
5.3.2: Algas pardas y diatomeas
- Page ID
- 58335

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Usar historia de vida, morfología y componentes celulares para identificar algas pardas.
- Identificar los componentes de un talo de algas marinas.
- Identificar estructuras y eventos en el ciclo de vida de Fucus y conocer su ploidía.
- Identificar estructuras y eventos en el ciclo de vida de Laminaria y conocer su ploidía.
- Usar historia de vida, morfología y componentes celulares para identificar diatomeas.
- Clasificar las diatomeas según la simetría y la ecología.
- Describir la reproducción sexual y asexual en diatomeas.
Las algas pardas (clase Phaeophyceae) y las diatomeas (clase Bacillariophyceae) pertenecen al filo Ochrophyta. Son el resultado de una endosimbiosis secundaria entre un heteroconte y un eucariota fotosintético. Los heterocontes, como el Oomycota, están unidos por la presencia de un flagelo texturizado o “peludo” y un flagelo adicional que carece de proyecciones similares a pelos (Figura\(\PageIndex{1}\)). Los miembros de este subgrupo varían en tamaño desde diatomeas unicelulares hasta las algas masivas y multicelulares.
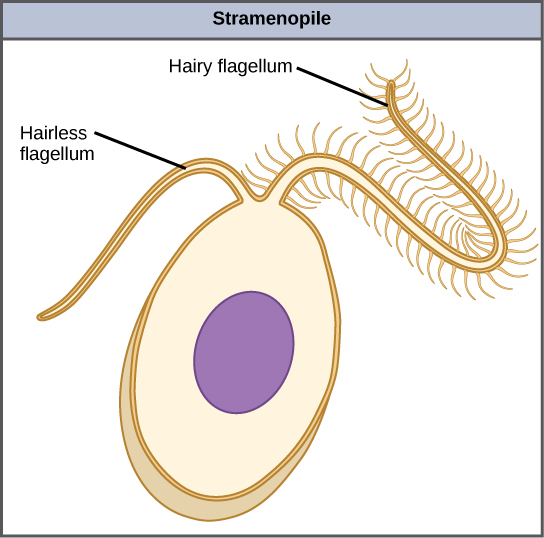
Las algas pardas, diatomeas y oomicetos pertenecen a un solo clado, los estramenopiles (también conocidos como heterocontes). Los estramenopiles fotosintéticos comparten las siguientes características:
- Cloroplastos de 4 membranas
- un pigmento amarillo-marrón (que les da su color). Es un carotenoide llamado fucoxantina.
- clorofilas a y c
- la mayoría tiene un ciclo de vida diplontico (como encontrarás a continuación, Laminaria tiene un ciclo de vida haplodiplontico)
Feofíceas
Este grupo se llama comúnmente algas pardas e incluye algas rocosas y algas marinas. ¡Las algas marinas son algunos de los organismos de más rápido crecimiento en el planeta! Las algas pardas son principalmente marinas y a menudo se encuentran en la zona intermareal. Los miembros de este filo se utilizan como alimento en algunas zonas costeras del mundo y se cosechan en los Estados Unidos para fertilizantes y como fuente de yodo.
Las algas pardas son pardas debido a las grandes cantidades de carotenoides que producen, principalmente uno llamado fucoxantina. Estos organismos son exclusivamente multicelulares y tienen células filamentosas, multinucleadas (muy parecidas a los oomicetos). Pueden llegar a ser tan grandes que requieren células conductoras especiales para transportar fotosintatos desde sus cuchillas hasta el resto de sus tejidos. Estas células conductoras se llaman hifas de trompeta y tienen placas de tamiz y se asemejan a tubos de tamiz que se encuentran en plantas con flores Las algas pardas tienen paredes celulares de celulosa y almacenan carbohidratos en forma de laminarina. El alginato polimérico también se puede encontrar en las paredes celulares de algas pardas y se usa comercialmente para una variedad de propósitos, incluyendo los moldes de alta fidelidad utilizados en odontología.
