
1.6: Desarrollo Temprano
- Page ID
- 121772

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
- Visión general del desarrollo temprano
- Patrones
- Primer trimestre:
- Preimplantación
- Gastrulación
- Diferenciación del ectodermo
- Neurulación
- Migración de la cresta neural
- Diferenciación del mesodermo
- Formación de somitas
- Formación del corazón
- Diferenciación del endodermo
- Formación de faringe
- Desarrollo de otras estructuras de la cavidad oral
- Desarrollo de las glándulas salivales
- Desarrollo de papilas linguales
- Aplicaciones clínicas del desarrollo temprano

Visión general del desarrollo temprano
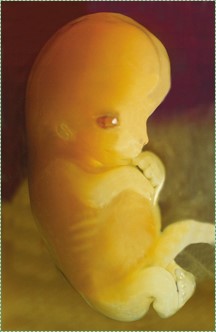
El desarrollo humano es el proceso en el que una sola célula, una célula visualmente poco notable excepto quizás por su tamaño, cambia lentamente a billones (37) de células de aspecto diferente. La embriología es el estudio de las fases del desarrollo, especialmente las fases tempranas donde ocurre la mayoría de las cosas interesantes. Una célula redonda, solo una bola sin lados y sin partes especiales, se convierte en un organismo con diferentes órganos en diferentes lugares. La Fig. 6.1 es el Sol. Lo ves todos los días, ¿puedes decir si la foto del lado derecho hacia arriba? ¿Hay siquiera un lado derecho arriba? No somos astrónomos, pero el Sol se parece un poco a un óvulo fertilizado, y en la Fig. 6.1 no hay hitos obvios para orientar nuestra visión. Un huevo tiene una apariencia similar amorfa (sin forma), pero se transforma (cambia suavemente de forma) en el embrión que se muestra en la Fig. 6.2, que tiene un extremo de cabeza y cola, y claros comienzos de brazos y piernas. La primera parte de este capítulo se centra en conceptos comunes a muchos eventos de desarrollo vistos en los capítulos 7 al 11. La segunda mitad del capítulo abarca los detalles del desarrollo temprano.
El embarazo se divide en tres periodos de tiempo iguales denominados trimestres. Durante el primer trimestre, una sola célula se multiplica en un organismo multicelular sin forma discernible (aparte de ser una mancha redonda, no un icosoedro). A partir de la semana 2, la gota de células se convierte en un embrión, que tiene diferentes formas. El embrión sigue desarrollando nuevas formas hasta que tiene todas las formas humanas básicas, como la cabeza y los brazos y el corazón. En este momento (semana 8) es feto, y el primer trimestre ha terminado. Los dos trimestres siguientes son relativamente aburridos, el feto crece más grande y las formas maduran, pero nada terriblemente emocionante pasa hasta el nacimiento. Por eso se trata de un libro de Histología y Embriología, no de Histología y Fetología.


A medida que las células humanas se dividen durante el primer trimestre, toman decisiones en cuanto a su destino celular, lo que finalmente determina el número y ubicación de nuestros brazos y piernas, nuestra cabeza y corazón. Estas decisiones implican que se envíen señales entre celdas. Los errores en estas señales o decisiones conducen a trastornos congénitos (o malformaciones congénitas, o defectos de nacimiento (pero ver más abajo)). Estos pueden ocurrir sin causa conocida. O bien, los químicos pueden interferir con las señales de desarrollo, y a estos los llamamos teratógenos. El teratógeno más famoso de mi país es el medicamento talidomida, que —si es tomado por mujeres embarazadas en el momento equivocado— interfiere con los procesos de toma de decisiones que usa un embrión para decidir dónde crecer las extremidades. Esto provoca focomelia, o malformación de las extremidades. No todos los países comparten este sesgo, los efectos teratogénicos del agente naranja (dioxina, que también se encuentra en el humo del cigarrillo) y la radiación atómica (descarga pdf) están notablemente ausentes en la mayoría de las clases escolares estadounidenses (agente naranja y radiación atómica también tienen efectos adversos en adultos, y la tasa que esta información llega al público estadounidense está sujeta a sesgos similares). Cubriremos un pequeño número de señales involucradas en algunas decisiones importantes que se toman durante el desarrollo facial y dental. Se toman 37 billones de decisiones entre la etapa unicelular y el nacimiento, los errores son inevitables. Algunos errores son menores, cambiando la apariencia pero no la función de un órgano; podríamos referirnos a esto como una marca de nacimiento o variación humana. Otros errores son más severos, interfieren con la salud de una persona, y nos referimos a esos como trastornos congénitos (o quizás errores congénitos de desarrollo, pero preferiblemente no defectos congénitos, porque nuestra elección de palabras puede influir en la cantidad de atención médica que la gente busca, depende de el paciente). Si en algún momento sientes que necesitas más información sobre embriología, estos enlaces son útiles:
- La página de Educación en Embriología
- por el Dr. Mark Hill en la Universidad de Nueva Gales del Sur, Sydney, Australia
- Imágenes embrionarias Desarrollo normal y anormal de mamíferos
- por la Dra. Kathleen K. Sulik y el Dr. Peter R. Bream Jr. en la Facultad de Medicina de la Universidad de Carolina del Norte.
- El Atlas 3D de la Embriología Humana
- una colección de código abierto de archivos PDF 3D (legibles usando Adobe Acrobat Reader) por más de 70 estudiantes diferentes en el Academic Medical Center, Países Bajos.
Patrones
Esta sección abarca los procesos básicos que ocurren durante el desarrollo. A menudo, más de un proceso está ocurriendo en el mismo lugar al mismo tiempo. Sin embargo, es útil pensar en los siguientes procesos por separado, la forma en que podrías enfocarte solo en las voces —o violonchelo, o guitarra, o acordeón— de una canción.

Clonación
La mitosis produce clones, o células idénticas con ADN idéntico. Sin embargo, las células se diferencian y expresan diferentes genes para convertirse en diferentes tejidos y órganos. En las primeras 2 semanas de desarrollo, las células se dividen y producen células más idénticas (aunque más pequeñas). Muy pronto, las células toman decisiones en cuanto a lo que van a convertirse, y estas decisiones se coordinan con otras células. Sin coordinación, un embrión podría terminar con dos colillas y sin cabeza.

Diferenciación
La diferenciación es el proceso donde las células madre comienzan a expresar diferentes genes, haciendo que aparezcan y funcionen de manera diferente a otras células. Todas las células del cuerpo humano tienen el mismo ADN, porque todas las células copiaron su ADN del ADN en el cigoto (algunas células excepcionales cambian un poco su ADN). El cigoto es una célula madre totipotente, los factores de transcripción podrían unirse y activar cualquier gen. A medida que las células se diferencian, sin embargo, limitan su destino celular al poner algunos genes en almacenamiento profundo (no cambiando realmente el código genético, sino cambiando lo que puede o no ser accesible para la ARN polimerasa).

