
15.4C: Células B y Células T
- Page ID
- 56744

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Los linfocitos son uno de los cinco tipos de glóbulos blancos (leucocitos) que circulan en la sangre. Aunque todos los linfocitos maduros se parecen bastante, son extraordinariamente diversos en sus funciones. Los linfocitos más abundantes son los linfocitos B (células B) y los linfocitos T (células T). Las células B se producen en la médula b uno. Los precursores de las células T también se producen en la médula ósea pero dejan la médula ósea y maduran en el himo t (lo que explica su designación). Cada célula B y célula T es específica para un antígeno particular. Lo que esto significa es que cada uno es capaz de unirse a una estructura molecular particular. La especificidad de unión reside en un receptor para antígeno: el receptor de células B (BCR) para antígeno y el receptor de células T (TCR) respectivamente.
Tanto los BCR como los TCR comparten estas propiedades:
- Son proteínas integrales de membrana.
- Están presentes en miles de copias idénticas expuestas en la superficie celular.
- Se hacen antes de que la célula se encuentre alguna vez con un antígeno.
- Están codificados por genes ensamblados por la recombinación de segmentos de ADN.
- Tienen un sitio de unión único.
- Este sitio se une a una porción del antígeno llamada determinante antigénico o epítopo.
- La unión, como la que existe entre una enzima y su sustrato, depende de la complementariedad de la superficie del receptor y la superficie del epítopo.
- La unión se produce por fuerzas no covalentes (nuevamente, como una enzima que se une a su sustrato).
- La unión exitosa del receptor de antígeno al epítopo, si se acompaña de señales adicionales, da como resultado:
- estimulación de la célula para dejar G 0 e ingresar al ciclo celular.
- La mitosis repetida conduce al desarrollo de un clon de células que portan el mismo receptor de antígeno; es decir, un clon de células de idéntica especificidad.
Los BCR y TCR difieren en su estructura, los genes que los codifican y el tipo de epítopo al que se unen.
Células B

Fig.15.4.3.1 Células B y linfocinas
- Los BCR se unen a antígenos intactos (como el toxoide diftérico, la proteína introducida en su cuerpo en la vacuna DTP). Estos pueden ser
- moléculas solubles presentes en el fluido extracelular
- moléculas intactas que la célula B extrae de la superficie de células presentadoras de antígeno como macrófagos y células dendríticas
- Las moléculas de antígeno unidas se engullan en la célula B mediante endocitosis mediada por receptores.
- El antígeno se digiere en fragmentos que luego se muestran en la superficie celular enclavada dentro de una molécula de histocompatibilidad de clase II.
- Los linfocitos T auxiliares específicos para esta estructura (es decir, con TCR complementarios) se unen a la célula B y secretan linfocinas que:
- estimular la célula B para que entre en el ciclo celular y se desarrolle, por mitosis repetida, en un clon de células con BCR idénticos
- cambiar de sintetizar sus BCR como proteínas de membrana integrales a una versión soluble
- diferenciarse en células plasmáticas que secretan estos BCR solubles, que ahora llamamos anticuerpos
Células T
La superficie de cada célula T también muestra miles de receptores idénticos de células T (TCR). Hay dos tipos de células T que difieren en su TCR:
- linfocitos T alfa/beta (αβ). Su TCR es un heterodímero de una cadena alfa con una cadena beta. Cada cadena tiene una región variable (V) y una región constante (C). Las regiones V contienen cada una 3 regiones hipervariables que conforman el sitio de unión al antígeno.
- células T gamma/delta (γδ). Su TCR también es un heterodímero de una cadena gamma emparejada con una cadena delta.
La discusión que sigue ahora se refiere a las células T alfa/beta. Las células T gamma/delta, que son menos entendidas, se discuten al final.
El TCR (de las células T alfa/beta) se une a un complejo bimolecular que se muestra en la superficie de alguna otra célula llamada célula presentadora de antígeno (APC). Este complejo consiste en un fragmento de un antígeno que se encuentra dentro del surco de una molécula de histocompatibilidad. El complejo ha sido comparado con un “hot dog in a bun”.

La mayoría de las células T del cuerpo pertenecen a uno de los dos subconjuntos. Estas se distinguen por la presencia en su superficie de una u otra de dos glicoproteínas designadas:
- CD4
- CD8
Cuál de estas moléculas está presente determina a qué tipos de células se puede unir la célula T.
- Las células T CD8+ se unen a epítopos que forman parte de moléculas de histocompatibilidad de clase I. Casi todas las células del cuerpo expresan moléculas de clase I.
- Las células T CD4+ se unen a epítopos que forman parte de moléculas de histocompatibilidad de clase II. Solo las células presentadoras de antígeno especializadas expresan moléculas de clase II. Estas incluyen células dendríticas, células fagocíticas como macrófagos y células B.
