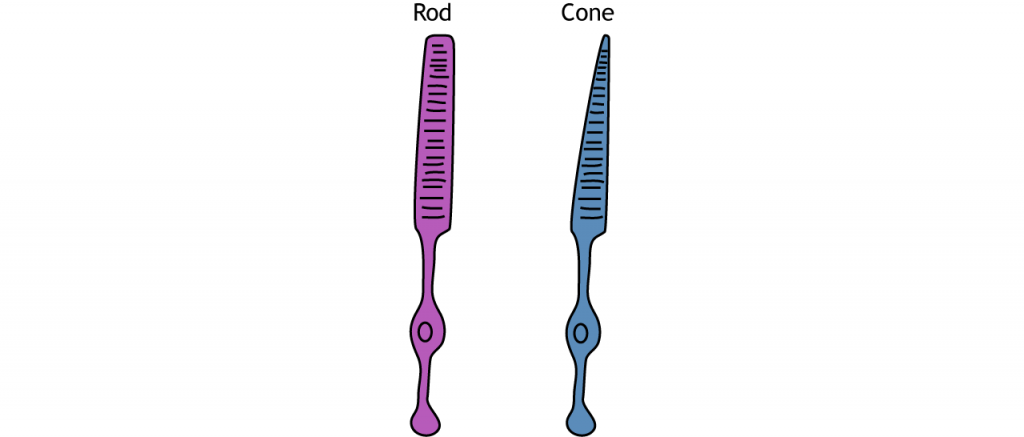
Receptores
Los fotorreceptores son los receptores especializados que responden a la luz. Existen dos tipos de fotorreceptores: bastones y conos. Las varillas son más sensibles a la luz, haciéndolas las principales responsables de la visión en condiciones de poca luz como por la noche. Los conos son menos sensibles a la luz y son más activos en condiciones de luz diurna. Los conos también son responsables de la visión del color.
Densidad del receptor
Además de tener diferentes funciones visuales, los bastones y conos también se distribuyen a través de la retina en diferentes densidades. Los conos se encuentran principalmente en la fóvea, la región de la retina con mayor agudeza visual. El resto de la retina es predominantemente bastoncillos. La región del disco óptico no tiene fotorreceptores debido a que los axones de las células ganglionares están saliendo de la retina y formando el nervio óptico.
Fototransducción
Los fotorreceptores son responsables de la transducción sensorial en el sistema visual, convirtiendo la luz en señales eléctricas en las neuronas. Para nuestros propósitos, para examinar la función de los fotorreceptores, A) nos enfocaremos en la luz en blanco y negro (no en la visión de color) y B) asumiremos que las células se están moviendo de un área de oscuridad a una zona de luz o viceversa.
Los fotorreceptores no disparan potenciales de acción; responden a cambios de luz con potenciales receptores graduados (despolarización o hiperpolarización). A pesar de esto, los fotorreceptores aún liberan glutamato sobre las células bipolares. La cantidad de glutamato liberado cambia junto con el potencial de membrana, por lo que una hiperpolarización conducirá a que se libere menos glutamato. Los fotorreceptores hiperpolarizan a la luz y despolarizan en la oscuridad. En las gráficas utilizadas en esta lección, el potencial de membrana inicial dependerá de la condición de iluminación inicial.
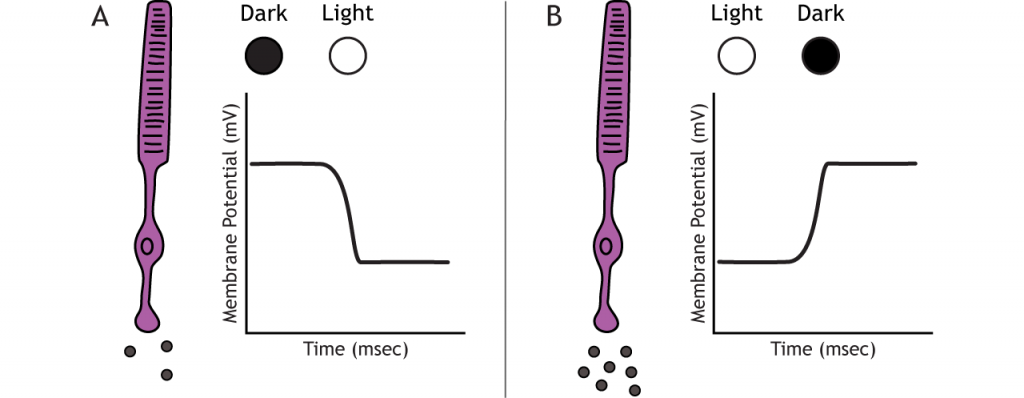
En la oscuridad, el fotorreceptor tiene un potencial de membrana que está más despolarizado que la neurona “típica” que examinamos en capítulos anteriores; el potencial de membrana fotorreceptora es de aproximadamente -40 mV. Los fotorreceptores tienen canales catiónicos abiertos que permiten la afluencia de sodio y calcio en la oscuridad. Estos canales están regulados por la presencia de GMP cíclico (GMPc), una molécula importante en cascadas de segundo mensajero que está presente en el fotorreceptor en la oscuridad.