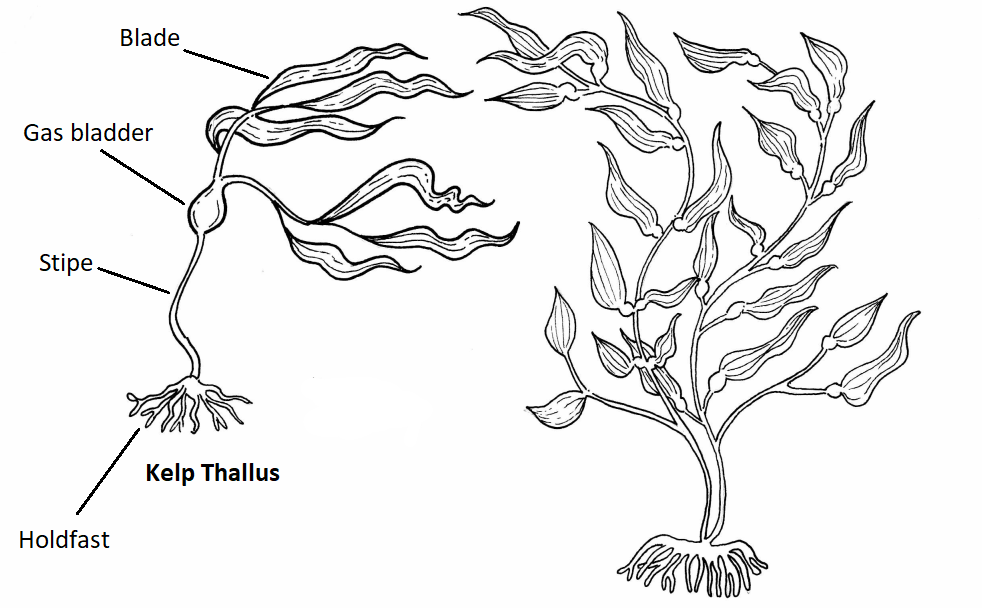
Kelp
Al igual que Saprolegnia, el cuerpo de una alga se denomina talo porque no se diferencia en tejidos especializados. La morfología general de una alga marrón incluye un sostén, estipe, vejiga (s) de gas y hoja (s) (Figuras\(\PageIndex{2-4}\)).


Fucus
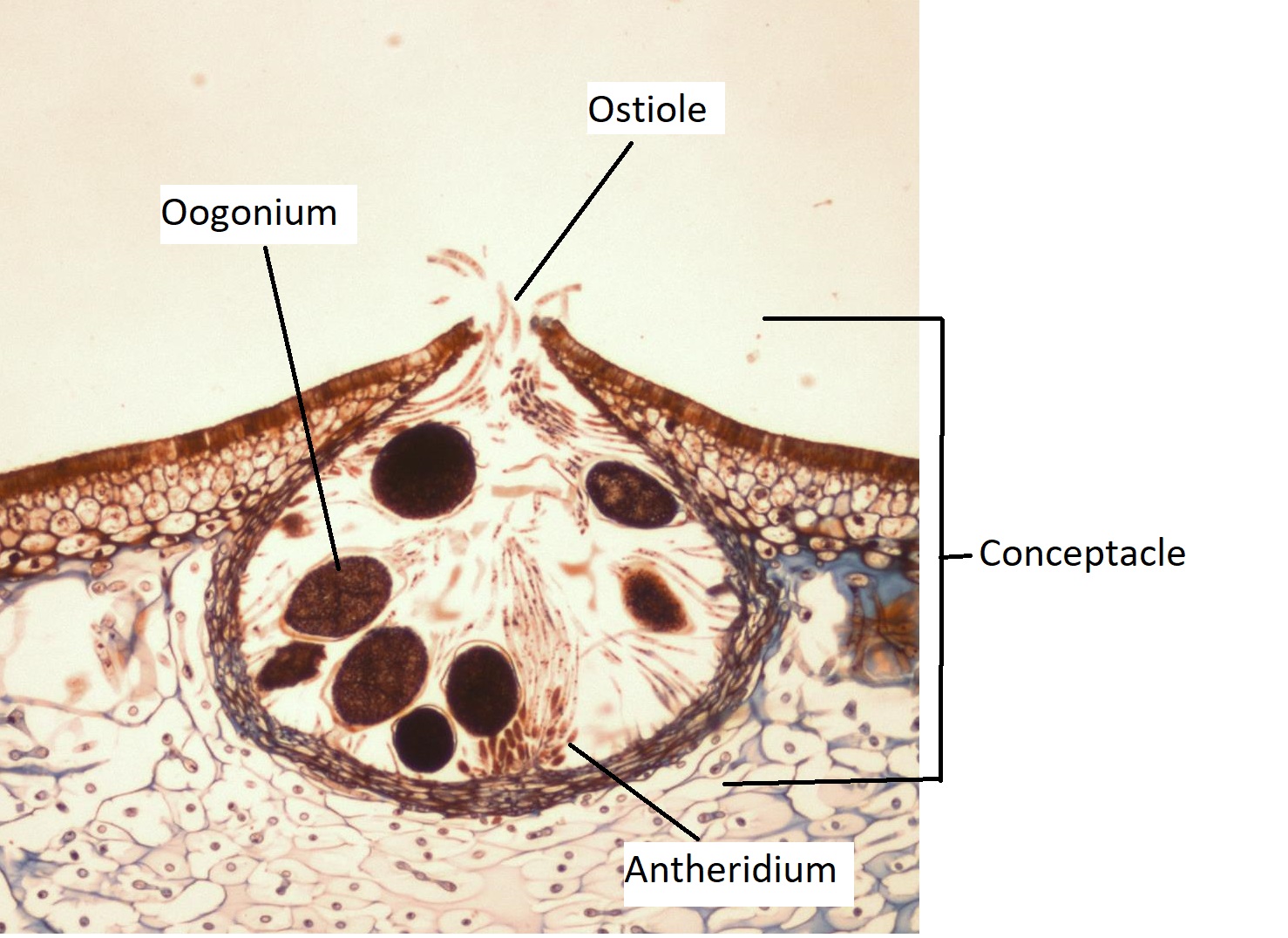
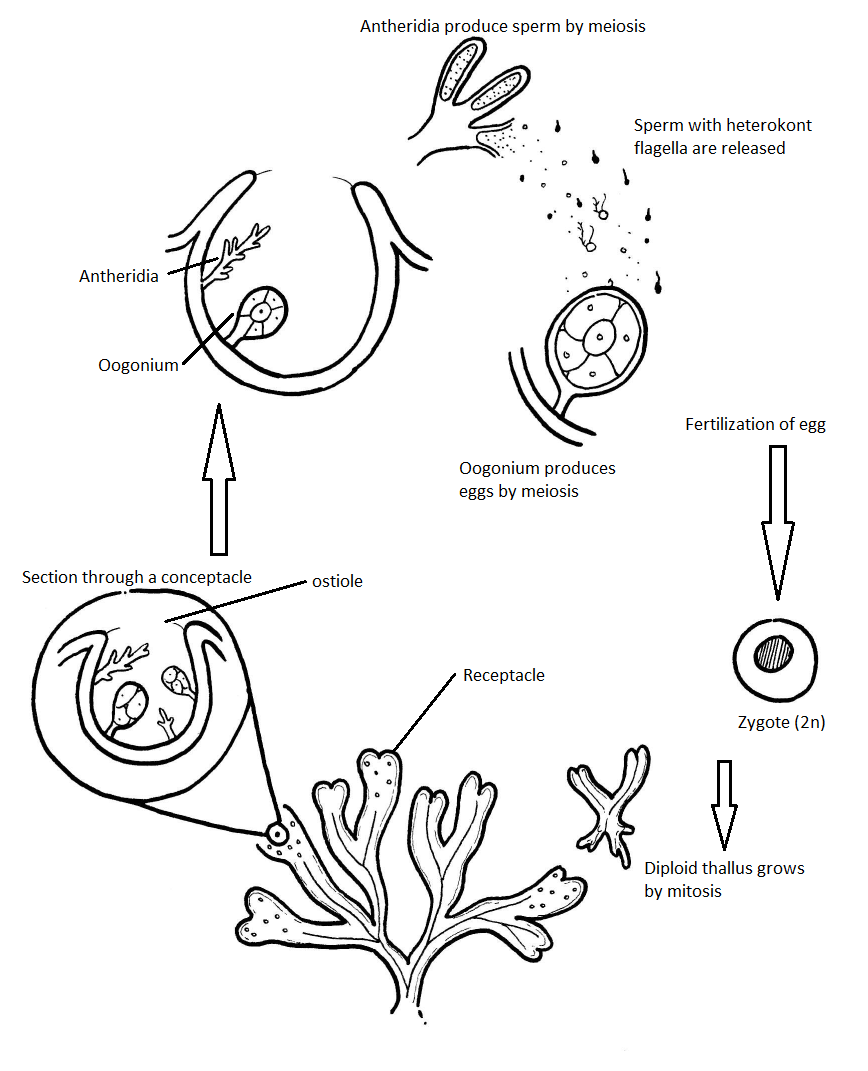
Un organismo modelo para el ciclo de vida de Phaeophyta es Fucus (rockweed), que, al igual que su relativa Saprolegnia, tiene un ciclo de vida diplónico. El Fucus thallus tiene ramificación dicotómica (bifurcada en dos ramas iguales) y puntas reproductivas hinchadas en forma de corazón de las ramas. Estas puntas de rama hinchadas se denominan receptáculos (Figura\(\PageIndex{5}\)).