La diferenciación se desencadena al recibir señales químicas. Estas señales hacen que las células diana metilen el ADN de ciertos genes no necesarios, empaqueten esas regiones de ADN alrededor de las histonas y cierren permanentemente esos genes. Por ejemplo, las células destinadas a convertirse en ovarios nunca sintetizan insulina, el gen de la insulina se empaqueta mucho antes de que los ovarios secreten estrógeno u ovulen. La metilación del ADN y el empaquetamiento de histonas limitan el destino de estas células. Después de eso, las señales pueden desaparecer porque el patrón de metilación se copia durante la mitosis y se pasa a ambas células hijas (y sus células hijas). En la Fig. 6.7, imagínese que una señal instruye a una célula a metilar genes verdes, dejando los genes rojos abiertos para ser potencialmente transcritos. Una señal diferente hace lo contrario a la otra celda. A medida que el glóbulo rojo duplica su ADN, copia el patrón de metilación, de manera que todas sus células hijas no pueden expresar genes verdes. En tanto, la célula verde produce una serie de células hijas que tienen el gen rojo metilado, ninguna de sus células hijas puede expresar el gen rojo. Ahora tenemos 4 glóbulos rojos, que pertenecen a un linaje, y 4 células verdes que pertenecen a un linaje diferente. Surge un patrón adicional a medida que las células transcriben genes no metilados. En la caricatura, a medida que los glóbulos rojos transcriben el gen rojo, esto crea una señal de corto alcance que induce a las células verdes cercanas a transcribir el gen verde oscuro. Diríamos que esas células diana se han diferenciado aún más, y este organismo se ha vuelto aún más complicado (3 tipos celulares diferentes). Esto debe continuar hasta que tengamos aproximadamente 200 tipos de células diferentes. En términos generales, a medida que una célula se vuelve más especializada, su destino se vuelve más limitado (más metilación del ADN y empaquetamiento de histonas), posiblemente hasta el punto de limitarse a un solo destino celular.

Inducción
Una célula puede señalizar a otra célula e inducir (instruye) a esa célula para que se diferencie. Los químicos que pueden inducir a otra célula a cambiar su forma (a transformarse) y cambiar el destino celular se denominan morfógenos (o a veces llamados Factores de Crecimiento). Los morfógenos son similares a los neurotransmisores u hormonas, a menudo se unen a las proteínas receptoras de la superficie celular. La activación de los receptores de morfógenos conduce a la activación de señales intracelulares, que a su vez pueden activar o inhibir factores de transcripción específicos para alterar la expresión de diferentes genes. Los cambios en la expresión génica hacen que la célula diana se vea y funcione de manera diferente. Los morfógenos también cambian la metilación de los genes y su empaquetamiento alrededor de las histonas, lo que es un cambio más permanente en la expresión génica. Este patrón de regulación génica se transmite a las células hijas, lo que aprendimos en el Capítulo 1 es un ejemplo de herencia epigenética. Al regular la expresión génica a través de la actividad del factor de transcripción y metilación, los morfógenos cambian no solo la morfología de la célula diana, sino también su destino celular. A menudo hay varias moléculas estrechamente relacionadas que pertenecen a una familia de morfogenos, cada uno de los cuales tiene un nombre como morfogen-1, morfogen-2a, morfogen-2b y morfogen-3. Hay una gran cantidad de morfógenos diferentes, en este libro de texto discutimos solo unos pocos por su nombre, en su mayoría los relacionados directamente con el desarrollo dental, incluyendo la familia de la Proteína Morfogénica Ósea (BMP), la familia Wnt y la familia del Factor de Crecimiento de Fibroblastos (FGF). Los morfógenos pueden actuar sobre diferentes distancias enumeradas en el Cuadro 6.1. Los nombres de morfógenos específicos son muy importantes en los exámenes de embriología, y mucho menos en los exámenes de higiene dental.
| Distancia | Tipo | Ejemplos |
|---|---|---|
| Directo | Celda a celda | Células madre epiteliales a neuromesenquimales (diferenciación de odontoblastos) |
| Celda a ECM | Integrina a colágeno (diferenciación de Ameloblastos) | |
| Corto alcance | Difusión por la sustancia molida | Proteína Morfogénica Ósea (BMP), Wnt, Factor de Crecimiento de Fibroblastos (FGF). (Inducción del brote dental) |
| Señales de todo el cuerpo | Hormonas | Hormona del Crecimiento, Calcitonina. |
Cuadro 6.1: Ejemplos de diferentes tipos de señales observadas durante la embriogénesis.
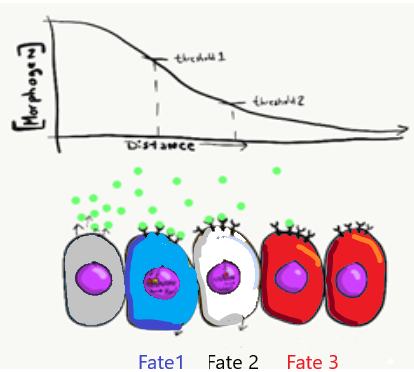
Los morfógenos se secretan a la sustancia molida, creando un gradiente de concentración de los niveles de morfógeno a medida que se difunden lentamente lejos de la fuente. La cantidad de moléculas de morfógeno producidas, la pegajosidad relativa de la sustancia fundamental, la distancia de la fuente y el número de proteínas receptoras en la célula diana determinan el nivel de una señal que reciben las células. A veces, un morfógeno produce efectos visibles que caen a lo largo de un gradiente (o espectro), que conceptualmente coincide con los gradientes que acabamos de enumerar, como el color de la piel (más claro a más oscuro) o el tamaño del cuerpo (más corto a más alto). Sin embargo, a veces los efectos de un morfógeno son más discretos (brote dental o ningún brote dental, no hay brotes de medio diente). Para ayudar a explicar estas observaciones se requiere un poco de conocimiento de cómo funcionan los factores de transcripción. Por ejemplo, en el modelo de bandera francesa (Fig 141), imagínese que recibir altos niveles de la señal de morfógeno desencadena la transcripción de un gen azul e inhibe un gen rojo, provocando que esas células se diferencien en células azules. Cantidades moderadas de la señal de morfógeno pueden no ser suficientes para reclutar ARN polimerasa para el gen azul, pero suficientes para inhibir el gen rojo, provocando que las células en regiones de niveles moderados de morfógeno se diferencien en glóbulos blancos. Por último, las células que reciben poca o ninguna señal de morfógeno no inhiben el gen rojo, lo que hace que se diferencien en glóbulos rojos. Hay otros modelos que ayudan a explicar el vínculo entre la inducción y la morfogénesis —entre una sustancia química y un organismo— por ejemplo, un campo morfogenético es menos francófilo en la naturaleza pero más difícil de visualizar. Todos estos modelos comparten un concepto importante: las señales químicas pueden comunicar información posicional en un embrión en desarrollo.
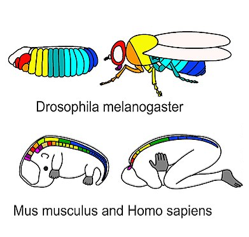
Durante un desarrollo muy temprano, la expresión regional de genes homeobox controla el destino celular, no la morfología. Los genes homeobox son factores de transcripción que especifican de qué parte del cuerpo forma parte ahora una célula en particular, como parte de un brazo o cara, aunque esa área aún no tenga una forma específica. No codifican proteínas, hacen que las células se conviertan en parte de un órgano específico. En cambio, los genes homeobox activan o inhiben una colección de otros genes (un programa), estableciendo el escenario para futuros cambios. En contraste, los morfógenos como FGF y BMP activan genes que alteran la forma de una célula o tejido (morfología). La expresión del gen homeobox representa un plan muy específico en embriología. Está altamente conservado en todas las especies animales, y el patrón es muy predecible en embriones. El patrón también es relativamente simple y lineal (Fig. 6.10). Piense en genes homeobox como topógrafos que determinan las ubicaciones donde se construirán las casas en una subdivisión, mientras que los morfógenos son los contratistas que siguen los planos de ADN para cada casa, los cuales son construidos por células. Podemos extender esta metáfora un paso más allá: en una subdivisión, las casas son similares, derivadas de un plano básico, pero dadas ligeras modificaciones con cada reutilización.
![[animar la imagen de salida]](https://med.libretexts.org/@api/deki/files/22913/animate-output-image.gif)
La capacidad de un tejido para crear sus propios patrones se llama autoorganización (en contraste, los genes de efecto materno de las células de la madre guían los patrones en las células del embrión). Esto involucra múltiples morfógenos, inicialmente expresados aleatoriamente. Las interacciones entre estos morfógenos conducen a un patrón estable de expresión y un patrón complejo de diferenciación. Por ejemplo, algunos morfógenos bloquean a otras células para que no produzcan el mismo morfógeno. En la Fig. 6.11, imagine que un morfógeno azul induce a una célula a diferenciarse en una célula azul, pero impide que las células vecinas se conviertan en células azules. Estos vecinos serían libres de diferenciarse en glóbulos rojos. Al mismo tiempo, una célula distante produce morfógeno rojo, lo que la induce a diferenciarse en un glóbulo rojo, pero impide que sus vecinos hagan lo mismo. Sus vecinos serían libres de producir morfógeno azul y diferenciarse en células azules. El patrón inicial que surge es alternar anillos de células rojas y azules. Sin embargo, los anillos producen regiones de rojo al lado del rojo y del azul al lado del azul, lo que viola nuestras reglas imaginarias, y esto inicia una batalla. Para minimizar el número de rojo siguiente a rojo y azul-próximo a azul, las células cambian su destino celular hasta que surge un patrón estable, en este caso rayas. Sería difícil predecir desde el principio qué celdas serían rojas versus azules, pero se podría predecir el desarrollo de franjas rojas y azules. Si estas fueran células productoras de pigmento en una cebra, tendríamos una cebra con rayas (el mejor tipo de cebra). Pero este patrón también podría ser regiones alternas de la mucosa oral que crecen un brote dental o no crecen un brote dental, asegurando que cada brote dental tenga suficiente espacio para crecer (si quieres bajar por una madriguera de un conejito, estas interacciones se pueden modelar matemáticamente). Cuando un morfógeno bloquea otra señal, a eso lo llamamos antagonismo, y las señales antagonistas son tan comunes como las señales de inducción. Deben, después de todo, es fundamentalmente igual de importante crecer una cabeza como lo es no crecer dos cabezas.
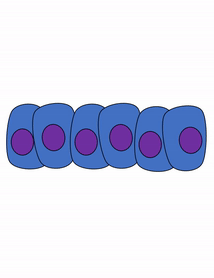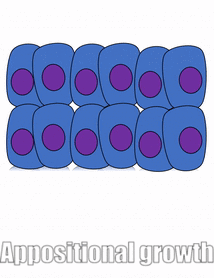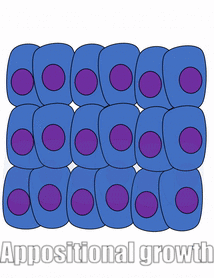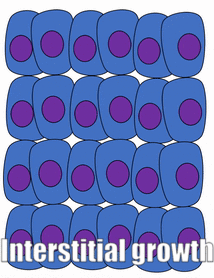
proliferación
El crecimiento de un tejido se llama proliferación. La proliferación ocurre ya sea por mitosis (adición de células) o por adición de matriz extracelular (síntesis de proteínas u otras moléculas). Cuando el nuevo crecimiento ocurre junto a una región de crecimiento más viejo, este es crecimiento aposicional, mientras que si el crecimiento ocurre desde dentro y empuja el tejido más viejo hacia afuera, ese es el crecimiento intersticial. Los tejidos densos, como el hueso, el esmalte y la dentina, experimentan crecimiento aposicional. Los tejidos más blandos como el mesénquima crecen intersticialmente.
El crecimiento de un tejido está regulado por hormonas, como la Hormona de Crecimiento, que coordina la tasa de crecimiento de los tejidos en todo el cuerpo. Por otro lado, los morfógenos de acción local hacen que las regiones pequeñas crezcan más rápido que las regiones vecinas. Para dejar espacio para nuevas células mientras se mantiene una sola capa de células conectadas a sus vecinos por desmosomas y uniones estrechas, un epitelio se abulta hacia adentro o hacia afuera si no puede ampliarse. Un crecimiento externo se conoce como brote, y un crecimiento hacia adentro es una invaginación. Los brazos y las piernas crecen de los brotes, mientras que los dientes, los pelos y el cerebro crecen a partir de invaginaciones. Ambas son formas de crecimiento intersticial.