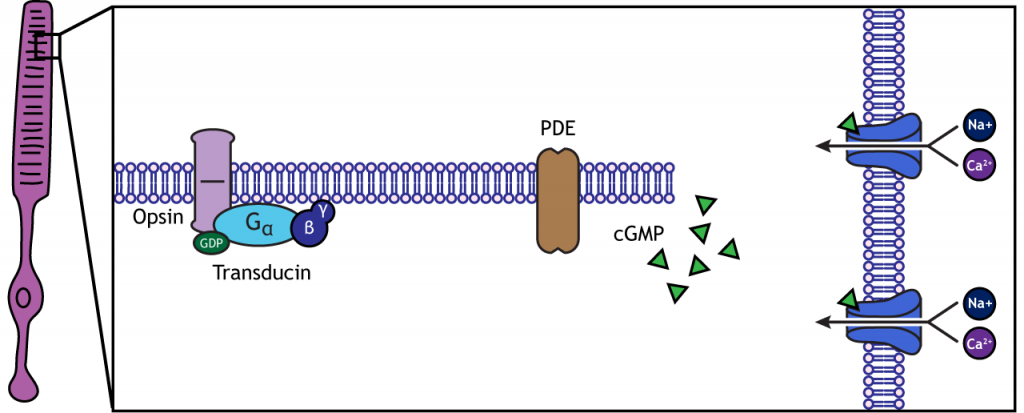
Cuando el fotorreceptor se mueve hacia la luz, la célula se hiperpolariza. La luz ingresa al ojo, llega a los fotorreceptores y provoca un cambio conformacional en una proteína especial llamada opsina. Este cambio activa una proteína G llamada transducin, que luego activa una proteína llamada fosfodiesterasa (PDE). PDE descompone cGMP a GMP, y los canales iónicos con GMPc que estaban abiertos en la oscuridad cierran. La disminución en el flujo de cationes hacia la célula hace que el fotorreceptor se hiperpolarice.
Animación 19.1. La luz que llega al fotorreceptor provoca un cambio conformacional en la proteína opsina, lo que activa la transducción de la proteína G. La transducin activa la fosfodiesterasa (PDE), que convierte el cGMP en GMP. Sin cGMP, los canales cationes se cierran, deteniendo la afluencia de iones positivos. Esto da como resultado una hiperpolarización de la célula. 'Fototransducción' por Casey Henley está bajo una Licencia Creative Commons Atribución No Comercial Compartir Igual (CC BY-NC-SA) 4.0 Internacional. Ver imagen estática de animación.
