
5.3.3: Algas rojas y verdes
- Page ID
- 58334

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Distinguir entre diferentes grupos de algas usando ciclo de vida, características morfológicas y composición celular.
- Conectar adaptaciones en las algas rojas y verdes con las características del hábitat y la ecología.
- Identificar estructuras y fases en los ciclos de vida de Polisifonía y Spirogyra; conocer la ploidía de estas estructuras.
Las algas rojas y verdes están incluidas en el supergrupo Archaeplastida. Fue a partir de un ancestro común de estos protistas que las plantas terrestres evolucionaron, ya que sus parientes más cercanos se encuentran en este grupo. La evidencia molecular sostiene que todos los Archaeplastida son descendientes de una relación endosimbiótica entre un protista heterotrófico y una cianobacteria. ¡Esta endosimbiosis primaria resultó en los cloroplastos de todos los eucariotas fotosintéticos discutidos en este libro! Las algas rojas y verdes incluyen formas unicelulares, multicelulares y coloniales.
Glaucophyta
Las glaucófitas son eucariotas unicelulares y fototróficas que se encuentran en ecosistemas de agua dulce. Los glaucófitos son probablemente una rama ancestral en la Archaeplastida y a menudo se utilizan como evidencia del origen cianobacteriano de los cloroplastos. Tienen orgánulos similares a cloroplastos, llamados cianelos o muroplastos, que tienen peptidoglicano entre las dos membranas. Tienen los mismos pigmentos que las cianobacterias y las algas rojas: clorofila a y ficobilinas. Al igual que las cianobacterias, aparecen azul-verdes debido a la interacción de clorofila y ficocianinas (glauco- proviene del griego glaukos que significa azul-verde). Las celdas no móviles tienen una pared celular rígida compuesta de celulosa. Las células móviles tienen dos flagelos de latigazo. La reproducción sexual aún no se ha documentado en este grupo.

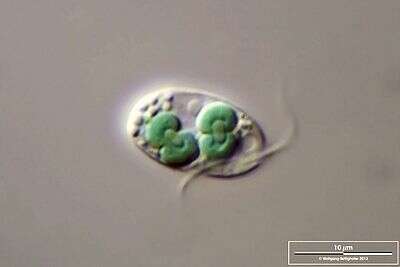
Rhodophyta
Las algas rojas descendieron del mismo evento endosimbiótico que la Glaucophyta. Las algas rojas son casi exclusivamente marinas. Algunos son unicelulares pero la mayoría son multicelulares. Se han identificado aproximadamente 6,000 especies. Tienen cloroplastos verdaderos con dos membranas (sin peptidoglicano remanente) que contienen clorofila a Al igual que las cianobacterias, utilizan ficobilinas como pigmentos de antena: ficoeritrina (que las hace rojas) y ficocianina. El pigmento rojo permite que las algas rojas se fotosinteticen a profundidades más profundas que las algas verdes o pardas, aprovechando más ondas de luz azul que penetran más profundamente en la columna de agua. A diferencia de las algas verdes y las plantas, las algas rojas almacenan carbohidratos como almidón de la Florida en el citosol. Algunos se utilizan como alimento en las regiones costeras de Asia. El agar, la base para el cultivo de bacterias y otros microorganismos, se extrae de un alga roja.
Selección de presiones y conductores
Un aspecto importante para comprender los rasgos de la historia de vida de la Rhodophyta es comprender los desafíos de vivir en un ambiente marino.
- Acceso a la luz solar: La mayoría de los colores de luz no pueden penetrar en aguas más profundas, ya que son dispersos por moléculas de agua. Las longitudes de onda de la luz que llegan a lo más profundo del océano son azules y verdes. Muchos peces que viven en las profundidades del océano son de color rojo. Debido a que la luz roja no penetra hasta las profundidades donde viven, esto los hace prácticamente indetectables a la vista. Recuerden, vemos las cosas por la luz que rebota en ellas. Los pigmentos rojos reflejan la luz roja, por lo que no hay luz roja, ni luz reflejada. Las algas rojas están usando una estrategia similar —absorben las longitudes de onda de la luz que no son rojas— con un objetivo diferente: usar esa luz absorbida para hacer comida. La ficoeritrina en sus cloroplastos refleja la luz roja, dándoles un aspecto rojo, y absorbe la luz azul que es capaz de penetrar en áreas más profundas de la columna de agua.
- Fertilización: El océano es un ambiente expansivo, a menudo con grandes áreas de espacio abierto entre poblaciones de organismos. En este ambiente, la fertilización exitosa de un óvulo por un espermatozoide no móvil, las algas rojas carecen de flagelos, presenta un desafío. Tener fases multicelulares haploides y diploides proporciona a las algas rojas más oportunidades para producir gametos y esporas. Una etapa diploide que clona el cigoto, el carposporófito, brinda más oportunidades para hacer meiosis a partir de cada evento de fertilización.
- Salinidad: Los ambientes marinos son relativamente altos en salinidad. Una posible adaptación para esto es tener polisacáridos sulfatados en la pared celular, como los galactanos presentes en Rhodophyta. Esta es una estrategia presente en (potencialmente todas) algas marinas y se infiere que es una adaptación para la tolerancia a la salinidad. Consulte este artículo de acceso abierto para obtener más información.

