
7.2: Descanso, Calificada y Potencial de Acción
- Page ID
- 121039
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Diferenciar entre potencial de reposo, gradual y de acción con referencia a las estructuras de la neurona, así como los movimientos iónicos y los potenciales de membrana
El sistema nervioso de la mosca común de laboratorio, Drosophila melanogaster, contiene alrededor de 100 mil neuronas, el mismo número que una langosta. Este número se compara con 75 millones en el ratón y 300 millones en el pulpo. Un cerebro humano contiene alrededor de 86 mil millones de neuronas. A pesar de estos números muy diferentes, los sistemas nerviosos de estos animales controlan muchos de los mismos comportamientos, desde reflejos básicos hasta comportamientos más complicados como encontrar comida y cortejar compañeros. La capacidad de las neuronas para comunicarse entre sí, así como con otros tipos de células, subyace a todos estos comportamientos. La mayoría de las neuronas comparten los mismos componentes celulares. Pero las neuronas también están altamente especializadas: diferentes tipos de neuronas tienen diferentes tamaños y formas que se relacionan con sus roles funcionales.
Partes de una neurona
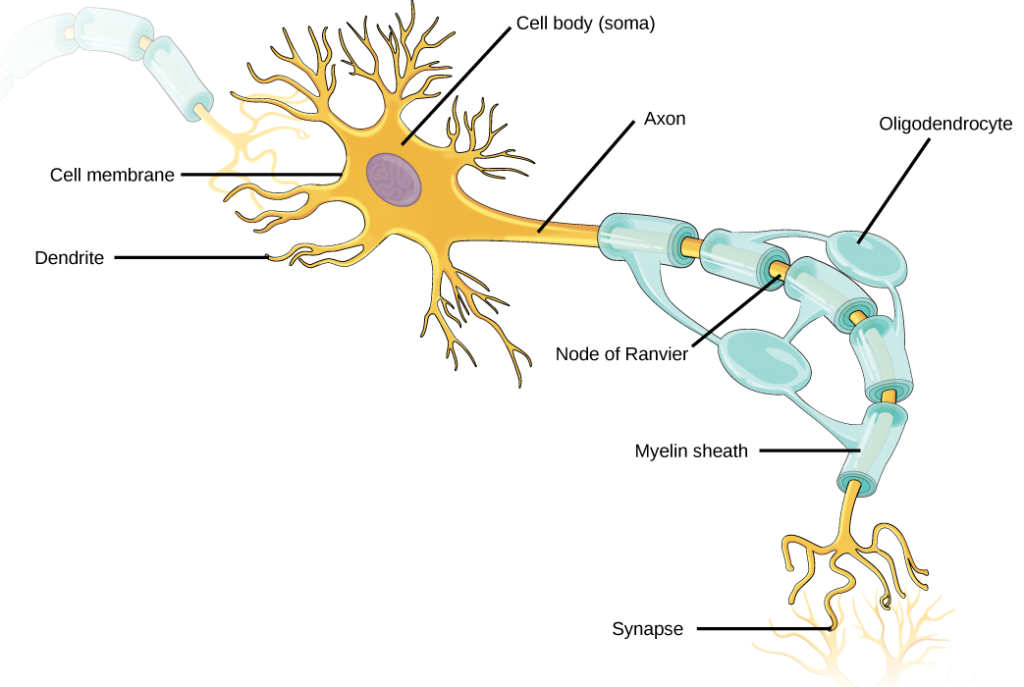
Al igual que otras células, cada neurona tiene un cuerpo celular (o soma) que contiene un núcleo, retículo endoplásmico liso y rugoso, aparato de Golgi, mitocondrias y otros componentes celulares. Las neuronas también contienen estructuras únicas, ilustradas en la Figura 7.5 para recibir y enviar las señales eléctricas que hacen posible la comunicación neuronal. Las dendritas son estructuras arbóreas que se extienden lejos del cuerpo celular para recibir mensajes de otras neuronas en uniones especializadas llamadas sinapsis. Aunque algunas neuronas no tienen ninguna dendrita, algunos tipos de neuronas tienen múltiples dendritas. Las dendritas pueden tener pequeñas protuberancias llamadas espinas dendríticas, que aumentan aún más el área de superficie para posibles conexiones sinápticas.