Transmisión de Información dentro de Retina
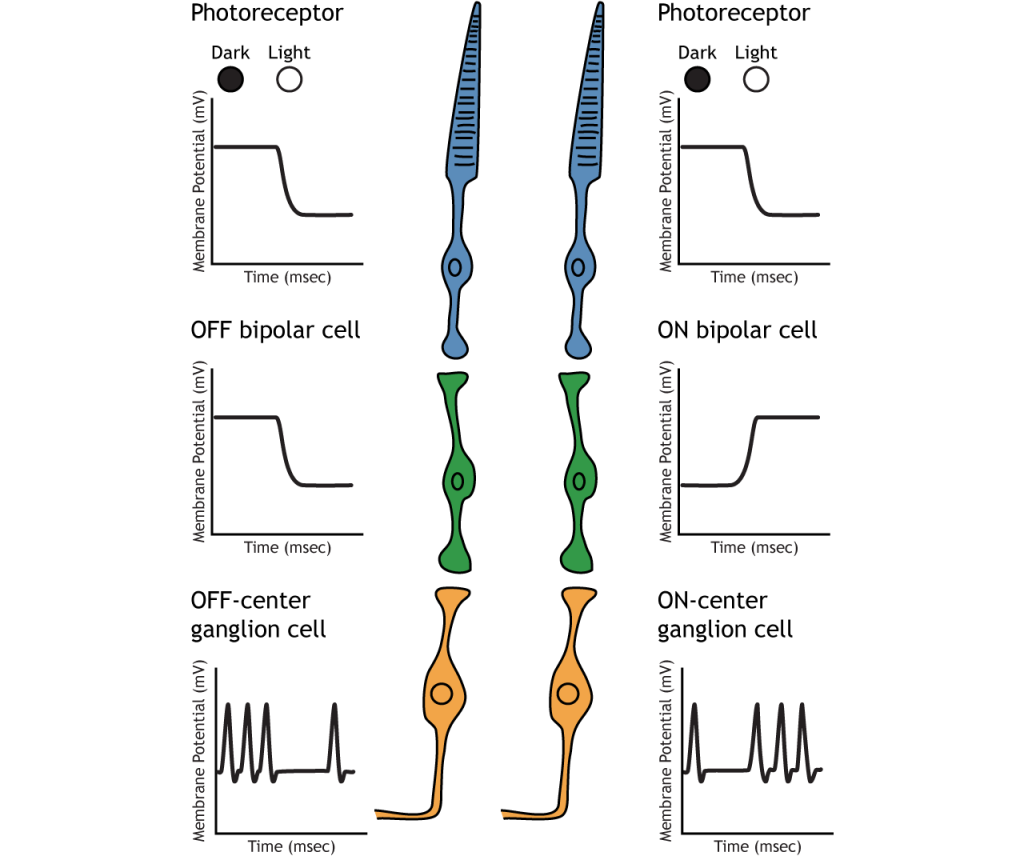
Sinapsis de fotorreceptores sobre células bipolares en la retina. Hay dos tipos de células bipolares: OFF y ON. Estas células responden de manera opuesta al glutamato liberado por los fotorreceptores porque expresan diferentes receptores de glutamato. Al igual que los fotorreceptores, las células bipolares no disparan el potencial de acción y solo responden con potenciales postsinápticos graduados.
OFF Células Bipolares
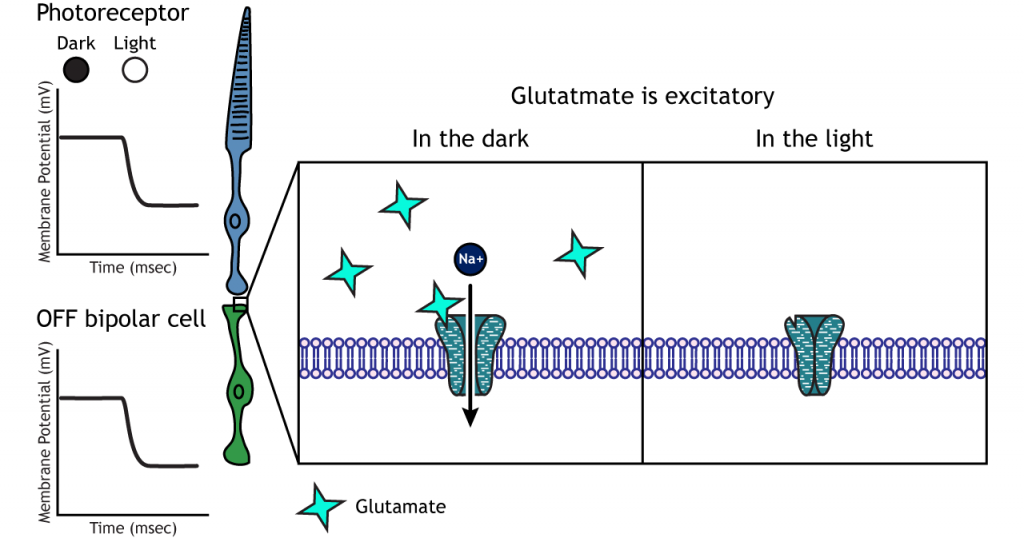
En las células bipolares OFF, el glutamato liberado por el fotorreceptor es excitatorio. Las células bipolares OFF expresan receptores ionotrópicos de glutamato. En la oscuridad, el glutamato liberado por el fotorreceptor activa los receptores ionotrópicos, y el sodio puede fluir hacia la célula, despolarizando el potencial de membrana. A la luz, la ausencia de glutamato hace que los receptores ionotrópicos se cierren, impidiendo la afluencia de sodio, hiperpolarizando el potencial de membrana.