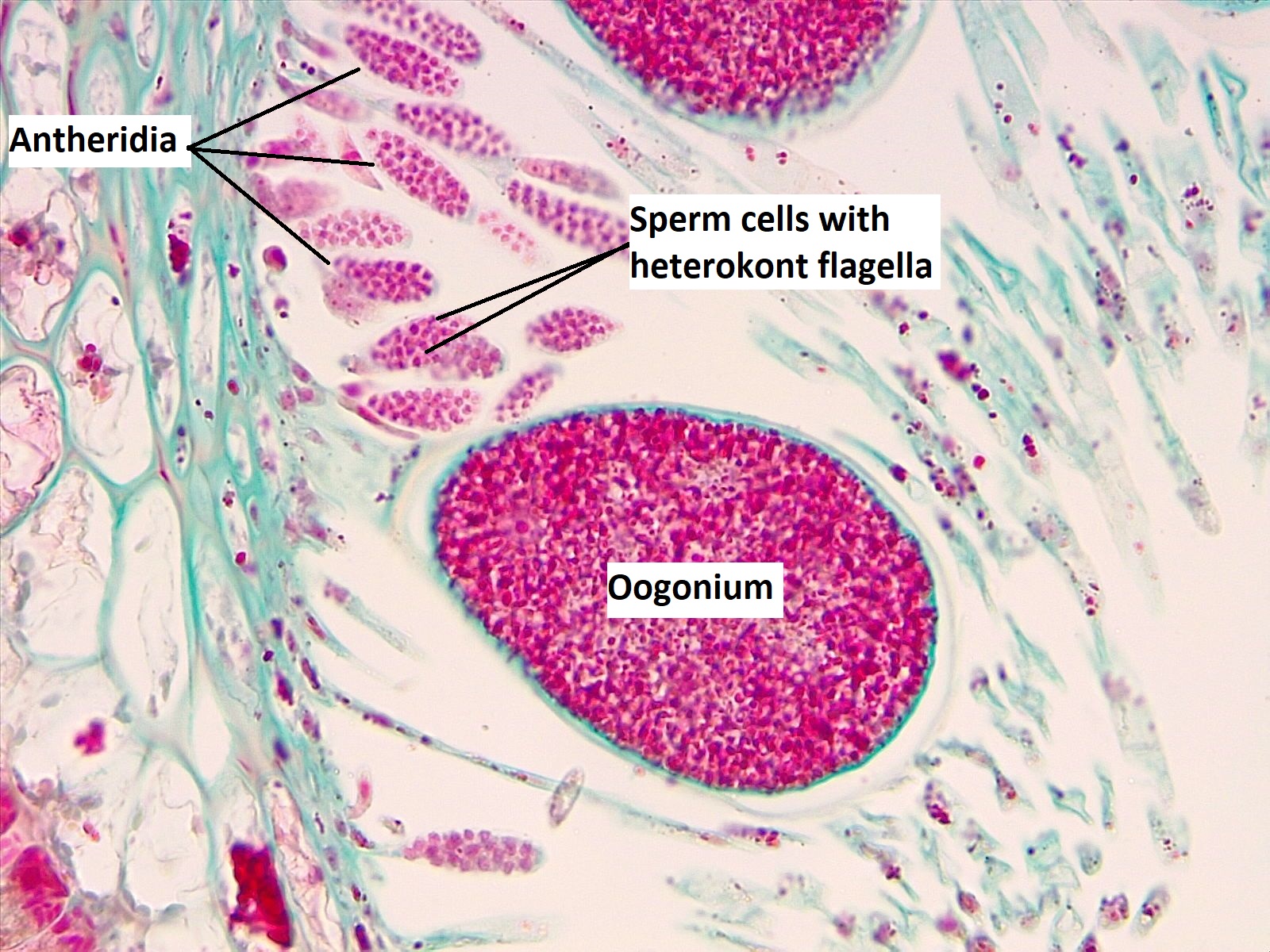
Los receptáculos están cubiertos de pequeñas protuberancias, cada una con un poro en el centro de la protuberancia llamado ostiole. Los baches son conceptacles, cámaras que albergan la gametangios (Figura\(\PageIndex{6}\)). La feofita produce oognia, gametangios globosos que se someten a meiosis para producir óvulos y anteridios, gametangios ramificados que se someten a meiosis para producir espermatozoides (Figura\(\PageIndex{7}\)).
Ciclo de vida de Fucus
El fucus tiene un ciclo de vida diplónico (Figura\(\PageIndex{8}\)) donde se producen gametos haploides a partir de un talo diploide. Estos