Una vez que una señal es recibida por la dendrita, entonces viaja pasivamente al cuerpo celular. El cuerpo celular contiene una estructura especializada, el montículo axónico que integra señales de múltiples sinapsis y sirve como unión entre el cuerpo celular y un axón. Un axón es una estructura similar a un tubo que propaga la señal integrada a terminaciones especializadas llamadas terminales axónicas. Estos terminales a su vez sinapsis en otras neuronas, músculos u órganos diana. Los químicos liberados en los terminales axones permiten que las señales se comuniquen a estas otras celdas. Las neuronas suelen tener uno o dos axones, pero algunas neuronas, como las células amacrinas en la retina, no contienen ningún axón. Algunos axones están cubiertos con mielina, que actúa como aislante para minimizar la disipación de la señal eléctrica a medida que viaja por el axón, incrementando en gran medida la velocidad en la conducción. Este aislamiento es importante ya que el axón de una neurona motora humana puede durar hasta un metro, desde la base de la columna hasta los dedos de los pies. La vaina de mielina en realidad no es parte de la neurona. La mielina es producida por las células gliales. A lo largo del axón hay huecos periódicos en la vaina de mielina. Estas brechas se denominan nodos de Ranvier y son sitios donde la señal se “recarga” a medida que viaja a lo largo del axón.
Es importante señalar que una sola neurona no actúa sola; la comunicación neuronal depende de las conexiones que las neuronas hagan entre sí (así como con otras células, como las células musculares). Las dendritas de una sola neurona pueden recibir contacto sináptico de muchas otras neuronas. Por ejemplo, se cree que las dendritas de una célula de Purkinje en el cerebelo reciben contacto de hasta 200,000 otras neuronas.
Ejercicio\(\PageIndex{1}\)
¿Cuál de las siguientes afirmaciones es falsa?
- El soma es el cuerpo celular de una célula nerviosa.
- La vaina de mielina proporciona una capa aislante a las dendritas.
- Los axones llevan la señal del soma al objetivo.
- Las dendritas llevan la señal al soma.
Tipos de neuronas
Existen diferentes tipos de neuronas, y el papel funcional de una neurona determinada depende íntimamente de su estructura. Existe una asombrosa diversidad de formas y tamaños de neuronas que se encuentran en diferentes partes del sistema nervioso (y entre especies), como lo ilustran las neuronas que se muestran en la Figura 7.6.

Si bien hay muchos subtipos de células neuronales definidos, las neuronas se dividen ampliamente en cuatro tipos básicos: unipolares, bipolares, multipolares y pseudounipolares. La Figura 7.7 ilustra estos cuatro tipos de neuronas básicas. Las neuronas unipolares tienen una sola estructura que se extiende alejándose del soma. Estas neuronas no se encuentran en los vertebrados sino que se encuentran en insectos donde estimulan músculos o glándulas. Una neurona bipolar tiene un axón y una dendrita que se extiende desde el soma. Un ejemplo de una neurona bipolar es una célula bipolar retiniana, que recibe señales de células fotorreceptoras que son sensibles a la luz y transmite estas señales a células ganglionares que transportan la señal al cerebro. Las neuronas multipolares son el tipo de neurona más común. Cada neurona multipolar contiene un axón y múltiples dendritas. Las neuronas multipolares se pueden encontrar en el sistema nervioso central (cerebro y médula espinal). Un ejemplo de una neurona multipolar es una célula de Purkinje en el cerebelo, que tiene muchas dendritas ramificadas pero sólo un axón. Las células pseudounipolares comparten características tanto con células unipolares como bipolares. Una célula pseudounipolar tiene un proceso único que se extiende desde el soma, como una célula unipolar, pero este proceso posteriormente se ramifica en dos estructuras distintas, como una célula bipolar. La mayoría de las neuronas sensoriales son pseudounipolares y tienen un axón que se ramifica en dos extensiones: una conectada a dendritas que reciben información sensorial y otra que transmite esta información a la médula espinal.