Células bipolares
En las células bipolares ON, el glutamato liberado por el fotorreceptor es inhibitorio. Las células bipolares ON expresan receptores metabotrópicos de glutamato. En la oscuridad, el glutamato liberado por el fotorreceptor activa los receptores metabotrópicos, y las proteínas G cierran los canales catiónicos en la membrana, deteniendo la afluencia de sodio y calcio, hiperpolarizando el potencial de la membrana. A la luz, la ausencia de glutamato da como resultado que los canales iónicos estén abiertos y permitan la afluencia de cationes, despolarizando el potencial de membrana.
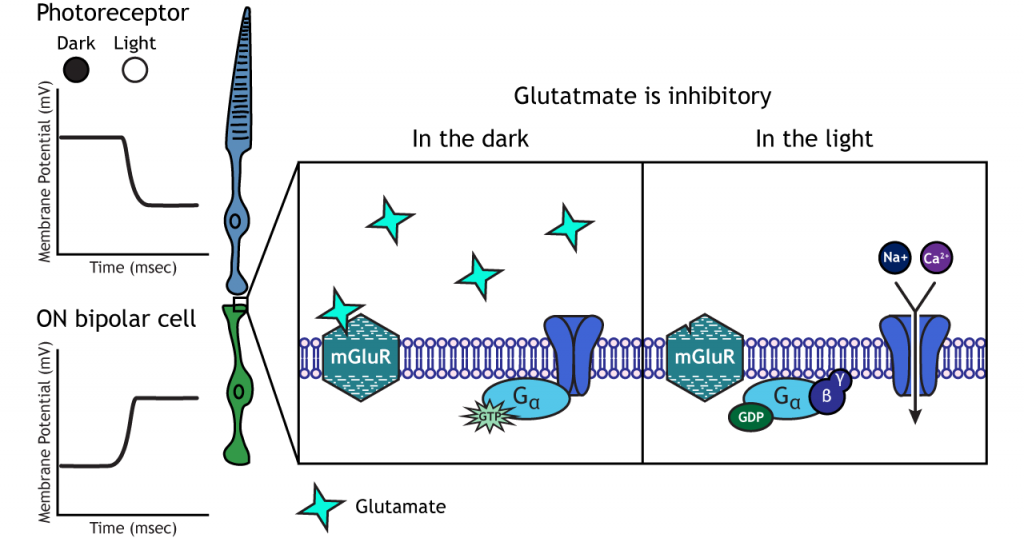
Células ganglionares
Sinapsis de células bipolares OFF y ON en células ganglionares Off-center y On-center, respectivamente. Las células ganglionares son el único tipo celular que envía información fuera de la retina, y también son la única célula que dispara potenciales de acción. Las células ganglionares disparan en todas las condiciones de iluminación, pero es la velocidad de disparo relativa la que codifica la información sobre la luz. Un movimiento de la oscuridad a la luz hará que las células ganglionares fuera del centro disminuyan su velocidad de disparo y las células ganglionares On-center aumenten su velocidad de disparo.
Campos Receptivos
Cada célula bipolar y ganglionar responde al estímulo de luz en un área específica de la retina. Esta región de la retina es el campo receptivo de la célula. Los campos receptivos en la retina son circulares.
El tamaño del campo receptivo puede variar. La fóvea tiene campos receptivos más pequeños que la retina periférica. El tamaño depende del número de fotorreceptores que sinapsan en una célula bipolar dada y del número de células bipolares que sinapsan en una célula ganglionar dada, también llamada la cantidad de convergencia.