Morfogénesis
La morfogénesis es la aparición de nuevas formas en un organismo. Significa pasar de ninguna forma a alguna forma. La palabra morfología puede significar el estudio de estas formas, o ser utilizada como adjetivo para significar la forma de algo (Por ejemplo, un paladar hendido es una oportunidad para la morfología saludable del paladar). Desde el cigoto hasta la etapa de blástula (a veces referida como las etapas germinales) la bola de células es amorfa, no tiene forma (aparte de ser una esfera). Al inicio de la gastrulación, sin embargo, el embrión sufre numerosos cambios morfológicos. Surgen nuevas formas en el embrión en desarrollo. Desarrolla un interior y un exterior, un extremo de cola y un extremo de cabeza, pronto desarrolla una izquierda y una derecha, y una parte delantera y trasera. Después de eso, se hacen visibles un sistema nervioso y un sistema circulatorio. ¿Cuánto tiempo crees que pasa antes de que se desarrollen los dientes? La respuesta puede sorprenderte.
 Figura 6.15: D'Arcy Wentworth Thompson propuso que lo que pueden parecer diferentes formas en especies relacionadas pueden ser transformaciones de un patrón básico. Crédito de imagen: “Fig 150-153” de D'Arcy Wentworth Thompson, Sobre el crecimiento y la forma es de dominio público, CC0
Figura 6.15: D'Arcy Wentworth Thompson propuso que lo que pueden parecer diferentes formas en especies relacionadas pueden ser transformaciones de un patrón básico. Crédito de imagen: “Fig 150-153” de D'Arcy Wentworth Thompson, Sobre el crecimiento y la forma es de dominio público, CC0La Fig. 6.14 es un famoso dibujo de diminutas criaturas marinas con exoesqueletos mineralizados. Esta imagen fue analizada posteriormente por un biólogo matemático llamado D'Arcy Wentworth Thompson, quien notó que diferentes formas biológicas pueden ser diferentes transformaciones de la misma forma. Las transformaciones, propuso, ocurrieron por diferentes tasas localizadas de crecimiento —algo que nuestro conocimiento actual de la señalización de morfógenos respalda— como lo ilustra en la Fig 6.15. Se le considera como la primera persona en describir la morfogénesis. Cuando Thompson miró a las diminutas criaturas marinas, no vio diferentes formas, vio cambios a una forma. Estos cambios ocurrieron a lo largo del tiempo evolutivo, y llevaron a criaturas marinas de aspecto diferente (que compartían un solo linaje). En este libro, observamos diferentes partes del cuerpo que comparten el mismo linaje pero cambian con el tiempo de desarrollo, como los folículos pilosos y los brotes dentales. Las ideas de Thompson fueron ampliadas por el matemático Alan Turing, quien acuñó el término morfogénesis. Sus otros logros incluyen inventar la informática y romper el código nazi durante la Segunda Guerra Mundial (con la ayuda de muchas mujeres matemáticas en el parque Bletchley). Lamentablemente, se suicidó tras su arresto, condena y encarcelamiento por homosexualidad por parte del gobierno británico al que ayudó a salvar. La visión de su gobierno sobre la homosexualidad se ha desarrollado desde entonces, podemos entender mejor la visión actual siguiendo su linaje.
Morfogénesis y genética
Para tener una comprensión completa de cómo funciona la señalización de morfógenos, ayuda a comprender algunos conceptos de genética. Cuando decimos entender, no nos referimos a meorizar los detalles específicos, enfocarnos en los conceptos. Primero, preste atención a que cuando discutimos un morfógeno, probablemente nos referimos a la molécula de señalización en sí. A veces discutimos la señal de morfógeno, y esa señal de palabra extra hace una gran diferencia. Las mutaciones que alteran las señales de morfógeno incluyen mutaciones en el gen para la propia molécula de morfógeno extracelular (si es una proteína como FGF y BMP) o enzimas que sintetizan el morfógeno (como es el caso del ácido retinoico), receptores de superficie celular para los morfógenos, segundos mensajeros citoplásmicos y factores de transcripción nuclear. Discutir los niveles de señalización de morfógenos nos salva de la complejidad de incluir todos los objetivos posibles. A continuación, algunas mutaciones podrían ser ganancia de función, como una mutación a un receptor que impide que se apague, y estas a menudo se heredan de manera autosómica dominante (solo se necesita 1 alelo defectuoso para crear una señal donde no debería estar). Alternativamente, las mutaciones de pérdida de función impiden que las proteínas funcionen correctamente, y a menudo se heredan en un patrón autosómico recesivo (se necesitan ambas copias para ser defectuosas para conducir a una pérdida de una señal donde debería estar, un alelo bueno sería suficiente). Algunas mutaciones se detienen por completo, o inician completamente, las señales de morfógeno. y en tales casos diríamos que estas mutaciones tienen penetrancia completa. Otras mutaciones podrían bloquear parcialmente o iniciar parcialmente las señales de morfógeno, y tener penetrancia parcial o incompleta, la gravedad de los síntomas recae en un espectro.