Neurogénesis
En un momento, los científicos creían que las personas nacieron con todas las neuronas que alguna vez tendrían. Investigaciones realizadas durante las últimas décadas indican que la neurogénesis, el nacimiento de nuevas neuronas, continúa hasta la edad adulta. La neurogénesis se descubrió por primera vez en pájaros cantores que producen nuevas neuronas mientras aprenden canciones. Para los mamíferos, las nuevas neuronas también juegan un papel importante en el aprendizaje: alrededor de 1000 nuevas neuronas se desarrollan en el hipocampo (una estructura cerebral involucrada en el aprendizaje y la memoria) cada día. Si bien la mayoría de las nuevas neuronas morirán, los investigadores encontraron que un aumento en el número de nuevas neuronas supervivientes en el hipocampo se correlacionaba con lo bien que las ratas aprendieron una nueva tarea. Curiosamente, tanto el ejercicio como algunos medicamentos antidepresivos también promueven la neurogénesis en el hipocampo. El estrés tiene el efecto contrario. Si bien la neurogénesis es bastante limitada en comparación con la regeneración en otros tejidos, la investigación en esta área puede conducir a nuevos tratamientos para trastornos como el Alzheimer, el accidente cerebrovascular y la epilepsia.
¿Cómo identifican los científicos nuevas neuronas? Un investigador puede inyectar un compuesto llamado bromodeoxiuridina (BrdU) en el cerebro de un animal. Si bien todas las células estarán expuestas a BrdU, BrdU solo se incorporará al ADN de células recién generadas que se encuentran en fase S. Se puede utilizar una técnica llamada inmunohistoquímica para unir un marcador fluorescente al BrdU incorporado, y un investigador puede utilizar la microscopía fluorescente para visualizar la presencia de BrdU, y así nuevas neuronas, en el tejido cerebral. La Figura 7.8 es una micrografía que muestra neuronas marcadas fluorescentemente en el hipocampo de una rata.

Transmisión de impulsos nerviosos dentro de una neurona
Para que el sistema nervioso funcione, las neuronas deben poder enviar y recibir señales. Estas señales son posibles porque cada neurona tiene una membrana celular cargada (una diferencia de voltaje entre el interior y el exterior), y la carga de esta membrana puede cambiar en respuesta a las moléculas de neurotransmisores liberadas de otras neuronas y estímulos ambientales. Para entender cómo se comunican las neuronas, primero hay que entender la base de la carga de membrana basal o 'en reposo'.
Membranas con carga neuronal
La membrana bicapa lipídica que rodea a una neurona es impermeable a moléculas o iones cargados. Para entrar o salir de la neurona, los iones deben pasar a través de proteínas especiales llamadas canales iónicos que abarcan la membrana. Los canales iónicos tienen diferentes configuraciones: abierto, cerrado e inactivo, como se ilustra en la Figura 7.9. Algunos canales iónicos necesitan ser activados para abrirse y permitir que los iones pasen dentro o fuera de la célula. Estos canales iónicos son sensibles al medio ambiente y pueden cambiar su forma en consecuencia. Los canales iónicos que cambian su estructura en respuesta a los cambios de voltaje se denominan canales iónicos regulados por voltaje. Los canales iónicos regulados por voltaje regulan las concentraciones relativas de diferentes iones dentro y fuera de la célula. La diferencia en la carga total entre el interior y el exterior de la célula se denomina potencial de membrana.

Este video analiza la base del potencial de la membrana en reposo.
Potencial de membrana en reposo
Una neurona en reposo está cargada negativamente: el interior de una célula es aproximadamente 70 milivoltios más negativo que el exterior (−70 mV, tenga en cuenta que este número varía según el tipo de neurona y por especie). Este voltaje se denomina potencial de membrana en reposo; es causado por diferencias en las concentraciones de iones dentro y fuera de la célula. Si la membrana fuera igualmente permeable a todos los iones, cada tipo de ion fluiría a través de la membrana y el sistema alcanzaría el equilibrio. Debido a que los iones no pueden simplemente cruzar la membrana a voluntad, existen diferentes concentraciones de varios iones dentro y fuera de la célula, como se muestra en la Tabla 7.1.