Morfología
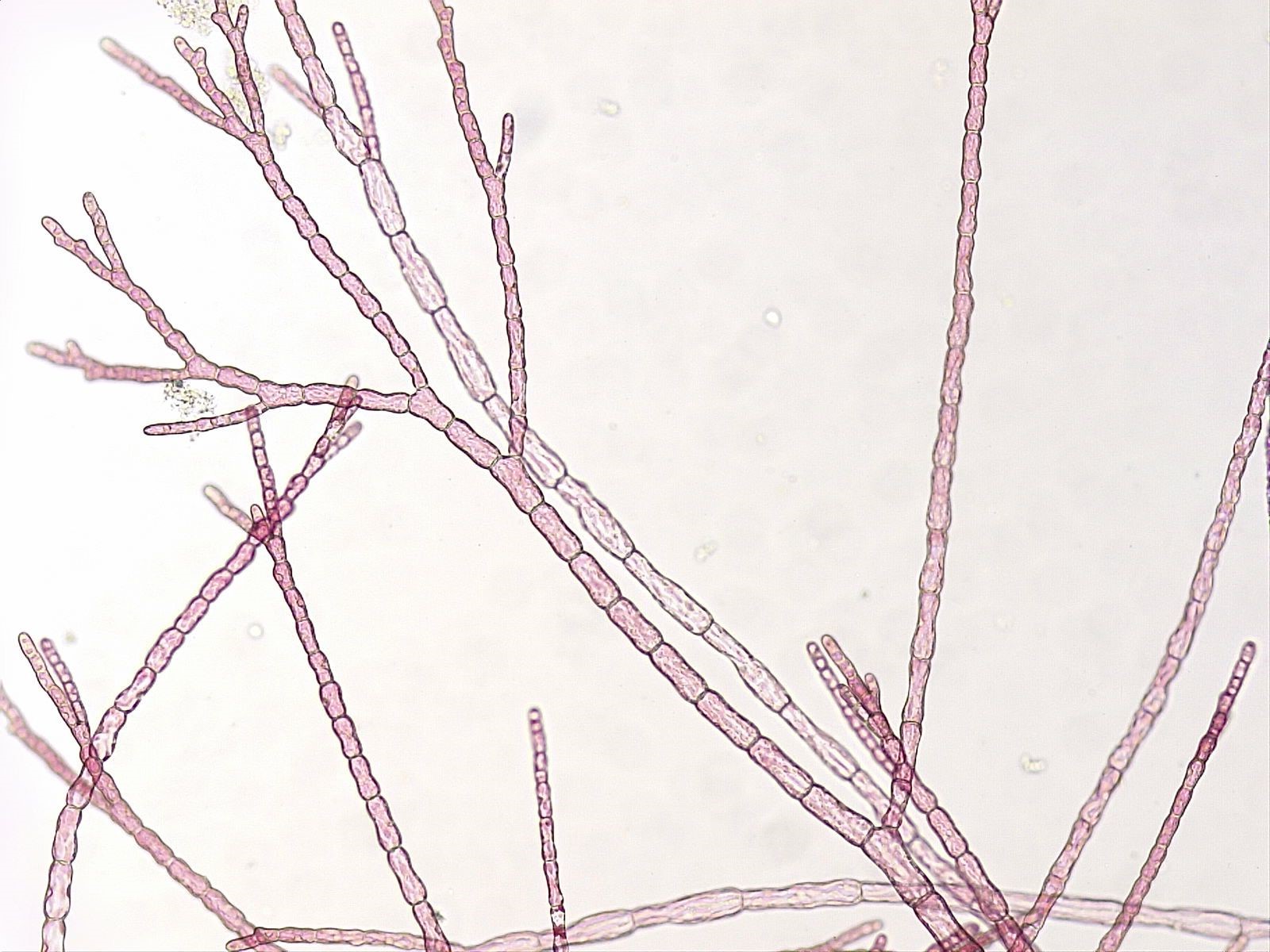
Las algas rojas tienen una amplia gama de morfologías. Las formas unicelulares pueden vivir solitariamente o como colonias pero, a diferencia de otros miembros de la Archaeplastida, carecen de flagelos. Los flagelos están ausentes de la Rhodophyta, perdidos en algún momento de su historia evolutiva. Las formas multicelulares pueden ser filamentosas, frondosas, laminares, coraloides o incluso costras (algunos ejemplos en Figura\(\PageIndex{4}\) y Figura\(\PageIndex{5}\)). Las extrañas algas rojas coralinas tienen depósitos calcerosos en las paredes celulares que hacen que el talo sea duro, como un coral. Estos pueden tomar una variedad de formas y son capaces de vivir a profundidades que otras algas no pueden (¡más de 500 pies de profundidad para algunas!).



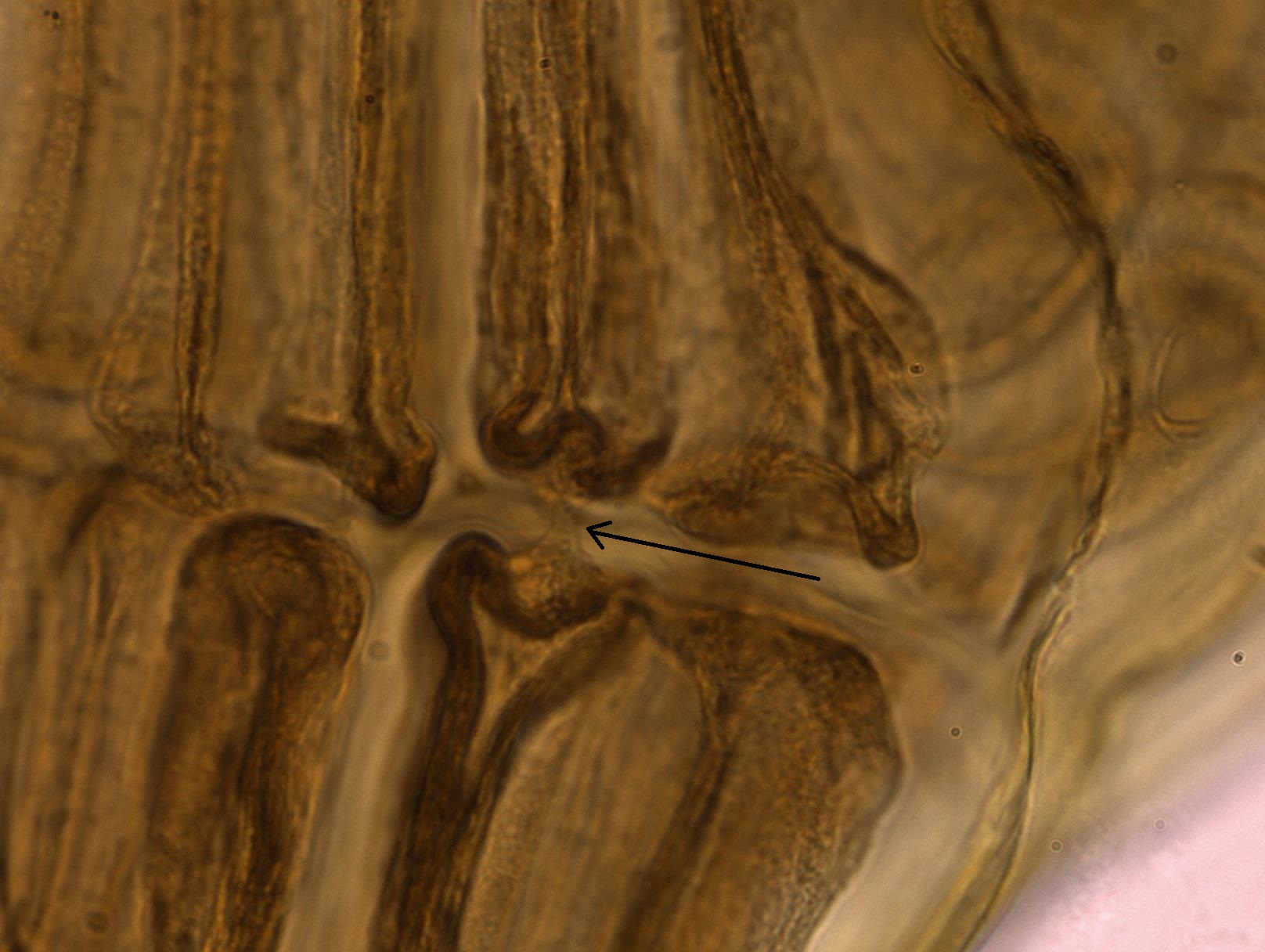
Las células de especies multicelulares se conectan a través de citocinesis incompleta, lo que resulta en conexiones de foso (Figura\(\PageIndex{6}\)).