gametos haploides no crecen, sino que se fusionan para formar un cigoto. Ver Figura\(\PageIndex{9}\) para un ejemplo de alternancia de generaciones en la Phaeophyta.
Ciclo de Vida de Laminaria
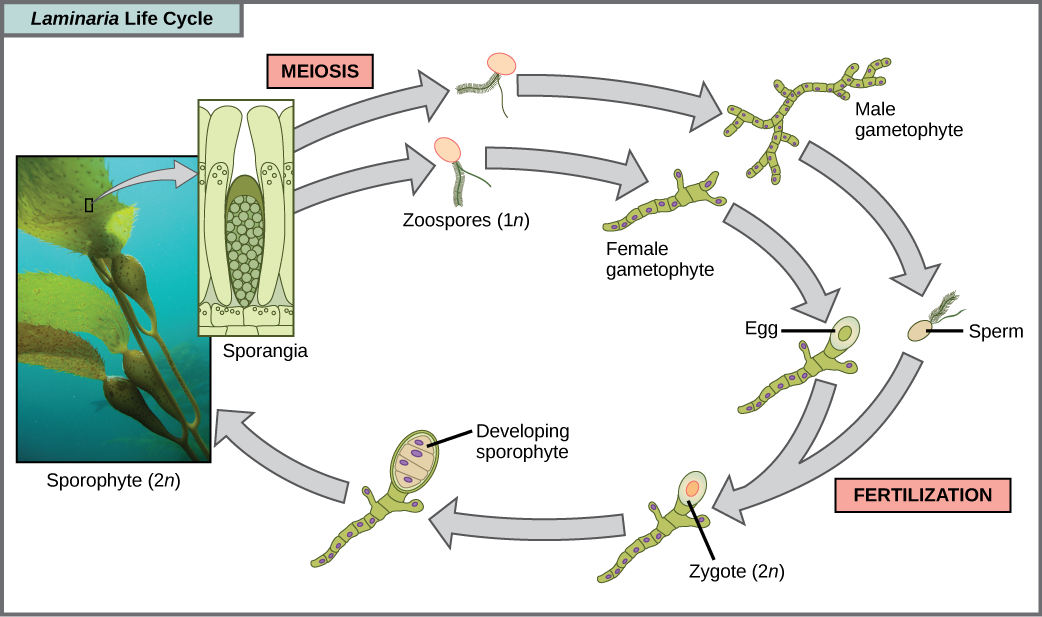
Una variedad de ciclos de vida de algas está representada por los estramenopiles, pero el más complejo es la alternancia de generaciones, en las que tanto las etapas haploide como diploide implican multicelularidad. Compara este ciclo de vida con el de los humanos, por ejemplo. Los gametos haploides producidos por la meiosis (esperma y óvulo) se combinan en la fertilización para generar un cigoto diploide que sufre muchas rondas de mitosis para producir un embrión multicelular y luego un feto. Sin embargo, los espermatozoides y óvulos individuales nunca se convierten en seres multicelulares. Las plantas terrestres también han evolucionado alternancia de generaciones. En el género de algas pardas Laminaria, las esporas haploides se desarrollan en gametofitos multicelulares, los cuales producen gametos haploides que se combinan para producir organismos diploides que luego se convierten en organismos multicelulares con una estructura diferente a la forma haploide (Figura\(\PageIndex{9}\)). Algunos otros organismos, como la alga roja Polysiphonia, realizan alternancia de generaciones en las que tanto la forma haploide como la diploide tienen el mismo aspecto.