| Ion | Concentración extracelular (mM) | Concentración intracelular (mM) | Relación exterior/interior |
|---|---|---|---|
| Na+ | 145 | 12 | 12 |
| K+ | 4 | 155 | 0.026 |
| Cl− | 120 | 4 | 30 |
| Aniones orgánicos (A−) | — | 100 |
La diferencia en el número de iones de potasio cargados positivamente (K+) dentro y fuera de la célula domina el potencial de membrana en reposo (Figura 7.10). Cuando la membrana está en reposo, los iones K+ se acumulan dentro de la célula debido a un movimiento neto con el gradiente de concentración. El potencial negativo de la membrana en reposo se crea y mantiene aumentando la concentración de cationes fuera de la célula (en el fluido extracelular) en relación con el interior de la célula (en el citoplasma). La carga negativa dentro de la célula es creada por la membrana celular siendo más permeable al movimiento de iones potasio que al movimiento de iones sodio. En las neuronas, los iones potasio se mantienen a altas concentraciones dentro de la célula mientras que los iones de sodio se mantienen a altas concentraciones fuera de la célula. La célula posee canales de fuga de potasio y sodio que permiten que los dos cationes se difundan por su gradiente de concentración. Sin embargo, las neuronas tienen muchos más canales de fuga de potasio que los canales de fuga de sodio. Por lo tanto, el potasio se difunde fuera de la celda a un ritmo mucho más rápido que el sodio se filtra. Debido a que más cationes están saliendo de la celda de los que están entrando, esto hace que el interior de la celda se cargue negativamente en relación con el exterior de la celda. Las acciones de la bomba de sodio-potasio ayudan a mantener el potencial de reposo, una vez establecido. Recordemos que las bombas de sodio y potasio traen dos iones K+ a la célula mientras eliminan tres iones Na+ por ATP consumido. A medida que se expulsan más cationes de la célula que los absorbidos, el interior de la célula permanece cargado negativamente en relación con el líquido extracelular. Cabe señalar que los iones calcio (Cl—) tienden a acumularse fuera de la célula porque son repelidos por proteínas cargadas negativamente dentro del citoplasma.

Potencial de acción
Una neurona puede recibir entrada de otras neuronas y, si esta entrada es lo suficientemente fuerte, enviar la señal a las neuronas aguas abajo. La transmisión de una señal entre neuronas generalmente es transportada por un químico llamado neurotransmisor. La transmisión de una señal dentro de una neurona (de dendrita a axón terminal) es transportada por una breve inversión del potencial de membrana en reposo llamado potencial de acción. Cuando las moléculas de neurotransmisores se unen a receptores localizados en las dendritas de una neurona, los canales iónicos se abren. En las sinapsis excitatorias, esta apertura permite que los iones positivos ingresen a la neurona y da como resultado la despolarización de la membrana, una disminución en la diferencia de voltaje entre el interior y el exterior de la neurona. Un estímulo de una célula sensorial u otra neurona despolariza la neurona diana a su potencial umbral (-55 mV). Los canales de Na+ en el montecillo axónico se abren, permitiendo que los iones positivos ingresen a la célula (Figura 7.10 y Figura 7.11). Una vez que los canales de sodio se abren, la neurona se despolariza completamente a un potencial de membrana de aproximadamente +40 mV. Los potenciales de acción se consideran un evento de “todo o nada”, en el sentido de que, una vez que se alcanza el potencial umbral, la neurona siempre se despolariza por completo. Una vez que se completa la despolarización, la célula ahora debe “restablecer” su voltaje de membrana de nuevo al potencial de reposo. Para lograrlo, los canales de Na+ cierran y no se pueden abrir. Esto inicia el periodo refractario de la neurona, en el que no puede producir otro potencial de acción porque sus canales de sodio no se abrirán. Al mismo tiempo, los canales K+ activados por voltaje se abren, lo que permite que K+ salga de la celda. A medida que los iones K+ salen de la célula, el potencial de membrana vuelve a ser negativo. La difusión de K+ fuera de la célula en realidad hiperpolariza la célula, ya que el potencial de membrana se vuelve más negativo que el potencial de reposo normal de la célula. En este punto, los canales de sodio volverán a su estado de reposo, lo que significa que están listos para abrirse nuevamente si el potencial de membrana vuelve a superar el potencial umbral. Finalmente, los iones K+ adicionales se difunden fuera de la célula a través de los canales de fuga de potasio, llevando a la célula de su estado hiperpolarizado, de vuelta a su potencial de membrana en reposo.