Células T CD8 +
Los linfocitos T CD8+ mejor entendidos son los linfocitos T citotóxicos (CTL). Secretan moléculas que destruyen la célula a la que se han unido.
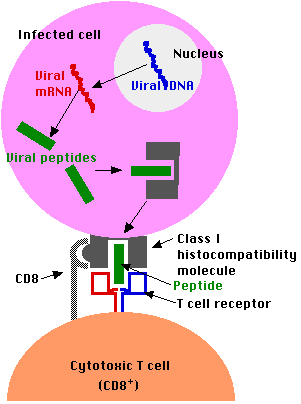
Esta es una función muy útil si la célula diana está infectada con un virus porque la célula generalmente se destruye antes de que pueda liberar una cosecha fresca de virus capaces de infectar a otras células. Un ejemplo mostrará la belleza y eficiencia biológica de este mecanismo.
Cada vez que contrae una infección por virus, digamos influenza (gripe), el virus invade ciertas células de su cuerpo (en este caso células de las vías respiratorias). Una vez dentro, el virus subvierte el metabolismo de la célula para producir más virus. Esto implica sintetizar una serie de diferentes macromoléculas codificadas por el genoma viral. A su debido tiempo, estos se ensamblan en un cultivo fresco de partículas de virus que salen de la célula (a menudo matándola en el proceso) y se propagan a nuevas células diana. Excepto mientras están en tránsito de sus antiguas casas a sus nuevas, los virus funcionan dentro de tus células a salvo de cualquier anticuerpo que pueda estar presente en la sangre, la linfa y las secreciones. Pero al principio del proceso, las células infectadas muestran fragmentos de las proteínas virales en sus moléculas de clase I de superficie. Los CTL específicos para ese antígeno podrán unirse a la célula infectada y a menudo podrán destruirla antes de que pueda liberar una cosecha fresca de virus.
En general, el papel de las células T CD8 + es monitorear todas las células del cuerpo, listas para destruir cualquiera que exprese fragmentos de antígeno extraños en sus moléculas de clase I.
Células T CD4 +
Las células T CD4+ se unen a un epítopo que consiste en un fragmento de antígeno que se encuentra en el surco de una molécula de histocompatibilidad de clase II. Las células T CD4+ son esenciales tanto para las ramas del sistema inmune mediadas por células como para las mediadas por anticuerpos:
- Inmunidad mediada por células: Estas células CD4+ se unen al antígeno presentado por las células presentadoras de antígeno (CPA) como macrófagos fagocíticos y células dendríticas. Las células T luego liberan linfocinas que atraen a otras células a la zona. El resultado es la inflamación: la acumulación de células y moléculas que intentan tapizarse y destruir el material antigénico (un absceso es un ejemplo, el sarpullido después de la exposición a la hiedra venenosa es otro).
- Inmunidad mediada por anticuerpos: Estas células CD4+, llamadas células T colaboradoras, se unen al antígeno presentado por las células B. El resultado es el desarrollo de clones de células plasmáticas que secretan anticuerpos contra el material antigénico.
El SIDA proporciona una vívida ilustración de la importancia de las células T CD4+ en la inmunidad. El virus de la inmunodeficiencia humana (VIH) se une a moléculas CD4 y así es capaz de invadir e infectar células T CD4 +. A medida que avanza la enfermedad, el número de células T CD4 + disminuye por debajo de su nivel normal de aproximadamente 1000 por microlitro (µl). Una explicación parcial de esto puede ser el incesante esfuerzo de las células T CD8+ del paciente para destruir las células CD4+ infectadas. Sin embargo, resulta que solo una pequeña fracción de los pacientes con células T CD4+ se infectan en un momento dado. Cómo las células CD4+ no infectadas pueden ser inducidas a suicidarse se discute en la página sobre apoptosis.
Cuando el número de células T CD4+ cae por debajo de 400 por microlitro, la capacidad del paciente para montar una respuesta inmune disminuye peligrosamente. No sólo el paciente se vuelve hipersusceptible a patógenos que nos dan pena a todos sino también a microorganismos, especialmente virus y hongos, que normalmente habitan nuestros tejidos sin dañarnos. Estas infecciones oportunistas pueden ser fatales.
Construyendo el repertorio de células T
Las células T tienen receptores (TCR) que se unen a fragmentos de antígeno ubicados en moléculas del MHC. Pero todas las células expresan moléculas MHC de clase I que contienen fragmentos derivados de proteínas propias. Muchas células expresan moléculas MHC de clase II que también contienen autopéptidos. Esto presenta un riesgo para el animal de las células T reconociendo estos complejos de autopéptido/autoMHC y montando un ataque autoinmune contra ellos. Afortunadamente, esto suele evitarse mediante un proceso de selección que se desarrolla en el timo (donde se desarrollan todas las células T).