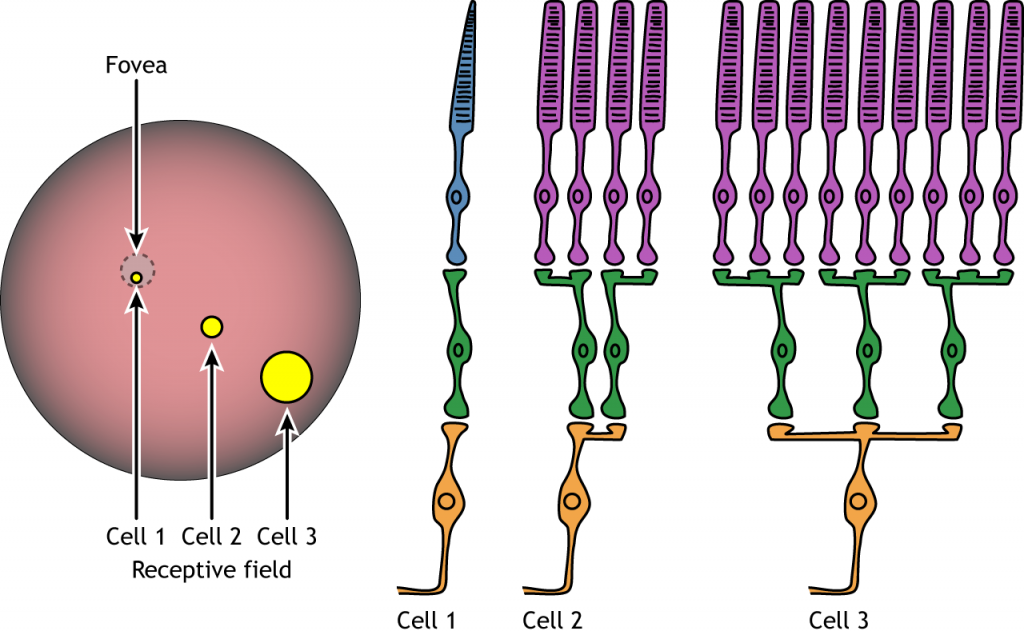
Ejemplo de Campo Receptivo
Usemos un ejemplo de una célula bipolar ON para observar la estructura de los campos receptivos en la retina. Los campos receptivos de células bipolares y ganglionares se dividen en dos regiones: el centro y el entorno. El centro del campo receptivo es el resultado de la inervación directa entre los fotorreceptores, las células bipolares y las células ganglionares. Si un punto de luz cubre el centro del campo receptivo, la célula bipolar ON se despolarizaría, como se discutió anteriormente; la luz golpea el fotorreceptor, se hiperpolariza, disminuyendo la liberación de glutamato. Menos glutamato conduce a una menor inhibición de la célula bipolar ON, y se despolariza.
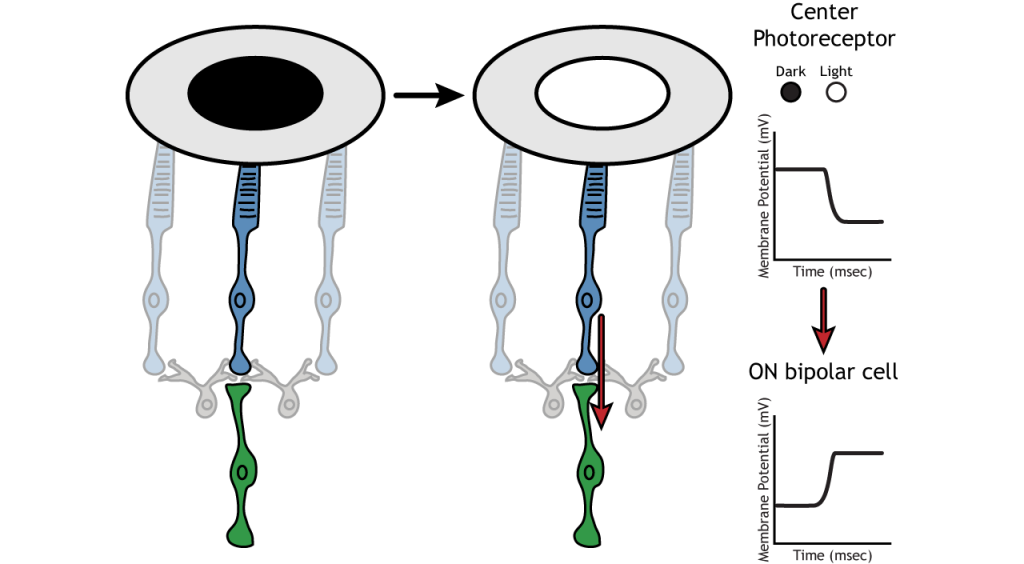
La porción envolvente del campo receptivo es el resultado de la comunicación indirecta entre las neuronas retinianas a través de células horizontales y amacrinas. La envolvente también tiene un efecto contrario sobre la célula bipolar o ganglionar en comparación con el efecto de la región central. Si un punto de luz cubre la porción envolvente, la célula bipolar ENCENDIDA respondería hiperpolarizando. La luz provocaría que el fotorreceptor en el entorno se hiperpolarizara. Esto provocaría que la célula horizontal también se hiperpolarizara. Las células horizontales tienen efectos sinápticos inhibitorios, por lo que una hiperpolarización en la célula horizontal conduciría a una despolarización en el fotorreceptor central. El fotorreceptor central provocaría entonces una hiperpolarización en la célula bipolar ON. Estos efectos imitan a los que se ven cuando el centro está en la oscuridad. Entonces, aunque el fotorreceptor central no está experimentando directamente un cambio en las condiciones de iluminación, las neuronas responden como si se estuvieran moviendo hacia la oscuridad.