En resumen, un potencial de acción es causado por movimientos de iones a través de la membrana celular como se muestra. La despolarización ocurre cuando un estímulo hace que la membrana sea permeable a los iones. La repolarización sigue a medida que la membrana vuelve a ser impermeable
los iones. La repolarización sigue a medida que la membrana vuelve a ser impermeable y
y se mueve de alta a baja concentración. A largo plazo, el transporte activo mantiene lentamente las diferencias de concentración, pero la célula puede disparar cientos de veces en rápida sucesión sin agotarlas seriamente.
se mueve de alta a baja concentración. A largo plazo, el transporte activo mantiene lentamente las diferencias de concentración, pero la célula puede disparar cientos de veces en rápida sucesión sin agotarlas seriamente.
 ) a través de la única membrana de 8 nm-espesor es inmenso (¡del orden de 11 mV/m!) y tiene efectos fundamentales sobre su estructura y permeabilidad. Ahora bien, si se toma que el exterior de una neurona esté a 0 V, entonces el interior tiene un potencial de reposo de aproximadamente —90 mV. Dichos voltajes se crean a través de las membranas de casi todos los tipos de células animales pero son más grandes en células nerviosas y musculares. De hecho, el 25% total de la energía utilizada por las células se destina a crear y mantener estos potenciales. y. Eventualmente, la célula debe reponer estos iones para mantener las diferencias de concentración que crean bioelectricidad. Esta bomba de sodio-potasio es un ejemplo de transporte activo, donde la energía celular se utiliza para mover iones a través de membranas contra gradientes de difusión y la fuerza de Coulomb.
) a través de la única membrana de 8 nm-espesor es inmenso (¡del orden de 11 mV/m!) y tiene efectos fundamentales sobre su estructura y permeabilidad. Ahora bien, si se toma que el exterior de una neurona esté a 0 V, entonces el interior tiene un potencial de reposo de aproximadamente —90 mV. Dichos voltajes se crean a través de las membranas de casi todos los tipos de células animales pero son más grandes en células nerviosas y musculares. De hecho, el 25% total de la energía utilizada por las células se destina a crear y mantener estos potenciales. y. Eventualmente, la célula debe reponer estos iones para mantener las diferencias de concentración que crean bioelectricidad. Esta bomba de sodio-potasio es un ejemplo de transporte activo, donde la energía celular se utiliza para mover iones a través de membranas contra gradientes de difusión y la fuerza de Coulomb.
El potencial de acción es un pulso de voltaje en una ubicación en una membrana celular. ¿Cómo se transmite a lo largo de la membrana celular, y en particular por un axón, como impulso nervioso? La respuesta es que el voltaje cambiante y los campos eléctricos afectan la permeabilidad de la membrana celular adyacente para que allí se lleve a cabo el mismo proceso. La membrana adyacente se despolariza, afectando a la membrana más abajo, y así sucesivamente, como se ilustra en la Figura 7.6. Así, el potencial de acción estimulado en una ubicación desencadena un impulso nervioso que se mueve lentamente (aproximadamente 1 m/s) a lo largo de la membrana celular.
Ejercicio\(\PageIndex{1}\)
Los bloqueadores de los canales de potasio, como la amiodarona y la procainamida, que se utilizan para tratar la actividad eléctrica anormal en el corazón, llamada disritmia cardíaca, impiden el movimiento de K+ a través de canales de K+ controlados por voltaje. ¿Qué parte del potencial de acción esperarías que afectaran los canales de potasio? Explique por qué.
- Responder
-
Agrega el texto de respuesta aquí y automáticamente se ocultará si tienes una plantilla de “AutoNum” activa en la página.
 y
y atravesando la membrana, equivale a una onda de carga que se mueve a lo largo del exterior y el interior de la membrana.
atravesando la membrana, equivale a una onda de carga que se mueve a lo largo del exterior y el interior de la membrana.Figura 7.13. El potencial de acción se conduce por el axón a medida que la membrana del axón se despolariza y luego se vuelve a polarizar.

notas
- Este video presenta una visión general de un potencial de acción.