Resumen de Características de las Algas Pardas
- Morfología: Talo multicelular
- Composición de la pared celular: Celulosa y alginato de calcio
- Cloroplastos: 4 membranas, los pigmentos son clorofila a, clorofila c y fucoxantina
- Carbohidrato de almacenamiento: Laminarina
- Ciclo de vida: Principalmente diplónico (alternancia de generaciones en algunas especies)
- Ecología: Marina
Bacillariophyceae
Las diatomeas son otro linaje fotosintético de heterocontes fotosintéticos que se derivó del evento endosimbiótico secundario. Las diatomeas son un grupo increíblemente diverso de organismos unicelulares que contienen entre 20,000 y 2 millones de especies. Estos organismos son unicelulares y están rodeados por un frustulo, una cáscara de sílice hecha de dos válvulas distintas que encierran la membrana plasmática. Los frustules son increíblemente intrincados, cubiertos de pequeños poros en un arreglo especialmente adaptado para capturar la luz solar (Figura\(\PageIndex{11}\)). Algunas diatomeas exhiben una hendidura en su cubierta de sílice, llamada rafe. Al expulsar una corriente de mucopolisacáridos del rafe, la diatomea puede unirse a las superficies o impulsarse en una dirección.
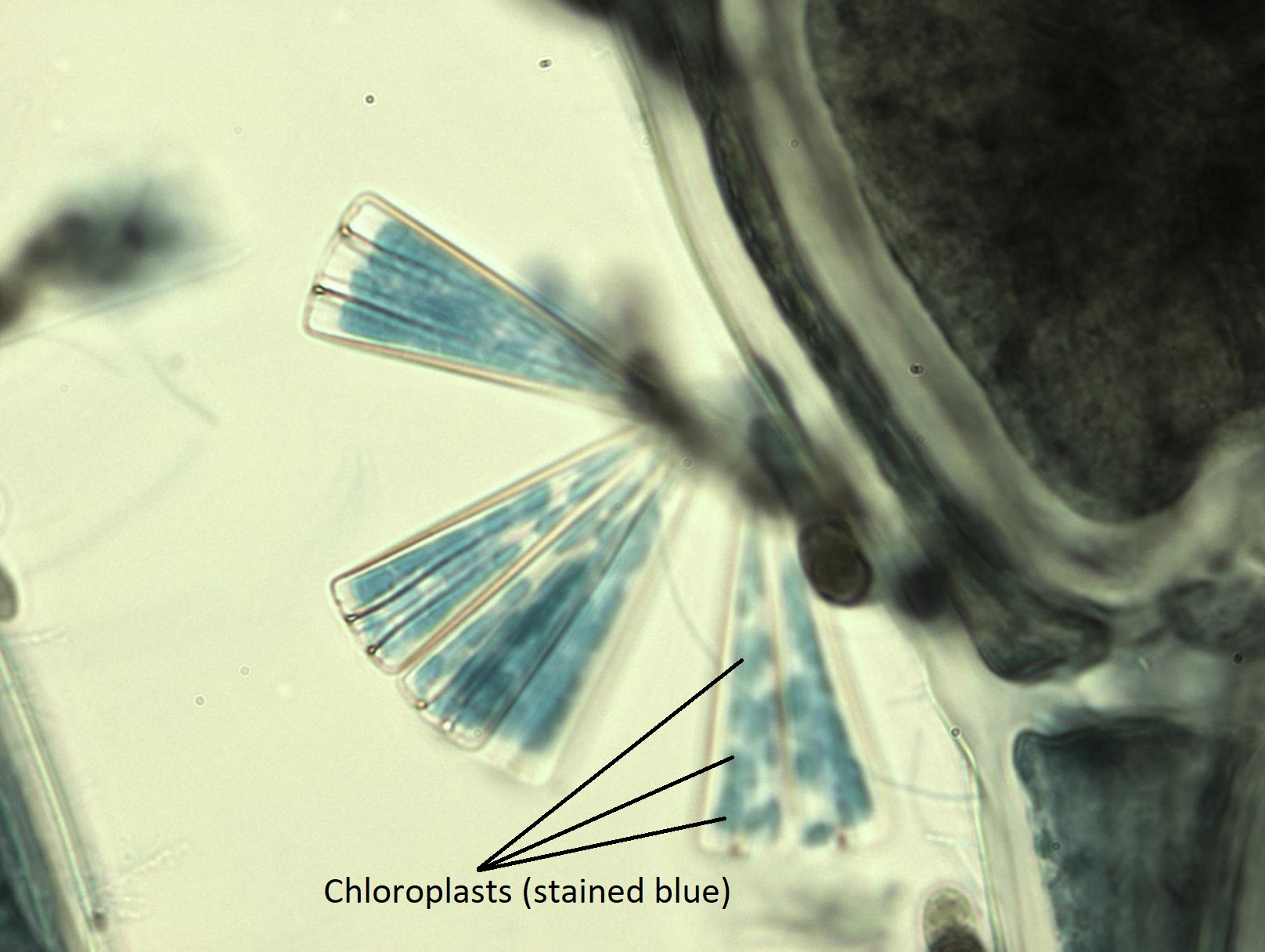
Al igual que las algas pardas, tienen cloroplastos dorados con 4 membranas (Figura\(\PageIndex{10}\)). Las diatomeas almacenan carbohidratos en forma de crisolaminarina. Los frustulos de sílice de diatomeas que se encuentran en los sedimentos (tierra de diatomeas) se utilizan para innumerables fines comerciales, incluyendo aditivos de pasta de dientes (como abrasivo), filtros y aislamiento.


Morfología