Ciclo de vida de la polisifonía
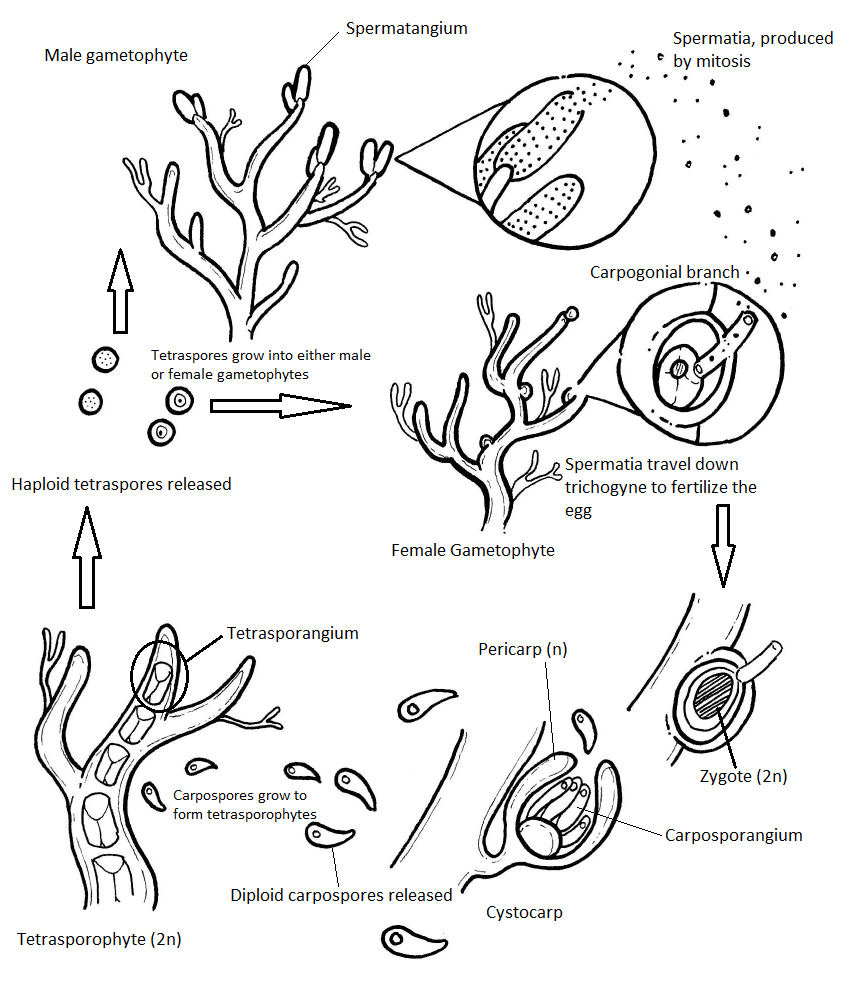
Las algas rojas tienen un ciclo de vida haplodiplónico (alternancia de generaciones) que tiene una etapa diploide extra: el carposporófito. La polisifonía es el organismo modelo para el ciclo de vida de Rhodophyta. Los gametofitos de Polisifonía son isomórficos (iso- significa mismo, forma de transformación), es decir, tienen la misma morfología básica. Cualquier diferencia que veas en la coloración de las imágenes en esta sección se debe a la tinción. Todos aparecerían de color rojo intenso en una diapositiva sin manchar.

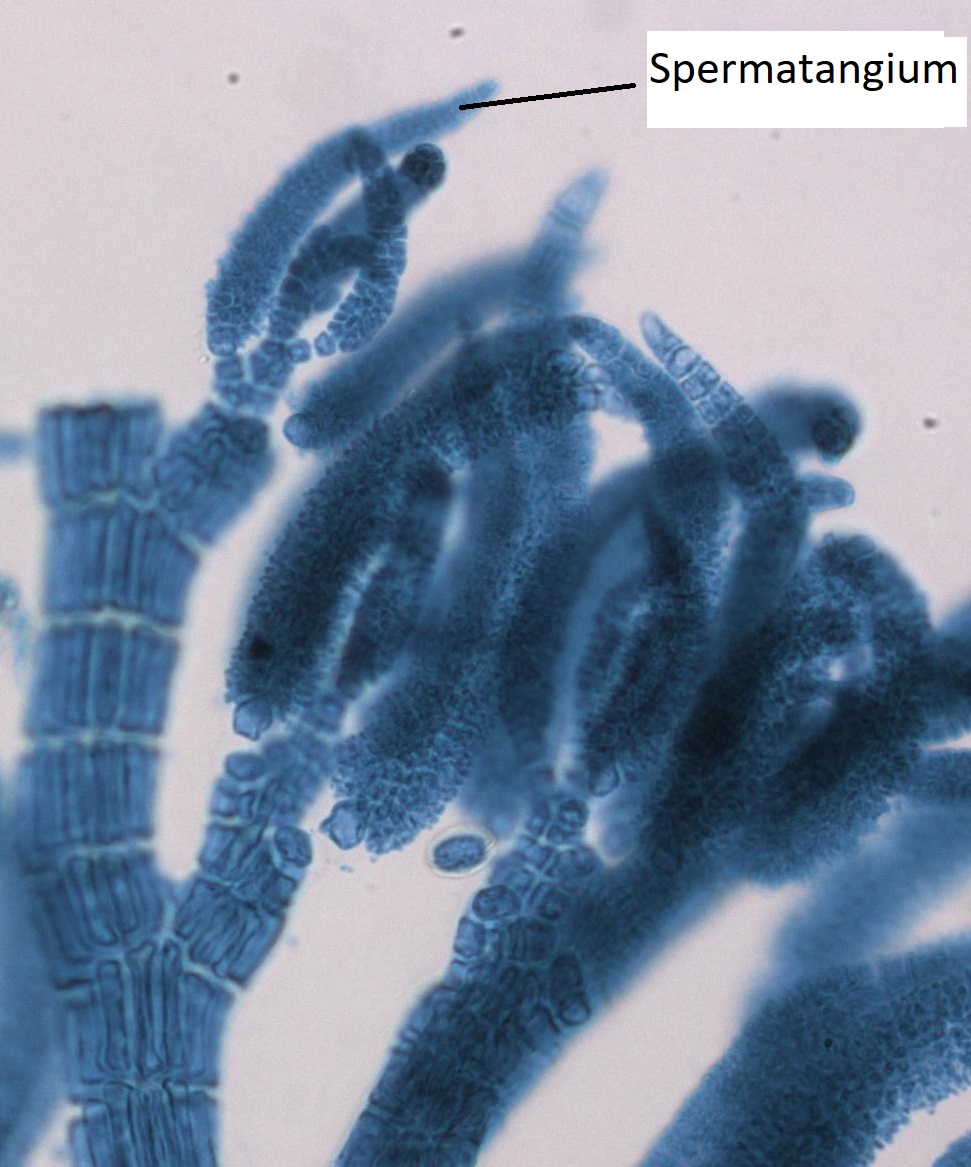
Gametofitos Masculinos
El gametofito masculino tiene estructuras alargadas que emergen de las puntas de las ramas del talo. Se trata de espermatangios, donde las espermatias son producidas por la mitosis.