La mielina y la propagación del potencial de acción
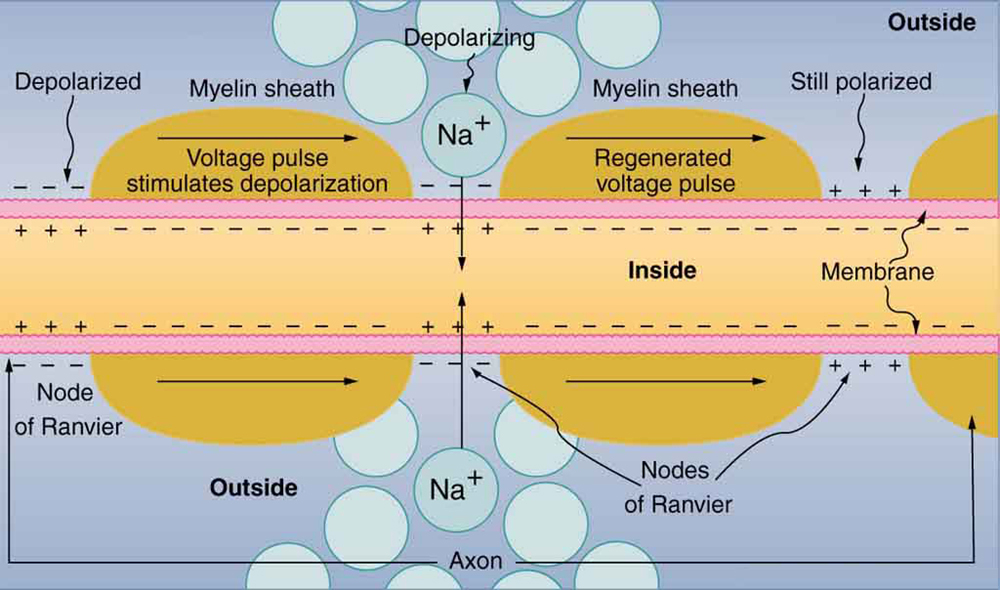
Para que un potencial de acción comunique información a otra neurona, debe viajar a lo largo del axón y llegar a las terminales del axón donde puede iniciar la liberación de neurotransmisores. La velocidad de conducción de un potencial de acción a lo largo de un axón está influenciada tanto por el diámetro del axón como por la resistencia del axón a la fuga de corriente. La mielina actúa como aislante que impide que la corriente salga del axón; esto aumenta la velocidad de conducción del potencial de acción. En enfermedades desmielinizantes como la esclerosis múltiple, la conducción del potencial de acción se ralentiza debido a que la corriente se filtra desde áreas axónicas previamente aisladas. Los nodos de Ranvier, ilustrados en la Figura 7.15, son huecos en la vaina de mielina a lo largo del axón. Estos espacios no mielinizados tienen aproximadamente un micrómetro de largo y contienen canales de Na+ y K+ regulados por voltaje. El flujo de iones a través de estos canales, particularmente los canales de Na+, regenera el potencial de acción una y otra vez a lo largo del axón. Este 'salto' del potencial de acción de un nodo al siguiente se llama conducción saltatoria. Si los nodos de Ranvier no estuvieran presentes a lo largo de un axón, el potencial de acción se propagaría muy lentamente ya que los canales Na+ y K+ tendrían que regenerar continuamente los potenciales de acción en cada punto a lo largo del axón en lugar de en puntos específicos. Los nodos de Ranvier también ahorran energía para la neurona ya que los canales solo necesitan estar presentes en los nodos y no a lo largo de todo el axón.

La Figura 7.16 muestra una vista ampliada de un axón que tiene vainas de mielina separadas característicamente por huecos no mielinizados (llamados nodos de Ranvier). Esta disposición le da al axón una serie de propiedades interesantes. Dado que la mielina es un aislante, evita que las señales salten entre nervios adyacentes (diafonía). Adicionalmente, las regiones mielinizadas transmiten señales eléctricas a una velocidad muy alta, como lo haría un conductor o resistor ordinario. No hay potencial de acción en las regiones mielinizadas por lo que no se utiliza energía celular en ellas. Hay una pérdida de señal en la mielina, pero la señal se regenera en los huecos, donde el pulso de voltaje activa el potencial de acción a voltaje completo. Por lo que un axón mielinado transmite un impulso nervioso más rápido, con menos consumo de energía, y está mejor protegido de la diafonía que uno no mielinizado. No todos los axones están mielinizados por lo que la diafonía y la transmisión lenta de la señal son una característica del funcionamiento normal de estos axones, otra variable en el sistema nervioso.