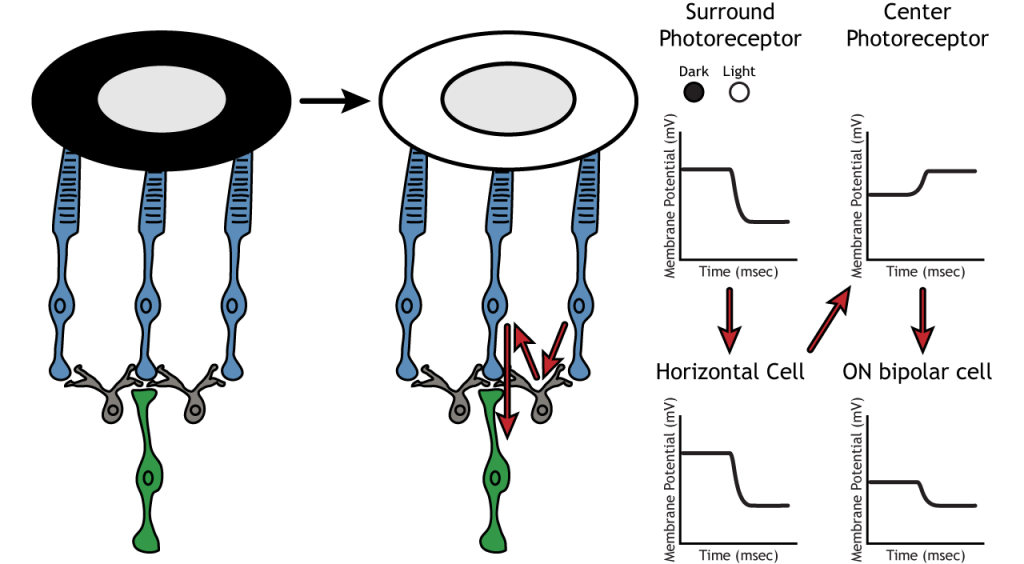
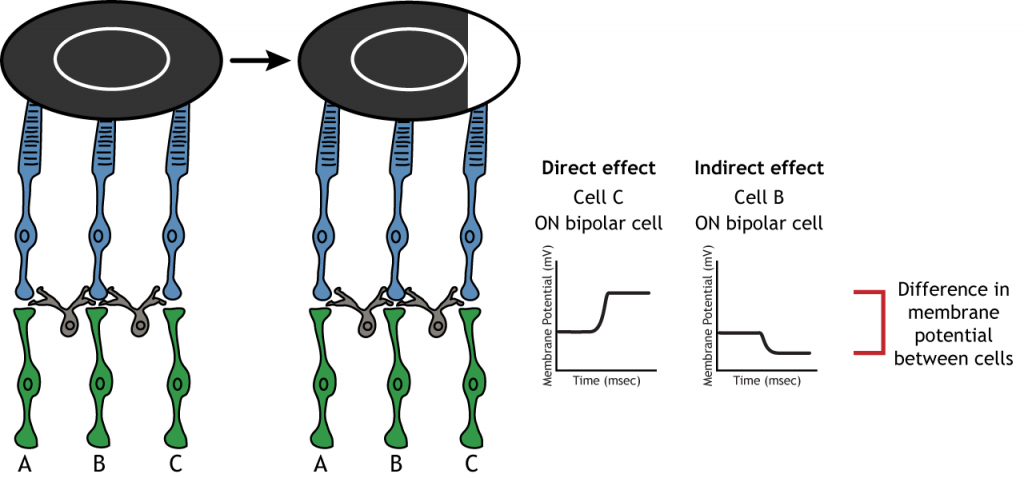
Inhibición Lateral
La estructura centro-envolvente del campo receptivo es crítica para que se produzca la inhibición lateral. La inhibición lateral es la capacidad de los sistemas sensoriales para mejorar la percepción de los bordes de los estímulos. Es importante señalar que los fotorreceptores que están en el entorno de una célula bipolar también estarían en el centro de una célula bipolar diferente. Esto conduce a un efecto sináptico directo sobre una célula bipolar a la vez que tiene un efecto indirecto sobre otra célula bipolar.