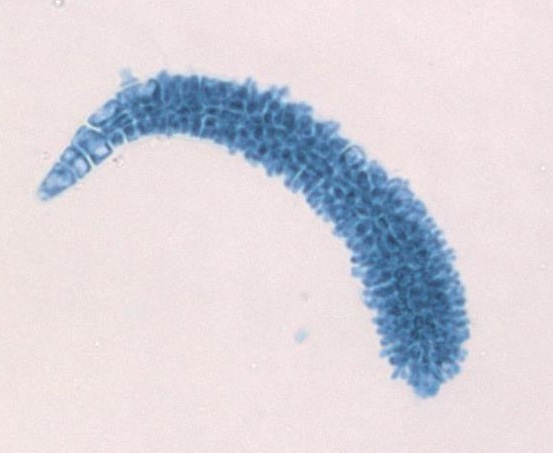
Gametofitos femeninos y carposporófitos
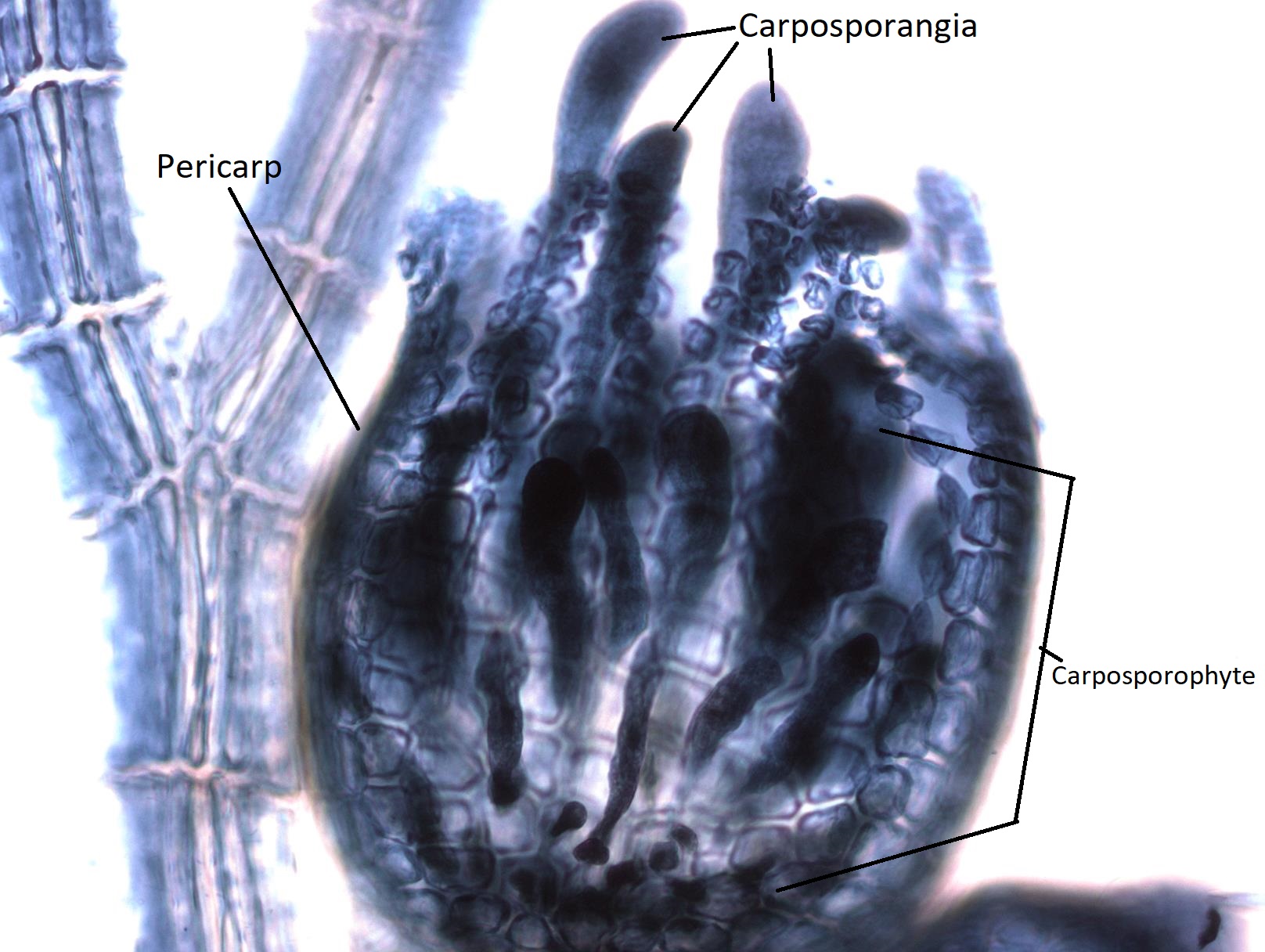
El gametofito hembra produce un óvulo que está contenido dentro de una estructura llamada carpogonio. Esta estructura tiene una proyección larga y delgada llamada trichogyne (trich- que significa pelo, -gyne que significa hembra). Durante la fertilización, un espermatium se fusiona con la tricogina y el núcleo del espermatium viaja por el tubo hasta el óvulo. Cuando el núcleo del espermatio se fusiona con el huevo, se produce un cigoto. Este cigoto es retenido y nutrido por el gametofito femenino a medida que crece.
Las estructuras globosas que se ven creciendo a partir del talo gametofítico femenino se llaman cistocarpos. Un cistocarpo está compuesto tanto por tejido gametofito femenino (n) como tejido carposporófito (2n). La capa externa del cistocarpio, el pericarpio (peri- significado alrededor) se deriva del gametofito femenino y es haploide. El interior del cistocarpio consiste en el carposporófito, que es diploide, y produce estructuras llamadas carposporangias, dentro de las cuales produce carposporas por mitosis. Todos estos, carposporófitos, carposporangios y carposporos, son diploides.
Tetrasporófito
Las carposporas diploides son liberadas a las aguas oceánicas, donde serán transportadas por corrientes a otro lugar. Si una carpospora aterriza en un ambiente apropiado, crecerá por mitosis hasta convertirse en un tetrasporófito (2n). El tetrasporófito produce tetrasporangios (2n) dentro de las ramas del talo. Cada tetrasporanjo produce cuatro tetrasporas haploides únicas por meiosis. Las tetrasporas (n) son liberadas y crecerán por mitosis en gametofitos masculinos o femeninos, completando el ciclo de vida.