señal en la mielina, pero la señal se regenera en los huecos, donde el pulso de voltaje activa el potencial de acción a voltaje completo. Por lo que un axón mielinado transmite un impulso nervioso más rápido, con menos consumo de energía, y está mejor protegido de la diafonía que uno no mielinizado. No todos los axones están mielinizados por lo que la diafonía y la transmisión lenta de la señal son una característica del funcionamiento normal de estos axones, otra variable en el sistema nervioso.
La degeneración o destrucción de las vainas de mielina que rodean las fibras nerviosas perjudica la transmisión de la señal y puede provocar numerosos efectos neurológicos. Una de las enfermedades más destacadas proviene del propio sistema inmunológico del cuerpo que ataca a la mielina en el sistema nervioso central, la esclerosis múltiple. Los síntomas de la EM incluyen fatiga, problemas de visión, debilidad de brazos y piernas, pérdida de equilibrio y hormigueo o entumecimiento en las extremidades (neuropatía). Es más apto para golpear a los adultos más jóvenes, especialmente a las hembras. Las causas pueden provenir de infecciones, efectos ambientales o geográficos, o genética. Por el momento no hay cura conocida para la EM.
La mayoría de las células animales pueden disparar o crear su propio potencial de acción. Las células musculares se contraen cuando se disparan y a menudo son inducidas a hacerlo por un impulso nervioso. De hecho, las células nerviosas y musculares son fisiológicamente similares, e incluso hay células híbridas, como en el corazón, que tienen características tanto de nervios como de músculos. Algunos animales, como la infame anguila eléctrica (Figura 7.17) utilizan músculos agrupados para que sus voltajes se agreguen con el fin de crear un choque lo suficientemente grande como para aturdir a las presas.
Propagación de un impulso nervioso por un axón mielinizado, de izquierda a derecha. La señal viaja muy rápido y sin entrada de energía en las regiones mielinizadas, pero pierde voltaje. Se regenera en las brechas. La señal se mueve más rápido que en los axones no mielinizados y está aislada de las señales en otros nervios, limitando la diafonía.

Ejercicio\(\PageIndex{1}\)
¿Cuál de las siguientes afirmaciones es falsa?
a. El soma es el cuerpo celular de una célula nerviosa.
b. La vaina de mielina proporciona una capa aislante a las dendritas.
c. Los axones llevan la señal del soma al objetivo.
d. Las dendritas llevan la señal al soma.
Ejercicio\(\PageIndex{2}\)
Las neuronas contienen ________, que puede recibir señales de otras neuronas.
a. axones
b. mitocondrias
c. dendritas
d. cuerpos de Golgi
Ejercicio\(\PageIndex{3}\)
Una neurona (n) ________ tiene un axón y una dendrita que se extiende directamente desde el cuerpo celular.
a. unipolar
b. bipolar
c. multipolar
d. pseudounipolar
Ejercicio\(\PageIndex{4}\)
Las glías que proporcionan mielina para las neuronas en el cerebro se llaman ________.
a. Células de Schwann
b. oligodendrocitos
c. microglía
d. astrocitos
Ejercicio\(\PageIndex{5}\)
¿Cómo son las neuronas similares a otras células? ¿Cómo son únicos?
Ejercicio\(\PageIndex{6}\)
¿Comparar y contrastar el potencial de descanso, gradación y acción? En tu respuesta, asegúrate de haber incluido canales y referencia de voltaje así como estructuras relevantes de las neuronas. Una vez que se te ocurra una respuesta, dásela a otro alumno para que la revise. Con base en la revisión de tu compañero, ¿hay algo en lo que necesites trabajar en términos de tu comprensión del potencial de descanso, grado y acción?
Ejercicio\(\PageIndex{7}\)
La esclerosis múltiple provoca desmielinización de axones en el cerebro y la médula espinal. ¿Por qué es esto problemático?