Maduración
La maduración es el proceso por el cual las nuevas formas creadas por la morfogénesis continúan desarrollándose y adquiriendo una forma más madura. Al final de la semana 9, se han formado todas las formas básicas de órganos, y decimos que el periodo embrionario ha terminado y comienzan las etapas fetales. El feto experimenta maduración, y la maduración involucra fragmentos de los procesos anteriores (otra forma de decir que es la maduración recapitula otros procesos de desarrollo).
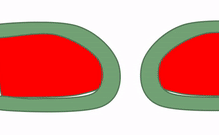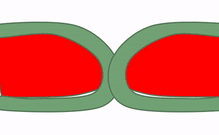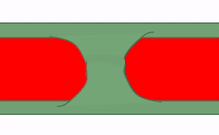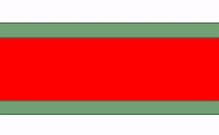
Fusión
En ocasiones, dos trozos separados del mismo tipo de tejido entran en contacto entre sí y se fusionan en un solo tejido. Para un epitelio, esto requiere que las células compartan el mismo tipo de proteína transmembrana conocida como molécula de adhesión celular. Para un tejido conectivo, esto requiere la correcta unión de integrina a la fibronectina. En la Fig 147, el patrón básico de verde en el exterior y rojo en el medio se ve interrumpido inicialmente por la fusión de dos estructuras. Hay tres formas de eliminar las células verdes del área media. La primera forma es que las células reciban señales de guía y hagan que las células se muevan. Algunas señales de orientación son repulsivas, como la falta de contactos correctos célula a célula, o altas concentraciones de un morfógeno repulsivo. Otras señales de orientación son atractivas, como el contacto correcto de célula a célula o un morfógeno de guía positivo. Si una conexión célula a célula o morfógeno es atractiva o repulsiva depende de cómo se haya programado la célula diana (qué genes receptores está expresando). Las células en la ubicación incorrecta también pueden ser inducidas a sufrir apoptosis (ver más abajo). Por último, otra forma de reorganizar las células es inducirlas a desdiferenciarse (revertir a una célula madre) y diferenciarlas en el tipo celular correcto. Esto se llama transición; tanto el desarrollo temprano como el desarrollo dental implican ejemplos de transiciones epitelial a mesenquimales y transiciones mesenquimales a epiteliales.

Apoptosis
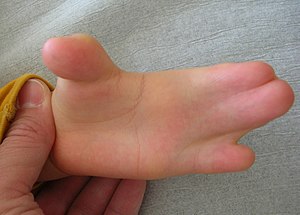
En la construcción de edificios altos o barcos grandes, los andamios se construyen primero para permitir que los trabajadores de la construcción lleguen a donde necesitan, luego se retiran después de que se complete la construcción. De igual manera, durante el desarrollo, la apoptosis es una parte integral de la construcción de partes del cuerpo. Muchos tejidos crecen más células de las necesarias en la forma madura. Para someterse a maduración, las células innecesarias sufren apoptosis. Por ejemplo, durante el crecimiento del tejido óseo, se hace que el tejido cartilaginoso adicional actúe como andamio para los osteoblastos. Se muestra en la Fig. 6.18, las manos y los pies comienzan como paletas, y solo con apoptosis en regiones alternas ocurre la morfogénesis de dedos de manos y pies. Sin suficiente apoptosis, las personas nacen con dedos de manos o pies palmeados, o sufren anquiloglosia (su lengua está rígidamente anclada al piso de la boca por un frenillo lingual pronunciado).
Desarrollo temprano

Periodo previo a la implantación
El periodo de preimplantación comienza con la fecundación, abarca aproximadamente una semana de desarrollo y termina con un embrión implantado en el útero. La fertilización es la unión de un espermatozoide y un óvulo; se necesitan miles de espermatozoides, pero solo se permite que uno entre en el óvulo. Esta unión une 23 cromosomas maternos con 23 cromosomas paternos coincidentes, formando un cigoto. La fertilización ocurre con mayor frecuencia hacia el extremo distal de una trompa de Falopio. A lo largo de la semana siguiente el cigoto sufre mitosis y aumenta en número de células, pero no crece de tamaño. Todos los aminoácidos y nucleótidos para hacer nuevas células durante esta primera semana provienen del citoplasma del cigoto, razón por la cual el huevo es una célula muy grande. Después de la implantación, el útero proporciona las materias primas para que un embrión crezca más grande.
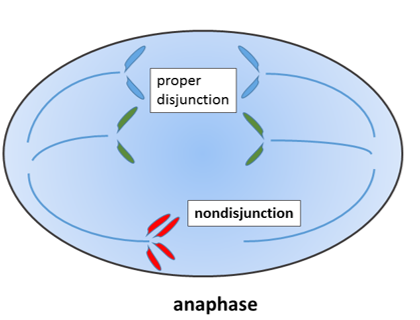
Durante la mitosis, los 23 pares de cromosomas se duplican y se dividen entre 2 células hijas de manera uniforme. Si se cometen errores y un número impar de cromosomas terminan en una célula hija, esto falla en un punto de control del ciclo celular y desencadena la apoptosis. Si un cromosoma se rompe, puede llevar al mismo resultado.