Todavía estamos tratando de averiguar cómo determinar qué es una “especie” de diatomeas y, hasta el momento, se han clasificado con base en la morfología de la frustula. Usando esta clasificación, históricamente hubo dos grupos principales de diatomeas: céntricas (tienen simetría radial, ver Figura\(\PageIndex{12}\)) y pennadas (tienen simetría bilateral, ver Figura\(\PageIndex{13}\)). Estas clasificaciones han mejorado y aumentado en complejidad, por lo que aquí cubriremos solo los trazos amplios. Para una mirada más profunda a la clasificación morfológica actual de las diatomeas y a las fantásticas imágenes, consulta este sitio web.






Ecología
Además de la morfología, las diatomeas también pueden clasificarse por dónde ocurren. Las diatomeas flotantes son planctónicas. Las diatomeas unidas a otros organismos (como las algas gigantes) son epífitas (Figura\(\PageIndex{15}\)). Las diatomeas epífitas se pueden encontrar en ecosistemas acuáticos sobre algas y angiospermas acuáticas como la eelgrass, así como ecosistemas terrestres, que viven en las grietas húmedas de la corteza de los árboles. Las diatomeas bentónicas tienden a habitar hacia el fondo de un cuerpo de agua. En general, estas tres categorizaciones se refieren a ecosistemas acuáticos. Sin embargo, las diatomeas se pueden encontrar casi en cualquier lugar donde haya agua en los ecosistemas terrestres. La composición comunitaria de las diatomeas varía según la ubicación. Debido a esto, las diatomeas se han utilizado en investigaciones forenses para determinar dónde se ahogó alguien (dependiendo de las especies de diatomeas presentes) y cuánto tiempo hace que se ahogan (en función de qué tan lejos habían migrado las diatomeas a sus tejidos).