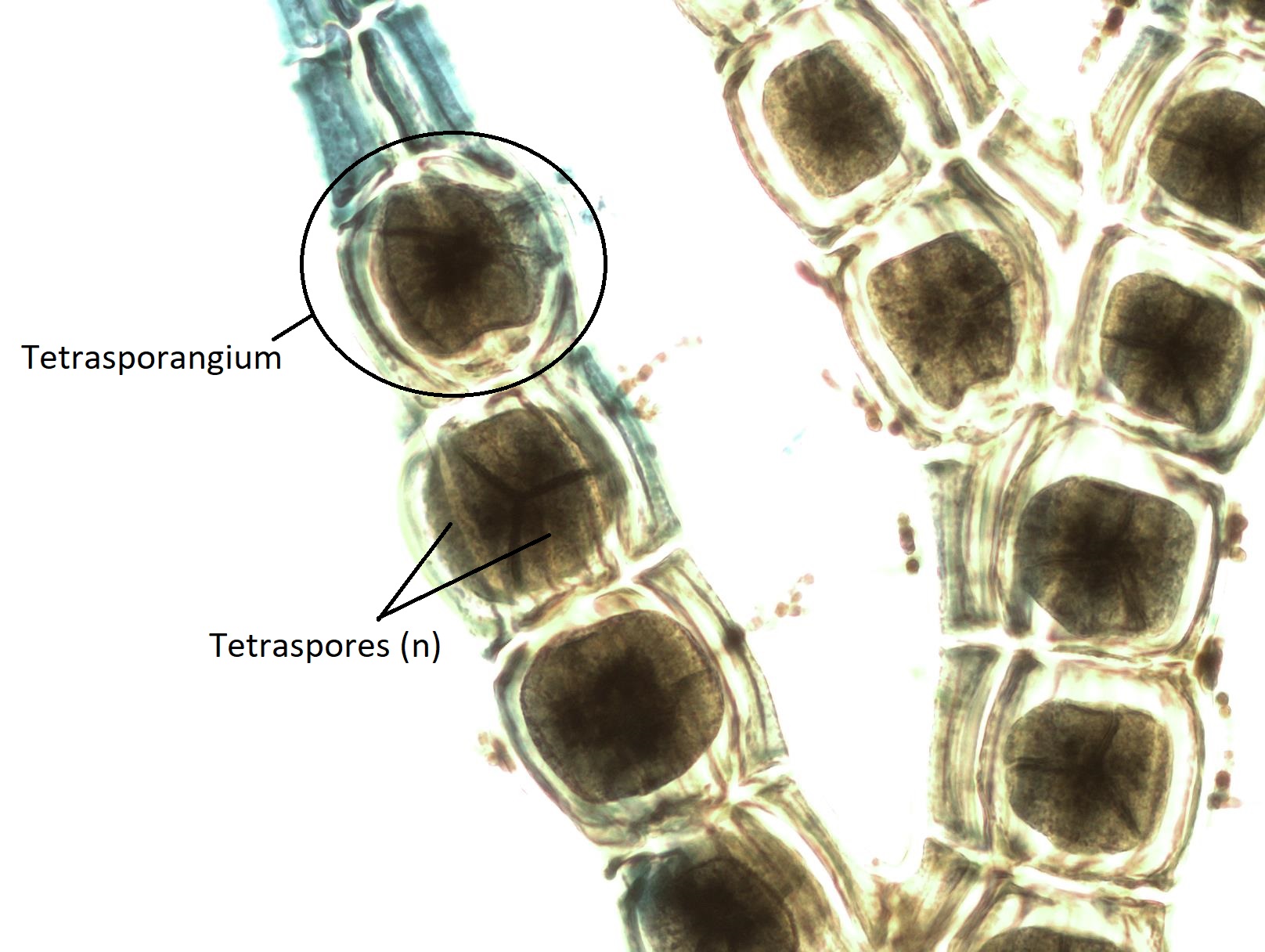
Diagrama de ciclo de vida completo
Resumen de Características para Algas Rojas
- Morfología: De unicelular a multicelular, sin estadios flagelados. Las células de especies multicelulares se conectan a través de citocinesis incompleta, lo que resulta en conexiones de foso.
- Composición de la pared celular: Celulosa y galactanos
- Cloroplastos: 2 membranas, los pigmentos son clorofila a y ficobilinas (principalmente ficoeritrina, aportando su color rojo)
- Almacenamiento de carbohidratos: Almidón Florideano
- Ciclo de vida: Alternancia de generaciones con una etapa diploide extra, el carposporófito
- Ecología: Principalmente marina (97% de las especies)
Algas Verdes
El grupo de algas más abundante son las algas verdes. La naturaleza de las relaciones evolutivas entre las algas verdes sigue siendo objeto de debate. A partir de 2019, los datos genéticos apoyan la división de las algas verdes en dos linajes principales: clorofitos y estreptofitos. Las estreptofitas incluyen varios linajes de algas verdes (como los charófitos) y todas las plantas terrestres. Las estreptofitas y clorofitas representan un grupo monofilético llamado Viridiplantae (literalmente “plantas verdes”). Las algas verdes presentan características similares a las plantas terrestres, particularmente en términos de estructura de cloroplastos. Tienen clorofila a y b, han perdido ficobilinas pero ganado carotenoides, y almacenan carbohidratos como almidón dentro de los plastidios. Aunque algunas de las formas multicelulares son grandes, nunca desarrollan más de unos pocos tipos de células diferenciadas y sus óvulos fertilizados no se convierten en un embrión.
Las algas verdes son una fuente importante de alimento para muchos animales acuáticos. Cuando los lagos y estanques son “fertilizados” con fosfatos y nitratos (por ejemplo, de aguas residuales y escorrentía de campos y céspedes fertilizados), las algas verdes a menudo forman extensas “flores” de algas. Los miembros de este grupo se pueden encontrar en hábitats de agua dulce y marinos, y muchos se han adaptado a la vida en tierra, ya sea dentro de líquenes o vida libre (ver Figura\(\PageIndex{12}\)).

Selección de presiones y conductores
- Daños Solares. Las algas verdes representan un grupo diverso de organismos con diversos rasgos de historia de vida, muchos de los cuales se comparten con plantas terrestres. El desarrollo de carotenoides —pigmentos amarillos, naranjas y rojos que actúan tanto en la recolección de luz como en la protección solar— ofrece a este grupo un mayor acceso a la luz solar al tiempo que protege contra el daño UV. Los rayos UV no penetran muy lejos en la columna de agua, por lo que los organismos que se mueven hacia aguas menos profundas o ambientes terrestres necesitarían hacer frente a este nuevo desafío. Muchas especies terrestres de algas verdes aparecen anaranjadas, en lugar de verdes, debido a la producción de grandes cantidades de carotenoides.
Morfología
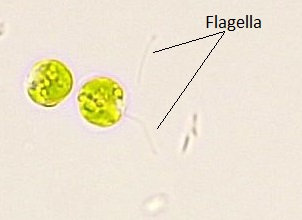
Estas algas presentan una gran diversidad de forma y función. Similar a las algas rojas, las algas verdes pueden ser unicelulares o multicelulares. Muchas especies unicelulares forman colonias y algunas algas verdes existen como células grandes, multinucleadas, individuales. Las algas verdes habitan principalmente agua dulce y suelo húmedo, y son un componente común del plancton. La clamidomonas es una clorofita simple, unicelular, con morfología en forma de pera y dos flagelos opuestos anteriores que lo guían hacia la luz percibido por su mancha ocular (Figura\(\PageIndex{13}\)). Las especies más complejas exhiben gametos haploides y esporas que se asemejan a Chlamydomonas.