Sin embargo, hay excepciones. Una célula con un número impar de cromosomas 21 o 23 puede sobrevivir. Si esa célula es el óvulo, entonces el cigoto y todas las células posteriores tendrán un número impar de cromosomas. Esto lleva a trastornos congénitos como la Trisomía 21. Si el error de segregación cromosómica ocurre en la etapa de desarrollo de 2 células, entonces solo la mitad de las células en el adulto tendrían un cromosoma extra, y la condición sería menos grave. Los cromosomas adicionales interfieren con los procesos de toma de decisiones en el desarrollo. Las personas con Trisomía 21 desarrollan frecuentemente microdoncia y pueden sufrir hiposalivación y bruxismo. Por lo tanto, necesitan un cuidado extra manteniendo una higiene bucal saludable.

Divisiones tempranas
Durante la primera semana de desarrollo, el cigoto sufre 3 divisiones de escisión, donde la mitosis ocurre de manera sincronizada (ver un video genial aquí). Las células se duplican a lo largo de un solo plano, 3 veces, produciendo un embrión de 8 células. La sincronización de la mitosis requiere comunicación célula a célula entre uniones de brecha para coordinar los puntos de control del ciclo celular. La mitosis sincronizada lleva al embrión a desarrollarse de manera uniforme. El hecho de que múltiples células se estén dividiendo en la misma dirección significa que estas células tienen cierta polaridad, incluso si el embrión en su conjunto no tiene todavía la parte superior o inferior visibles. A medida que la mitosis continúa, el número de células sigue duplicándose, pero las células no se dividen a lo largo del mismo plano. Esta bola sólida de células se llama mórula. A medida que la mitosis continúa, el recuento de células se duplica y las células continúan haciéndose más pequeñas, hasta que la bola sólida de células se convierte en una bola hueca de células llamada blástula. Desde el estadio cigoto hasta la blástula, ninguna de las células aparece diferente excepto en tamaño. Una forma más elegante de decir esto es que no hay cambios en la morfología. Durante la etapa de blástula, un grupo de células llamadas disco embrionario se aleja de las demás. El destino del disco embrionario es convertirse en el embrión, mientras que las células externas, conocidas como el trofoblasto, están destinadas a convertirse en estructuras extra-embrionarias como la placenta. La blástula se implanta en el endometrio aproximadamente una semana después de la fecundación y continúan desarrollándose.

Gastrulación
No es nacimiento, matrimonio, o muerte, sino gastrulación, que es realmente el momento más importante de tu vida. —Lewis Wolpert.
Resumen de gastrulación
Durante la gastrulación, algunas células invaginan, formando una capa de células en el interior y una capa de células en el exterior del embrión. El significado de esto es que esta es la primera instancia donde las células embrionarias son diferentes entre sí. Pero, espera, si todas las células blástulas son idénticas, ¿por qué solo migraron algunas de ellas? De hecho, la mitosis genera 2 células con ADN idéntico, pero no necesariamente con citoplasma idéntico. Si un óvulo tiene ARNm para un factor de transcripción agrupado en una mancha en el citoplasma, la primera división de escisión produce dos células no idénticas, una con el factor de transcripción ARNm y otra sin. Posteriormente, cuando se traduce el ARNm, podría inducir a esa célula a migrar hacia adentro, mientras que otras células permanecen en el exterior. Esto es un efecto materno, porque el ARNm para el factor de transcripción no se transcribe a partir del ADN de los cigotos, sino que es hecho por células de ovario de mamá y depositado en el óvulo. Volvamos a gastrulatiojn: el resultado final de la gastrulación es un embrión con dos capas (exterior e interior). Imagínese sostener una pelota de tenis y empujar los pulgares hacia adentro, obligando a que parte de la goma exterior se pliegue hacia adentro. El tubo que se forma se convierte en el tracto gastrointestinal, y la abertura (¿puedes adivinar?) se convierte en el ano. Por lo tanto, el embrión ahora tiene una dirección de cola y dirección de cabeza distintas. La Fig. 6.23 representa la forma en que un erizo de mar gastrula. Los embriones humanos se ven diferentes porque es solo el disco embrionario el que se somete a gastrulación, no toda la blástula. Pero las 2 capas son iguales, las gastrulas humanas simplemente se ven más aplastadas. Ah, y los erizos de mar no forman ano primero. La razón por la que la gastrulación de erizos de mar parece más simple que la gastrulación humana es que las blastulas humanas reservan algunas células (el trofoblasto) para formar una placenta y amnios, mientras que los erizos de mar arrojan sus huevos al mar. Estamos sacrificando un poco de precisión aquí para enfocarnos en conceptos importantes compartidos entre especies. Si tienes curiosidad, ver películas de estos procesos en humanos no es posible, pero es para organismos como laevis xenpous (ranas).
A continuación, las celdas de la capa externa migran entre las dos capas. Esto le da al embrión 3 capas, que son las 3 capas germinales embrionarias (Fig. 6.23). La capa externa del ectodermo y la capa interna del endodermo permanecen epiteliales en apariencia, mientras que la capa media de células (mesodermo) sufre un cambio morfológico, y se convierte en mesénquima. Esta migración comienza en una única ubicación llamada nódulo de Hensen, que se desplaza hacia abajo por el embrión formando la racha primitiva detrás de él (Fig 6.24). El lado del embrión con el nódulo está destinado a convertirse en el lado dorsal, y la raya divide al embrión en una mitad izquierda y una mitad derecha. Ahora has sido testigo, aunque en forma de caricatura, cómo algo (todas las direcciones básicas del cuerpo) surgió de la nada. No dejes pasar desapercibido el significado de esto. A los científicos les cuesta explicar lo que había antes de que se formara el universo, o cómo la Tierra pasó de no tener vida a tener vida. Los embriólogos, por el contrario, explican cómo los embriones cambian de no tener forma a tener forma con gran detalle (hemos tenido el beneficio de observar y manipular directamente este proceso).
| 3 capas embrionarias | Destino celular |
|---|---|
| Ectodermo | Epitelio de piel y mucosa oral, tejido neural |
| Mesodermo | Tejido conectivo y muscular |
| Endodermo | Revestimiento epitelial de órganos huecos |
Cuadro 6.2: Las tres capas germinales se desarrollan durante la gastrulación