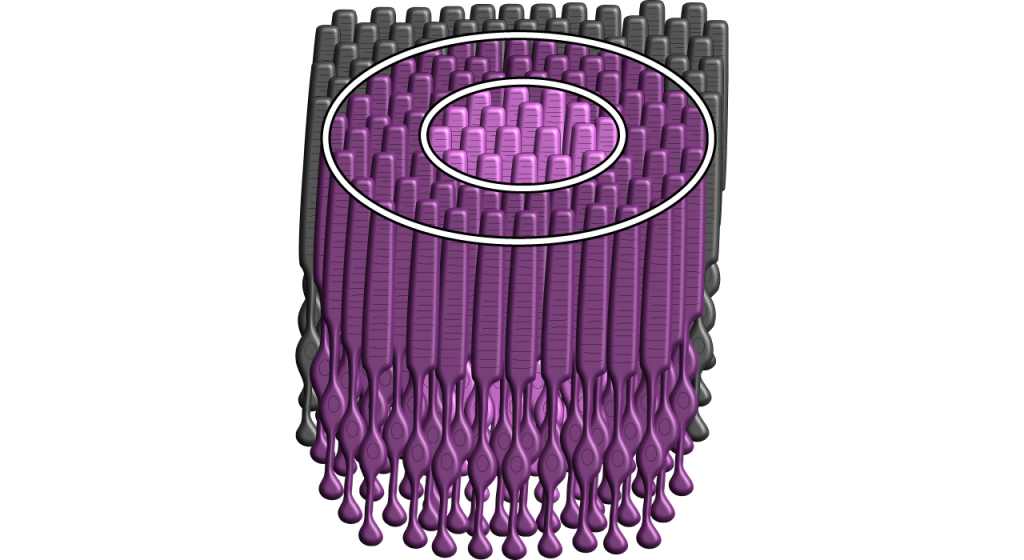
Si bien algunas de las imágenes aquí utilizadas simplificarán el campo receptivo a una célula en el centro y a una pareja en el entorno, es importante recordar que los fotorreceptores cubren toda la superficie de la retina, y el campo receptivo es bidimensional. Dependiendo del nivel de convergencia en las células bipolares y ganglionares, los campos receptivos pueden contener muchos fotorreceptores.
Claves para llevar
- Los fotorreceptores y las células bipolares no disparan potenciales de acción
- Los fotorreceptores hiperpolarizan en la luz
- Las células bipolares ON expresan receptores de glutamato metabotrópicos inhibidores
- Las células bipolares OFF expresan receptores de glutamato ionotrópicos excitadores
- Los campos receptivos son circulares, tienen un centro y un entorno, y varían en tamaño
- La estructura de campo receptivo permite que se produzca inhibición lateral
¡Ponte a prueba!
Un elemento H5P interactivo ha sido excluido de esta versión del texto. Puedes verlo en línea aquí:
https://openbooks.lib.msu.edu/neuroscience/?p=438#h5p-18
Revisión adicional
- Compara y contrasta varillas y conos.
- Compara y contrasta la fóvea y el disco óptico.
RESPUESTAS
Versión en video de la lección

Un elemento de YouTube ha sido excluido de esta versión del texto. Puedes verlo en línea aquí: https://openbooks.lib.msu.edu/neuroscience/?p=438