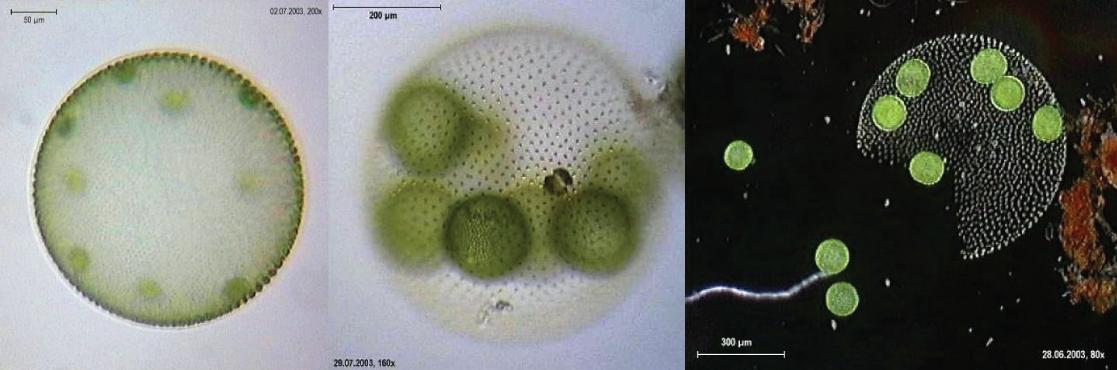
El alga Volvox es uno de un organismo colonial, que se comporta de alguna manera como una colección de células individuales, pero de otras formas como las células especializadas de un organismo multicelular (Figura\(\PageIndex{14}\)). Las colonias Volvox contienen de 500 a 60,000 células, cada una con dos flagelos, contenidas dentro de una matriz hueca y esférica compuesta por una secreción de glicoproteína gelatinosa. Las células Volvox individuales se mueven de manera coordinada y están interconectadas por puentes citoplásmicos. Sólo unas pocas de las células se reproducen para crear colonias hijas, ejemplo de especialización celular básica en este organismo.
Volvox puede reproducirse tanto asexualmente como sexualmente. En la reproducción asexual, los gonidios se convierten en nuevos organismos que se desprenden del progenitor (que luego muere). En la reproducción sexual, la presencia de una sustancia química inductora provoca lo siguiente:
- Los gonidios de los machos se convierten en racimos de espermatozoides.
- Los gonidios de las hembras se desarrollan en nuevas esferas cada una de cuya gonidia propia se desarrolla en un par de huevos.
- Los espermatozoides salen del progenitor masculino y nadan hasta la hembra donde fertilizan sus óvulos.
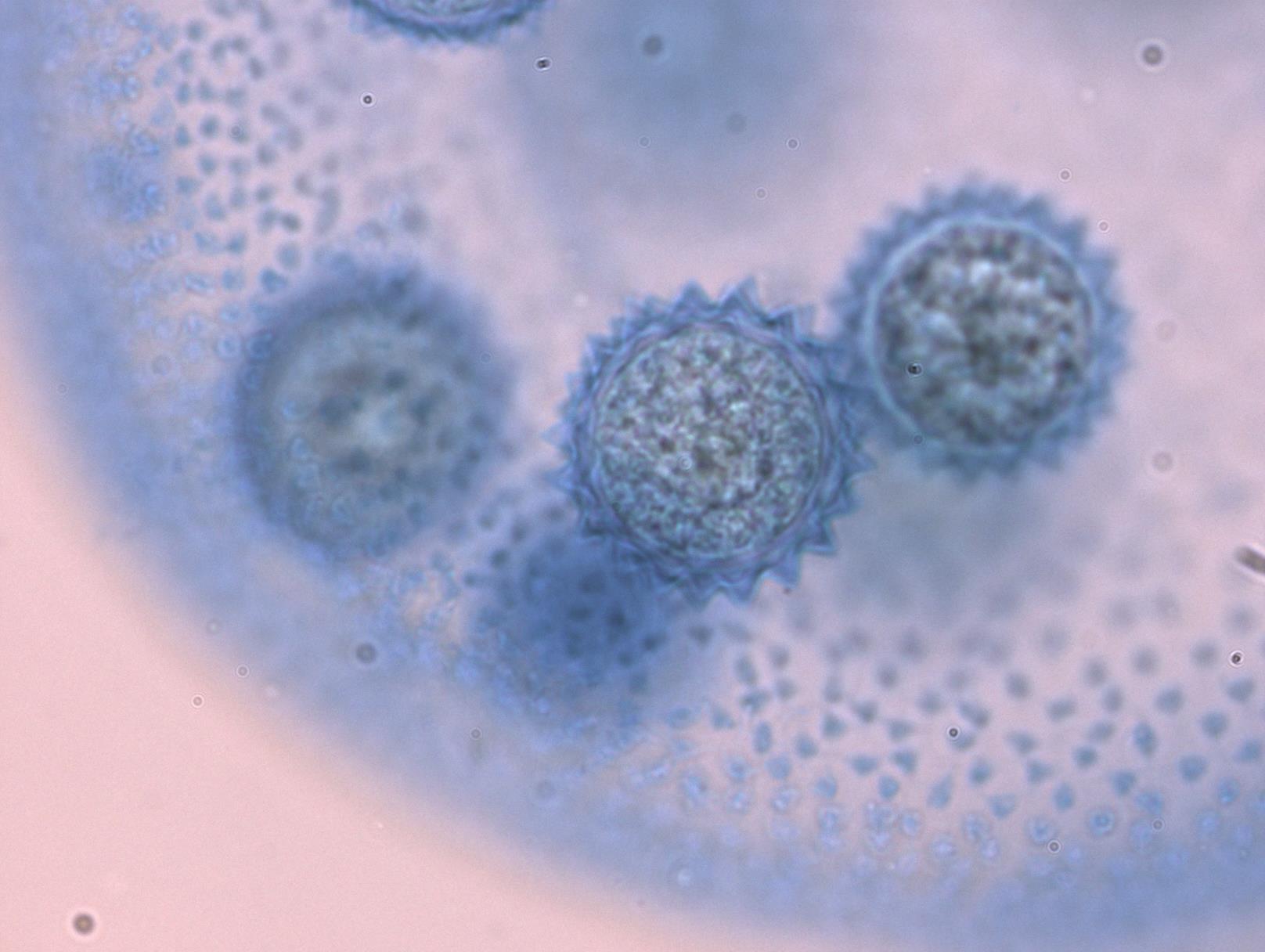
- Los cigotos forman una etapa de reposo que permite a Volvox sobrevivir a duras condiciones (Figura\(\PageIndex{15}\)).