Transición epitelial a mesenquimal
El proceso por el cual las células ectodérmicas dejan de ser ectodermo y migran hacia el medio y forman mesodermo se denomina transición epitelial a mesenquimatosa (EMT). Las células epiteliales del ectodermo primero deben perder sus contactos célula a célula con células vecinas, lo que las hace menos epiteliales. Pierden su polaridad, se desdiferencian en una célula madre, luego se diferencian en células madre mesenquimales. Este proceso no solo es necesario para formar mesodermo durante la gastrulación, sino que ocurre durante la migración de la cresta neural, la cicatrización de heridas y la metástasis del cáncer. Las células en el mesodermo (el nombre para la capa media) forman mesénquima (un tipo de tejido), mientras que las otras dos capas permanecen epiteliales. Existe un proceso inverso, llamado transición mesenquimal a epitelial (MET), que también ocurre durante la cicatrización de heridas. La EMT permite que las células madre epiteliales de una región sana de la mucosa oral se conviertan en células madre mesenquimales, migren lejos de la región sana a una región dañada. El MET permite entonces que las células madre mesenquimales se conviertan de nuevo en células madre epiteliales, las cuales se dividen y se diferencian en nuevos queratinocitos. Debido a que esto imita lo que sucede durante la gastrulación, decimos “la cicatrización de heridas recapitula (afirma nuevamente) el desarrollo”.
Diferenciación de ectodermo

Neurulación
La gastrulación le dio al embrión su primera polaridad, así como el inicio del tracto GI. Uno de los siguientes sistemas de órganos en desarrollar es el sistema nervioso central. Los procesos básicos utilizados en la neurulación se reutilizan cuando se forman otros órganos huecos. Primero, el mesodermo subyacente conocido como notocorda envía morfógenos a las células ectodérmicas cercanas. Esto induce a la región cercana del ectodermo a diferenciarse en neuroectodermo, seguido de proliferación local. Debido a que estas células epiteliales no pueden propagarse fácilmente de lado a lado, invaginan. En última instancia, las células neuroectodérmicas invaginantes pierden sus contactos con el ectodermo y se fusionan con otras células neuro-ectodérmicas, creando una nueva estructura llamada tubo neural. Este tubo se desarrolla en el cerebro y la columna vertebral. Si te estás preguntando sobre la notocorda, su destino celular es sufrir mayoritariamente apoptosis, aunque algunos permanecen como el núcleo pulposis de los discos vertebrales. En pequeños cordados parecidos a peces sin columna vertebral llamados anfioxis, la notocorda persiste. Esta no será la última vez que veas a un embrión humano hacer una estructura que desaparece o cambia por completo.
El tubo neural comienza a plegarse desde la porción anterior del embrión y se cierra en una dirección anterior a posterior. La cremallera no es perfecta, el extremo anterior también va creciendo más ancho a medida que se pliega para formar el cerebro primitivo. Para todos los procesos de desarrollo que cubrimos y no cubrimos, el extremo anterior suele ser la región que se desarrolla primero, seguido de regiones más posteriores. La compresión del tubo neural requiere niveles adecuados de ácido fólico, por lo que es importante que la melanina de la madre haya protegido esta vitamina, de lo contrario puede resultar un cierre incompleto del tubo neural (espina bífida).

Algunas de las células del neuroectodermo no se fusionan. En cambio, estas células experimentan una transición epitelial a mesenquimatosa y migran lejos del tubo neural. Estas células son células de la cresta neural, están destinadas a convertirse en una serie de células y tejidos importantes en todo el cuerpo, incluyendo melanocitos, odontoblastos, pulpa dental, cementoblastos y el neuromesénquima de los arcos faríngeos. Estas células son algunas veces referidas como el cuarto tejido embrionario (además de las tres que surgen durante la gastrulación), lo que sugiere que algunas personas piensan que la cresta neural es importante. Incluso después de que las células de la cresta neural migran a tejidos nuevos y distantes, a menudo retienen signos visibles de su linaje neural, como las dendritas en los melanocitos o el proceso odontoblástico de los odontoblastos. Para facilitar la migración, las células de la cresta neural expresan una enzima llamada metaloproteinasa de matriz (MMP). Esta enzima digiere las proteínas que se encuentran en la matriz extracelular. A estas alturas esperamos recordar la frase cicatrización de heridas recapitula el desarrollo, las enzimas metaloproteinasas de matriz se reutilizan para curar cierto tipo de heridas dentales. Cuando las células de la cresta neural alcanzan los arcos faríngeos, secretan los morfógenos FGF y BMP, que se antagonizan entre sí, produciendo un patrón rayado a lo largo del ectodermo. Algunas regiones son inducidas a formar yemas dentales, las regiones entre ellas producen mucosa oral.
Diferenciación del mesodermo
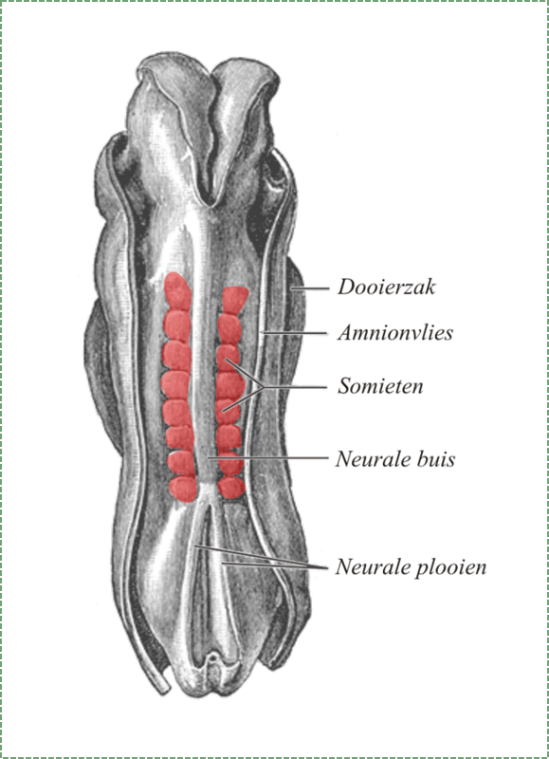
Formación de somitas
Durante el mismo tiempo que la neurulación, el mesodermo experimenta diferenciación. El mesodermo comienza como una capa amorfa de mesénquima. Luego, las regiones del mesénquima se pellizcan en estructuras esféricas repetitivas llamadas somitas, formando segmentos a lo largo del eje anterior a posterior del embrión. El destino de los somitas es convertirse en órganos sólidos, ya sea repitiendo unidades de tejido conectivo como las costillas y vértebras, o unidades repetitivas de tejido muscular como el recto abdominal y los músculos intercostales. La formación de cada somita implica una transición mesenquimal a epitelial, algunas células madre mesenquimales se diferencian en un epitelio que separa a un somita del siguiente.

Formación del corazón
En la tercera semana, el mesodermo también comienza a desarrollarse en sangre, corazón y sistema circulatorio. Las células madre mesenquimales se diferencian primero en islas sanguíneas. A continuación, comienza la angiogénesis. Dos grandes vasos sanguíneos se fusionan —con alguna ayuda de las células de la cresta neural— para formar el corazón primitivo, que comienza a latir.
Diferenciación del endodermo
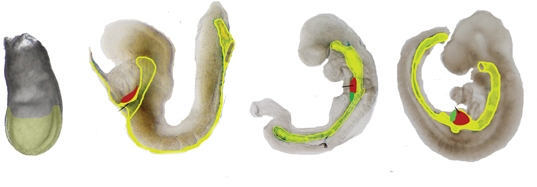
Formación de la faringe
El endodermo que migra interiormente durante la gastrulación se extiende hacia el extremo anterior del embrión. Esto forma el intestino anterior primitivo, que posteriormente se convierte en la faringe, la cavidad oral y la cavidad nasal. En este momento, sin embargo, no hay aberturas para la boca y las fosas nasales.