El proceso funciona así:
- Los precursores de las células T como todas las células sanguíneas se forman en la médula ósea.
- Estas células luego migran a la corteza del timo. En este momento no tienen ni un TCR completo ni CD4 o CD8 (así se denominan celdas “doble-negativas” o DN).
- En la corteza del timo comienzan a formar un TCR y sintetizan tanto CD4 como CD8 (por lo que ahora son células “doblemente positivas” o DP). Las células corticales del timo expresan una amplia variedad de moléculas pequeñas, generalmente un péptido de 6—8 aminoácidos derivado de proteínas corporales; es decir, proteínas “propias” como proteínas dentro del citosol y proteínas séricas, proteínas que circulan en la sangre y linfa enclavada en una molécula de histocompatibilidad ( codificado por el MHC).
- La mayoría de las células (~ 97%) producirán un TCR que no se une a ninguna de las moléculas péptido-MHC presentes en la superficie de las células corticales. A menos que puedan volver a intentarlo con un nuevo TCR, estas células mueren por “abandono” (por apoptosis, en realidad).
- Aquellas células restantes cuyo TCR se ha unido a un antígeno peptídico presentado en la molécula MHC clase II dejan de expresar CD8 y se convierten en células T CD4+. Son estas células las que pasarán a convertirse
- Células Th1 en respuestas inmunes mediadas por células
- Células Th1 colaboradoras para linfocitos T citotóxicos (CTL)
- Células Th2 colaboradoras para células B
- Aquellas células restantes cuyo TCR se ha unido a un antígeno peptídico presentado en la molécula MHC clase I dejan de expresar CD4 y se convierten en células T CD8+.
- Se dice que ambos conjuntos de células han sido sometidos a selección positiva.
- Después de la selección positiva, estas células migran a la médula del timo.
- Allí se destruyen aquellas células cuyo TCR se une muy fuertemente a complejos de auto-péptido y auto-MHC (nuevamente por apoptosis).
- Este proceso de selección negativa es importante ya que elimina las células que de otro modo podrían montar un ataque autoinmune. Es una de las formas en que se logra la tolerancia a los autoantígenos. [Enlace a la discusión sobre la tolerancia de células T.]
- Las células cuyos TCR se unen al antígeno con una afinidad por debajo del umbral que desencadena la apoptosis son libres de abandonar el timo y migrar por todo el sistema inmunitario (ganglios linfáticos, bazo, etc.)
- Es esta población de la que dependemos para montar respuestas inmunitarias contra antígenos extraños. Un TCR que se une a autopéptido/autoMHC con baja afinidad puede unirse a un péptido extraño en el MHC propio con alta afinidad.
Células T Gamma/Delta (γδ)
Las células T gamma/delta difieren de sus primos alfa/beta de varias maneras:
- Su TCR está codificado por diferentes segmentos génicos.
- Su TCR se une a
- antígenos que pueden ser proteínas intactas (tal como lo hacen los anticuerpos) así como una variedad de otros tipos de moléculas orgánicas (a menudo contienen átomos de fósforo).
- antígenos que no se “presentan” dentro de moléculas de histocompatibilidad de clase I o clase II;
- antígenos que no son presentados por células presentadoras de antígeno (APC) “profesionales” como las células dendríticas.
- La mayoría de estas células T no tienen ni CD8 ni CD4 en su superficie. Esto tiene sentido porque no tienen necesidad de reconocer moléculas de histocompatibilidad de clase I y clase II.
- Las células T gamma/Delta, como las células T alfa/beta, se desarrollan en el timo. Sin embargo, migran de allí a los tejidos corporales, especialmente a los epitelios (por ejemplo, intestino, piel, revestimiento de la vagina), y no recirculan entre la sangre y los ganglios linfáticos (representan no más del 5% de las células T en la sangre y son aún más raras en los ganglios linfáticos). Se encuentran con antígenos en la superficie de las células epiteliales que las rodean en lugar de depender de las APC que se encuentran en los ganglios linfáticos.
¿Cuál es la función de las células T γδ?
Eso sigue siendo una especie de misterio. Situados como se encuentran en las interfaces entre el mundo externo e interno, pueden representar una primera línea de defensa contra patógenos invasores. Su respuesta parece ser más rápida que la de las células T αβ. Curiosamente, muchos de los antígenos a los que responden las células T γδ se encuentran no solo en ciertos tipos de invasores (por ejemplo, Mycobacterium tuberculosis, el agente de la tuberculosis) sino también en células hospedadoras que están bajo ataque por patógenos.
Los ratones knockout que no pueden producir linfocitos T γδ son más lentos para curar lesiones en su piel. También son mucho más susceptibles a los cánceres de piel que los ratones normales. Quizás la vigilancia inmune es una de las funciones de las células T γδ.