Video\(\PageIndex{1}\): Este video muestra cómo ocurre la reproducción sexual en la alga verde colonial Volvox. Fuente de YouTube.
El genoma de Volvox carteri consta de 14,560 genes que codifican proteínas, ¡solo 4 genes más que en el unicelular Chlamydomonas reinhardtii! La mayoría de sus genes también se encuentran en Chlamydomonas. Los pocos que no son codifican las proteínas necesarias para formar la matriz extracelular masiva de Volvox.
Las especies del género Caulerpa exhiben follaje aplanado similar a helecho y pueden alcanzar longitudes de 3 metros (Figura\(\PageIndex{16}\)). Las especies de Caulerpa experimentan división nuclear, pero sus células no completan la citocinesis, permaneciendo en su lugar como células individuales masivas y elaboradas.

Los verdaderos organismos multicelulares, como la lechuga de mar, Ulva, también están representados entre las algas verdes (Figura\(\PageIndex{17}\) y Figura\(\PageIndex{18}\)).


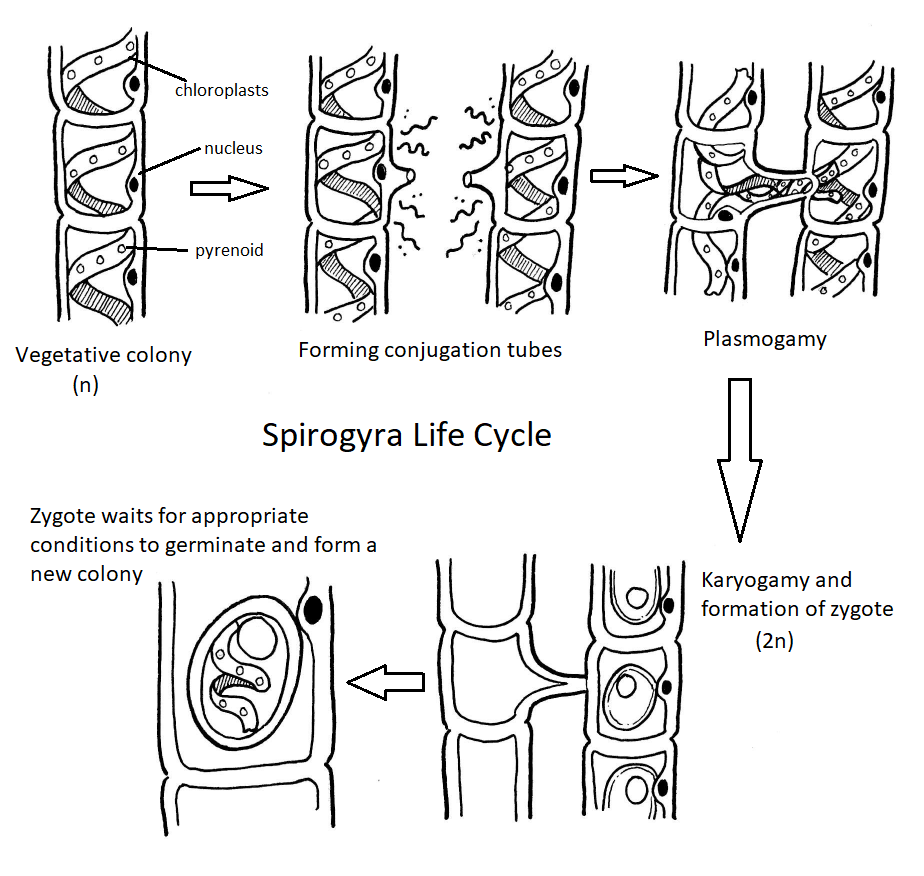
Ciclo de vida de Spirogyra
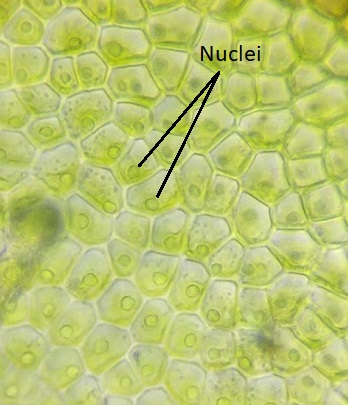
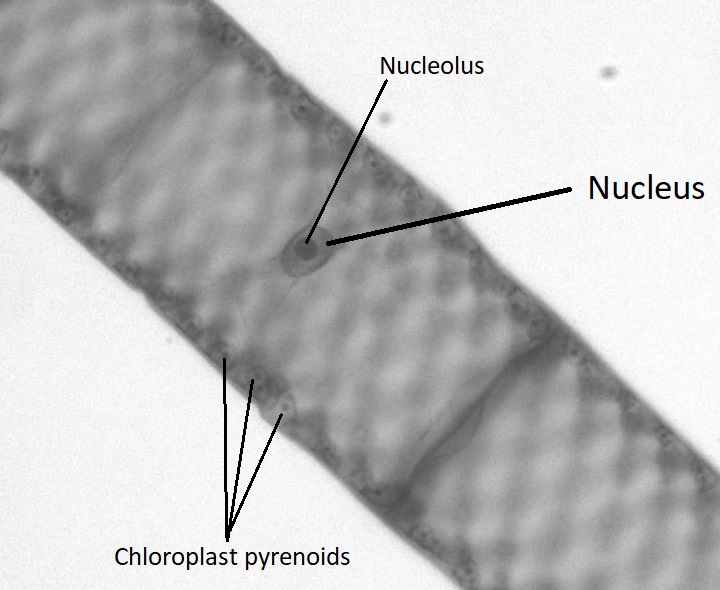
Aunque las algas verdes muestran una diversidad de ciclos de vida, muchas tienen un ciclo de vida haplónico. Un organismo modelo para las algas verdes es Spirogyra (Figura\(\PageIndex{19}\)). La espirogyra es una alga verde unicelular que crece en colonias largas y filamentosas, haciéndola parecer un organismo multicelular. A pesar de que es técnicamente unicelular, su naturaleza colonial nos permite clasificar su ciclo de vida como haplónico. En las células vegetativas haploides de la colonia, los cloroplastos se disponen en espirales, conteniendo regiones oscurecidas llamadas pirenoides donde ocurre la fijación de carbono. Cada célula haploide del filamento es un individuo, lo que hace que la reproducción sexual entre colonias sea un proceso interesante.
Cuando se encuentran dos colonias de Spirogyra que son de un tipo de apareamiento complementario (+/-), se produce la reproducción sexual. Las dos colonias se alinean, cada célula frente a una célula complementaria en el otro filamento. Un tubo de conjugación se extiende desde cada célula en una colonia (Figura\(\PageIndex{20}\)), induciendo la formación de un tubo sobre las células de la otra colonia. Los tubos de conjugación de cada colonia se fusionan.