4ª semana
Ahora sólo estamos hasta la 4ta semana de desarrollo. ¡Han pasado muchas cosas! En este momento, el embrión continúa atravesando regiones locales de proliferación, modelado, fusión y los demás procesos cubiertos al inicio del capítulo. Esto conduce a la formación de estructuras rudimentarias (básicas). Cuando una estructura se vuelve visible por primera vez desde la superficie de un embrión, a menudo se le llama placode (un engrosamiento ectodérmico). Las células de un placode están destinadas a convertirse en un órgano, pero su forma general no es más que una protuberancia elevada o un pozo poco profundo. A la 4ta semana comienzan a desarrollarse los órganos relacionados con la cabeza enumerados en el Cuadro 6.3.
| Estructura | Destino |
|---|---|
| Placodes oculares | ojos primitivos |
| Placodes óticas | orejas primitivas |
| Placodes nasales | nariz primitiva |
| Arcos faríngeos | tantas cosas que obtienen su propio capítulo |
| Membrana orofaríngea | boca primitiva (aún no abierta) |
Cuadro 6.3: Estructuras primitivas formadas durante la 4ª semana de desarrollo
Aplicaciones clínicas del desarrollo temprano
Las alteraciones en estas primeras etapas de desarrollo suelen ser tan graves que el embrión no sobrevive. Sin embargo, existen condiciones clínicas relacionadas con estas etapas tempranas y con la salud bucal.
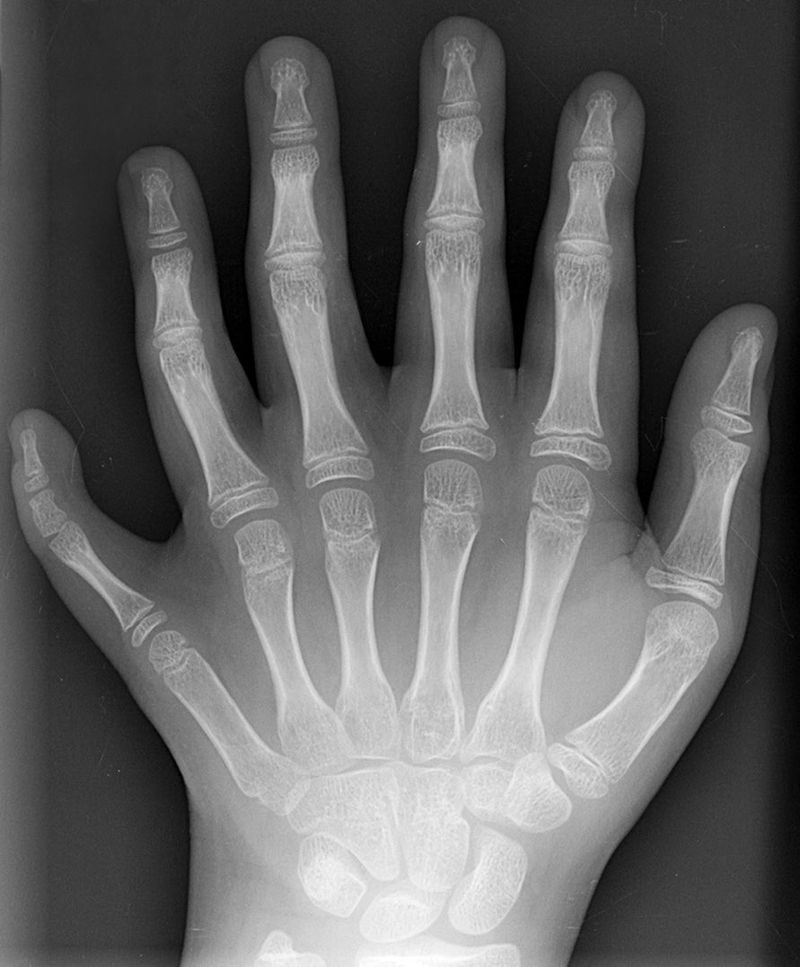
Mutaciones del gen homeobox
Las mutaciones en el gen homeobox pueden hacer que falten estructuras enteras o que se desarrollen en la ubicación incorrecta. Los primeros experimentos con moscas condujeron a observaciones de patas donde deberían ubicarse las antenas, o antenas donde deberían ubicarse las alas. Existen algunos trastornos genéticos relacionados con la homeobox en humanos. Los trastornos relacionados con la homeobox son raros porque las mutaciones suelen ser letales, pero cuando no lo son, las afecciones suelen ser graves. Una pequeña lista se encuentra en la Tabla 6.4
| Síndrome | Gen homeobox involucrado | Los síntomas incluyen |
|---|---|---|
| Axenfeld-Rieger Syndome | FOXC1, PITX2 | Principalmente relacionado con los ojos, a veces hipodoncia y microdoncia |
| Sindactilia autosómica dominante | HOXD13 | Sindactilia (dedos de manos o pies fusionados) |
| Síndrome de Guttmacher | HOXA13 | Polidactilia (dedos de manos o pies adicionales) |
| Displasia ectodérmica | HOXC13 | Ver abajo |
Cuadro 6.4: Algunos ejemplos de mutaciones homeobox en humanos.
Síndrome de Apert
El síndrome de Apert es causado por una mutación en el receptor del morfógeno FGF. Se categoriza por una amplia gama de síntomas, incluyendo deformaciones craneales y sindactilia. El FGF está involucrado en la formación del aparato de arco faríngeo (cubierto en el siguiente capítulo), lo que explica las anomalías craneofaciales. Relevante para este capítulo es la capacidad del FGF (como muchos factores de crecimiento) para inhibir la apoptosis. Tener una mutación que provoca que un receptor de FGF esté encendido todo el tiempo inhibe la apoptosis en las paletas de manos y pies. Se requieren regiones de apoptosis para producir dedos de manos y pies, de ahí que esta mutación conduzca a la sindactilia. Las perturbaciones parciales a las señales de FGF y BMP pueden provocar sindactilia parcial o cincha de los dedos de las manos o los pies.
 Figura 6.35: Figura 185: “Michael Berryman, actor” de Stefan Borggraefe está licenciado bajo CC BY 4.0
Figura 6.35: Figura 185: “Michael Berryman, actor” de Stefan Borggraefe está licenciado bajo CC BY 4.0Displasia ectodérmica (parte 1 de 2)
Los problemas con la inducción de las células de la cresta neural durante la neurulación conducen a alteraciones en la formación de dientes, folículos pilosos, glándulas salivales y otras estructuras. Lo que estos tienen en común es que todas son estructuras especializadas del ectodermo, inducidas por las células de la cresta neural para diferenciarse. En un embrión sano, las células ectodérmicas reciben morfógenos que activan o inactivan los factores de transcripción correctos para desencadenar la diferenciación en células de la cresta neural. Las células de la cresta neural migran a regiones distantes del cuerpo, determinan su ubicación interactuando con morfógenos en la sustancia del suelo y liberan otros morfógenos para inducir regiones de ectodermo a diferenciarse en glándulas sudoríparas, glándulas salivales, yemas dentales o folículos pilosos. Las mutaciones en cualquiera de los genes para estos morfógenos, sus proteínas receptoras o factores de transcripción descendentes pueden conducir a una afección llamada Displasia Ectodérmica. Este grupo de síndromes es raro, con sólo 7.000 casos a nivel mundial, pero hay al menos 40 genes diferentes que han sido implicados en estos síndromes. Compare eso con la enfermedad de células falciformes, que actualmente afecta a más de 100,000 estadounidenses (predominantemente afroamericanos, hispanoamericanos, grecoamericanos, turcoamericanos e italianos americanos), todo debido a mutaciones en un solo gen, la Hemoglobina-Beta. El punto de esta comparación es resaltar cuándo se expresan estos genes. La inducción de células madre ectodérmicas a proliferar y diferenciarse en diferentes apéndices es compleja y ocurre durante la embriogénesis. La mutación en los morfógenos, los receptores de morfógenos, los segundos mensajeros o factores de transcripción y los genes aguas abajo que se activan para inducir la diferenciación son todas dianas posibles que causan displasia ectodérmica. Las mutaciones en estos genes en una célula de la cresta neural conducen a una alteración en cualquiera de las células posteriores inducidas por este nuevo tipo celular, similar a la forma en que abordar al portador de pelota en socer (fútbol) despliega la jugabilidad de cualquiera de sus posibles objetivos pasantes. En comparación, solo los glóbulos rojos expresan hemoglobina-beta, y son células diferenciadas terminalmente, no se convierten en ningún otro tipo celular. Obviamente, los glóbulos rojos son un tipo celular importante, uno que se puede encontrar temprano en la embriogénesis, pero los síntomas de la Enfermedad de Células falciformes presentan menos espectro que los tipos de enfermedades que hemos estado discutiendo, como la Displasia Ectodérmica.
