
15.4A: Selección Clonal y Memoria Inmunológica
- Page ID
- 56772

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
La capacidad del sistema inmune para responder a un antígeno existe antes de que se encuentre con ese antígeno. El sistema inmunológico se basa en la formación previa de una población increíblemente diversa de:
- Células B (linfocitos B) cada una con su superficie cubierta con miles de copias idénticas de un receptor para antígeno (el receptor de células B para antígeno = BCR)
- Células T (linfocitos T) cada una con su superficie cubierta con miles de copias idénticas de un receptor de células T para antígeno (TCR)

La figura anterior ilustra la activación de una célula B en un conjunto de células B cuyo BCR es específico para un epítopo (pequeñas esferas oscuras) en el antígeno. Este fenómeno se denomina selección clonal porque es antígeno el que selecciona linfocitos particulares para la expansión clonal. La selección clonal conduce a la eventual producción de:
- Un conjunto de células plasmáticas secretoras de anticuerpos. Las células plasmáticas son células B que se han labrado (por ejemplo, formando un gran retículo endoplásmico) para la síntesis masiva y secreción de un anticuerpo. El anticuerpo es la versión secretada del BCR. (Para mayor claridad, cada BCR se muestra con un único sitio de unión para el epítopo; en realidad, los BCR son IgM y cada uno tiene 10 sitios de unión idénticos.
- Un pool de celdas de “memoria”. Se trata de linfocitos B con receptores de la misma especificidad que los de la célula B activada original.
Cómo las células B y las células T se encuentran con los antígenos
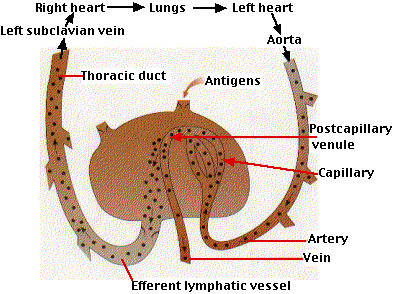
Fig.15.4.1.2 Vénulas postcapilares
¿Cuál es la probabilidad de que esos pocos linfocitos capaces de unirse a un epítopo en particular se encuentren realmente con el antígeno portador de ese epítopo?
Sorprendentemente, es bastante alta porque tanto las células B como las células T migran dentro y fuera de los ganglios linfáticos y el bazo. Los ganglios linfáticos sirven como filtros linfáticos, atrapando la materia extraña que obtiene acceso a los tejidos. Se ha demostrado que hasta el 99% de las bacterias que entran en un nódulo son removidas por éste. De igual manera, el bazo atrapa antígenos que obtienen acceso a la sangre. Incluso si un invasor no logra ingresar a los vasos linfáticos o sanguíneos, sus antígenos aún pueden llegar a los ganglios linfáticos y al bazo transportados allí por células dendríticas que
- engullir el antígeno en los tejidos
- migrar en los vasos linfáticos a los ganglios linfáticos cercanos o al bazo
- procesar el antígeno y “presentarlo” a las células T y también a las células B

El rechazo del injerto es una forma de inmunidad mediada por células. Si un trozo de piel de un ratón de una cepa (B) se injerta en el flanco de un ratón de una segunda cepa (A), el injerto funciona bien al principio. Los vasos sanguíneos del huésped crecen en él, y funciona normalmente. Después de unos 10-14 días, sin embargo, el suministro de sangre al injerto se descompone y el injerto degenera. Finalmente se desprende como una vieja costra. Esto se llama rechazo del primer set. Que el rechazo del injerto es una respuesta inmune se demuestra ahora injertando al ratón con dos trozos de piel una repetición de piel de la cepa B y piel de otra cepa (C)
Los resultados son bastante diferentes.
- Es posible que la piel B ni siquiera sobreviva el tiempo suficiente para adquirir un suministro de sangre. Se rechaza en un periodo mucho más corto (menos de una semana). Este fenómeno de “segundo conjunto” es el equivalente de células T de la respuesta secundaria de “memoria” de las células B. Su especificidad y memoria se demuestra por el hecho de que
- El injerto de piel C es rechazado en el periodo normal de “primer conjunto”.