El contenido de una célula se moverá a través del tubo de conjugación y se fusionará con el contenido de la célula complementaria, dando como resultado un cigoto diploide (Figura\(\PageIndex{21}\)). El cigoto aparece como una gran estructura similar a un huevo contenida dentro de la célula complementaria. Tiene una pared gruesa que proporciona resistencia a la desecación y al frío, permitiendo que las colonias de Spirogyra hibernen, cuando sea necesario. La otra colonia es ahora un filamento de celdas vacías que serán descompuestas por algún descomponedor. Cuando las condiciones son adecuadas, el cigoto se somete a meiosis para producir otra colonia vegetativa de células haploides.
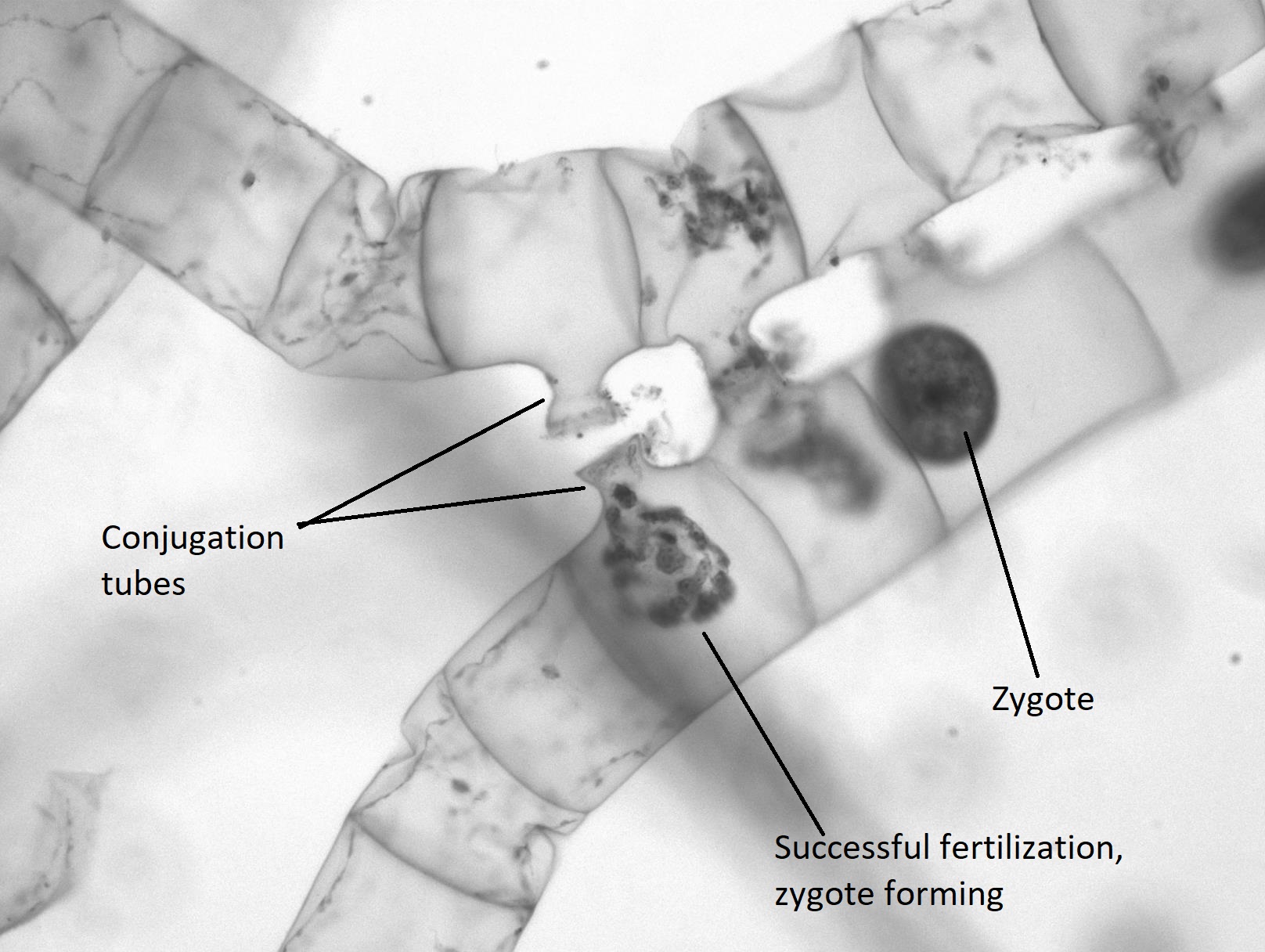
Diagrama de ciclo de vida completo
Resumen de Características de las Algas Verdes
- Morfología: De unicelular a multicelular; dos flagelos de latigazo cervical en células móviles
- Composición de la pared celular: Celulosa
- Cloroplastos: 2 membranas, los pigmentos son clorofila a, clorofila b y carotenoides
- Almacenamiento de carbohidratos: Almidón
- Ciclo de vida: Varía, pero principalmente haplontico. Algunas especies marinas tienen alternancia de generaciones.
- Ecología: Especies de agua dulce, marinas y terrestres
Resumen
Glaucófitas, algas rojas y algas verdes forman parte de la Archaeplastida. Estos organismos descienden del mismo evento de endosimbiosis primaria. Se cree que los glaucófitos son uno de los primeros linajes en divergir debido a la presencia de peptidoglicano remanente entre las membranas de sus cianelas similares a cloroplastos. Como era de esperar, las glaucófitas y las algas rojas comparten los mismos pigmentos que las cianobacterias.
Las algas rojas (phylum Rhodophyta) están unidas por varias sinapomorfias (características derivadas compartidas). Carecen de flagelos, tienen conexiones de foso entre las células y almacenan carbohidratos como almidón floridano. Los galactanos sulfatados en sus paredes celulares les permiten aumentar la aptitud en ambientes marinos, mientras que el pigmento ficoeritrina les permite fotosintetizar más profundamente en la columna de agua. Tienen una alternancia de ciclo de vida de generaciones con una fase diploide extra, el carposporófito, que clona el cigoto. Estas características pueden estar relacionadas con los factores estresantes ambientales que presentan los hábitats marinos en los que se encuentran la mayoría de las algas rojas.
Las algas verdes representan varios linajes distintos. Al igual que las plantas, almacenan carbohidratos como almidón dentro de sus plastidios y tienen los pigmentos clorofila a y b, así como carotenoides. Los organismos de este grupo tienen ciclos de vida haplonticos (por ejemplo, Spirogyra) o haplodiplonticos (por ejemplo, Ulva). Muchas algas verdes son unicelulares, formando colonias complejas. Las algas verdes se pueden encontrar en ambientes marinos, de agua dulce y terrestres (¡incluso dentro de líquenes!).
Atribuciones
Curada y autoría de Maria Morrow, CC BY-NC, utilizando las siguientes fuentes:
- 19.1.2 Protistas, 19.1.4 Volvox, y 19.1.5 Diversidad y Relaciones Evolutivas de las Plantas de Biología por John. W. Kimball (con licencia CC-BY)
- 23.3 Grupos de Protistas de Biología 2e por OpenStax (licenciado CC-BY). Accede gratis en openstax.org.