 Figura 6.36: Gaten John Matarazzo III, actor y activista del CCD. Crédito de imagen: Gaten John Matarazzo III, de Gage Skidmore, tiene licencia CC BY SA 3.0
Figura 6.36: Gaten John Matarazzo III, actor y activista del CCD. Crédito de imagen: Gaten John Matarazzo III, de Gage Skidmore, tiene licencia CC BY SA 3.0
Disostosis cleido-craneal
La distostosis cleido-craneal (CCD) es un trastorno congénito causado por una mutación a un factor de transcripción requerido para la diferenciación de hueso y dientes. Se requiere desencadenar células osteo-condroprogenitoras para que salgan del ciclo celular y se diferencien en osteoblastos. También se reutiliza para inducir la diferenciación de odontoblastos. Además, una vez formados los dientes, este factor de transcripción se reutiliza para activar la expresión de una enzima metaloproteinasa Matrix, que es necesaria para la remodelación de las cavidades alveolares. Sin esta enzima, se produce la retención de dientes caducifolios. Los implantes dentales o dentaduras postizas (como las que recibió el actor y el filántropo CCD-Gaten John Matarazzo III en la Fig. 6.36) son el tratamiento preferido. Además, una persona con CCD puede tener clavículas pequeñas y cambios en la forma del cráneo, huesos que se forman por osificación intramembranosa. Esto ilustra dos conceptos principales en el desarrollo. Primero, muchas estructuras forman una sola vía, pero son remodeladas posteriormente para cumplir una función diferente (los dientes se forman doblando hacia adentro, luego se mueven hacia afuera). En segundo lugar, muchos patrones diferentes en embriología son re-demandados (recapitulados), como la extracción de tejido durante la migración de la cresta neural y la extracción de tejido durante la erupción dental.
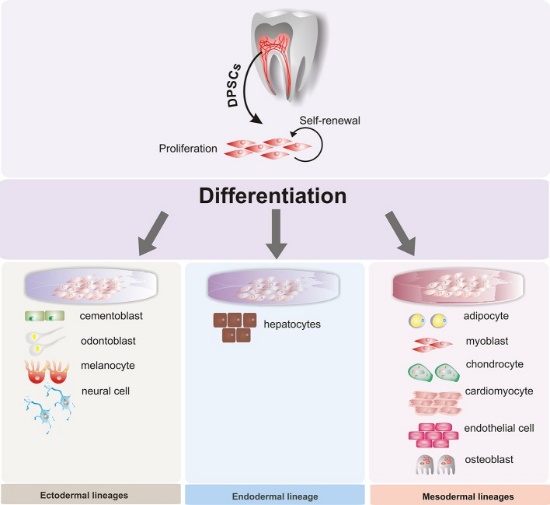
Terapias con células madre
Cuando las células se diferencian terminalmente, inactivan permanentemente genes innecesarios por metilación y almacenamiento alrededor de las histonas. Los investigadores están aprendiendo formas de revertir este proceso y guiar a las células diferenciadas para que vuelvan a un estado de células madre. Esto plantea la posibilidad de promover la regeneración de tejidos que de otra manera no se regeneran, eliminando la necesidad de injertos o trasplantes de tejidos. Debido a que algunos tejidos dentales no se regeneran bien, existe la posibilidad de que estas tecnologías se apliquen a la cavidad oral, como cultivar implantes dentales biológicos en lugar de usar metales y cerámicas. Sin embargo, se ha puesto más interés en adquirir células madre mesenquimales de tejidos dentales. Por ejemplo, las células madre aisladas de terceros molares maxilares se han utilizado en ensayos clínicos para mejorar la cicatrización y reducir la necesidad de trasplantar tejido en cirugía maxilofacial. Debido a su potencial para diferenciarse en una amplia gama de células, se ha puesto gran interés en recolectar células madre dentales para tratar enfermedades no relacionadas con la cavidad oral. Con el morfógeno correcto y abundancia de células madre dentales, es posible revertir el daño causado por enfermedades neurodegenerativas como la Enfermedad de Alzheimer (EA) y la Enfermedad de Parkinson (EP), trauma espinal, infarto de miocardio (ataque cardíaco) y Distrofia Muscular (MD). El linaje de las células que producen dentina, pulpa, cemento y periodonto ayudará a explicar el vínculo con desobedientes neurodegenerativos. ¿Qué más harás con los terceros molares extraídos? Desafortunadamente, en Estados Unidos no existe una definición legal para lo que constituye una terapia segura y efectiva con células madre. Aunque se pongan buenas reglas, el turismo médico no se atiene. Además de una investigación honesta y buena en terapias con células madre (el Instituto de Células Madre y Medicina Regenerativa de la Universidad de Washington es un buen ejemplo), hay una gran cantidad de clínicas de células madre dudosas a abajo-curanderas que operan en el mercado gris, utilizando arriesgados, no probados y haciendo afirmaciones absurdas para tomar la mayor cantidad de dinero posible de las personas vulnerables. Entonces, ¿quién es quién? Las terapias que usan o potencian las células madre de un paciente tienen la mejor oportunidad posible de proporcionar resultados positivos con un riesgo mínimo de efectos secundarios, especialmente si esa terapia es dirigida, como agregar un morfógeno específico para impulsar un resultado específico. Por otro lado, arrojar células madre exógenas ante un problema —células de otras personas, otros animales, o de fuentes no animales— conlleva el mayor riesgo de efectos adversos graves con un mínimo potencial de beneficio. Tenga en cuenta que 19 de cada 20 (más o menos) fármacos potenciales que se muestran científicamente que funcionan en modelos animales fallan en los ensayos en humanos. Eso significa que hay evidencia científica en la publicación de tratamientos que sabemos que no funcionan, pero otras personas pueden seleccionar la información positiva y usarla para vender un producto.
Capítulo 5 * Capítulo 7