Las diatomeas son los principales productores en ambientes acuáticos; es decir, son responsables de hasta el 40% de la fotosíntesis que ocurre en el agua dulce y en los océanos. Sirven como base principal de las cadenas alimentarias en estos hábitats, aportando calorías a protistas heterótrofos y pequeños animales. Estos, a su vez, alimentan a animales más grandes. Durante los períodos de disponibilidad de nutrientes, las poblaciones de diatomeas florecen a números mayores que los que pueden consumir los organismos acuáticos. Las diatomeas sobrantes mueren y se hunden hasta el fondo marino donde no son fácilmente alcanzadas por los saprotrofos que se alimentan de organismos muertos. Como resultado, el dióxido de carbono que las diatomeas habían consumido e incorporado a sus células durante la fotosíntesis no se devuelve a la atmósfera. En general, este proceso por el cual el carbono es transportado profundamente al océano se describe como la bomba biológica, debido a que el carbono es “bombeado” a las profundidades oceánicas donde es inaccesible a la atmósfera como dióxido de carbono. La bomba de carbono biológico es un componente crucial del ciclo del carbono que mantiene niveles más bajos de dióxido de carbono atmosférico.
Reproducción
Las diatomeas se reproducen principalmente asexualmente por fisión binaria, similar a los procariotas. Durante la fisión binaria, las dos válvulas del frustulo se separan y cada nueva celda forma una nueva válvula dentro de la antigua. Sin embargo, la nueva válvula siempre es más pequeña. Si las diatomeas solo se reproducen de esta manera, se traduce en una disminución continua del tamaño promedio. Cuando se alcanza algún tamaño mínimo, esto puede desencadenar la reproducción sexual. Cuando las diatomeas se reproducen sexualmente, tienen un ciclo de vida diplónico y producen una auxosporas muy grandes (Figura\(\PageIndex{16}\)).

Diversidad
Video\(\PageIndex{1}\): Este video muestra parte de la increíble diversidad de formas de diatomeas y el increíble arte que Klaus Kemp hace con ellas. Se obtiene de YouTube.
Resumen de Características para Diatomeas
- Morfología: Unicelular
- Composición de la pared celular: Frustule de sílice
- Cloroplastos: 4 membranas, los pigmentos son clorofila a, clorofila c y fucoxantina
- Carbohidrato de almacenamiento: Crisolaminarina
- Ciclo de vida: Diplontic
- Ecología: En todas partes! Marina, de agua dulce y terrestre.
Resumen
Aunque las algas pardas y las diatomeas parecen tener muy poco en común morfológicamente, descienden de un ancestro común. Ambos grupos tienen un ciclo de vida diplónico durante alguna etapa del cual una célula tendrá flagelos heterocónicos. Tienen cloroplastos de 4 membranas que contienen los pigmentos clorofila a, clorofila c y fucoxantina. Este último pigmento da a los cloroplastos de estos grupos un color dorado. Se trata de donde terminan las similitudes.
Las algas pardas son exclusivamente multicelulares y se encuentran en hábitats marinos, más típicamente en la zona intermareal. Sus paredes celulares contienen celulosa y almacenan sus carbohidratos como laminarina.
Las diatomeas son exclusivamente unicelulares y se encuentran en casi todos los hábitats donde hay agua. Su celda única está rodeada por un frustulo de sílice compuesto por dos válvulas distintas. Almacenan sus carbohidratos como crisolaminarina.
Atribución
Curada y autoría de Maria Morrow, CC-BY-NC, utilizando las siguientes fuentes:
- 19.1.2 Protistas de la Biología por John. W. Kimball (con licencia CC-BY)
- 23.3 Grupos de Protistas de Biología 2e por OpenStax (licenciado CC-BY). Accede gratis en openstax.org.