
9.3: Tejidos vegetales
- Page ID
- 58682

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describir la diferencia entre tejidos meristemáticos y no meristemáticos.
- Comparar y contrastar tejido dérmico, suelo y vascular.
Las plantas son eucariotas multicelulares con sistemas tisulares hechos de diversos tipos celulares que realizan funciones específicas. Los tejidos vegetales están compuestos por células que son similares y realizan una función específica. Juntos, los tipos de tejidos se combinan para formar órganos. Cada órgano en sí es también específico para una función particular.
Los sistemas de tejidos vegetales se dividen en uno de dos tipos generales: tejido meristemático y tejido permanente (o no meristemático). Las células del tejido meristemático se encuentran en los meristemas, que son regiones vegetales de división celular continua y crecimiento. Las células de tejido meristemático son indiferenciadas o incompletamente diferenciadas, y continúan dividiéndose y contribuyendo al crecimiento de la planta. Por el contrario, el tejido permanente consiste en células vegetales que ya no se dividen activamente.
Los tejidos meristemáticos constan de tres tipos, en función de su ubicación en la planta. Los meristemos apicales contienen tejido meristemático localizado en las puntas de tallos y raíces, lo que permite que una planta se extienda en longitud. Los meristemos laterales facilitan el crecimiento en grosor o contorno en una planta en maduración. Los meristemos intercalares ocurren solo en monocotiledóneas, en las bases de las láminas foliares y en los nódulos (las áreas donde las hojas se unen a un tallo). Este tejido permite que la hoja monocotiledónea aumente de longitud desde la base de la hoja; por ejemplo, permite que las hojas de césped se alarguen incluso después de siega repetidas.
Los meristemas producen células que se diferencian rápidamente o se especializan, y se convierten en tejido permanente. Dichas células asumen roles específicos y pierden su capacidad de dividirse aún más. Se diferencian en tres tipos principales: tejido dérmico, vascular y tejido molido. El tejido dérmico cubre y protege la planta. El tejido molido sirve como sitio para la fotosíntesis, proporciona una matriz de soporte para el tejido vascular y ayuda a almacenar agua y azúcares. El tejido vascular transporta agua, minerales y azúcares a diferentes partes de la planta. El tejido molido es un tejido simple, lo que significa que cada tejido molido consiste en un solo tipo de célula. Los tejidos dérmicos y vasculares son tejidos complejos porque consisten en múltiples tipos de células.
Tejido dérmico
El tejido dérmico cubre la planta y se puede encontrar en la capa externa de raíces, tallos y hojas. Sus principales funciones son la transpiración, el intercambio de gases y la defensa. La epidermis es un ejemplo de tejido dérmico (Figura\(\PageIndex{1}\)). Se compone de una sola capa de células de la epidermis. Puede contener estomas y celdas de guarda que permiten el intercambio de gases. Puede contener pelos radiculares que incrementen la superficie o tricomas utilizados en transpiración o defensa. Puede contener una cutícula cerosa si se encuentra en la superficie superior de las hojas, para ayudar a disminuir la transpiración.
En las plantas leñosas, la epidermis se rompe en un peridermo grueso ya que el crecimiento secundario permite que la planta crezca en circunferencia. El cámbium del corcho, que produce las células del corcho, las células del corcho (que están muertas en la madurez), y el felodermo (células del parénquima en el interior del cámbium del corcho) forman juntas el peridermo (Figura\(\PageIndex{2}\)). El peridermo funciona como la primera línea de defensa de la planta, protegiéndola de lesiones por fuego o calor, deshidratación, condiciones de congelación y/o enfermedades.

Tejido Terrestre
Muchas veces, los tejidos que no se consideran tejido dérmico o vascular se notan como tejido molido. Estas células almacenan moléculas (como el almidón), fotosintetizan (como las células mesófilas) o apoyan a la planta. El tejido del suelo a menudo se divide en tres tipos de células: Collenquima, esclerénquima y parénquima.
El collenquima (Figuras\(\PageIndex{3-4}\)) es tejido vivo de soporte que tiene células alargadas y una pared celular primaria desigualmente engrosada. Su función principal es el soporte mecánico de tallos y hojas jóvenes a través de la turgencia.
El esclerénquima es un tejido de soporte muerto que consiste en fibras largas de esclerénquima (Figura\(\PageIndex{4}\)) o células cortas, cristalinas (esclereidas; Figura\(\PageIndex{5}\)). Las fibras de esclerénquima ocurren en grupos (haces). Los esclereidos pueden ser ramificados o no y ocurren individualmente o en pequeños racimos. Cada celda tiene una pared secundaria uniformemente gruesa que es rica en lignina. Su función principal es un soporte de órganos vegetales más viejos, y también endurecer diferentes partes de las plantas (por ejemplo, hacer que el fruto no sea comestible antes de la madurez para que nadie tome el fruto antes de que las semillas estén listas para ser distribuidas). Sin esclerénquima, si no se riega una planta, las hojas caerán porque las vacuolas disminuirán de tamaño lo que disminuye la turgencia. Las fibras dentro del floema (ver abajo) a veces se consideran como un esclerénquima separado.



El parénquima (Figura\(\PageIndex{4}\)) son células esféricas alargadas con una delgada pared celular primaria. Es un componente principal de los órganos de plantas jóvenes. Las funciones básicas del parénquima son la fotosíntesis y el almacenamiento. También son importantes en la regeneración porque son totipotentes (capaces de diferenciarse en cualquier tipo celular). Las células del parénquima están muy extendidas en el cuerpo vegetal. Llenan la hoja, frecuentes en la corteza del tallo y médula y es un componente de tejidos vasculares complejos (ver más abajo).
Tejido Vascular
El tejido vascular es el sistema de plomería de la planta. Permite que el agua, los minerales y los azúcares disueltos de la fotosíntesis pasen a través de raíces, tallos, hojas y otras partes de la planta. Es primario compuesto por dos tipos de tejido conductor: xilema y floema. Las venas de las hojas son un ejemplo de tejido vascular, moviendo material a través de la planta de la misma manera que nuestros vasos sanguíneos transportan nutrientes a través de nuestro cuerpo. El xilema y el floema siempre se encuentran adyacentes entre sí (Figura\(\PageIndex{6}\)). En los tallos, el xilema y el floema forman una estructura llamada haz vascular; en las raíces, esto se denomina estela vascular o cilindro vascular.
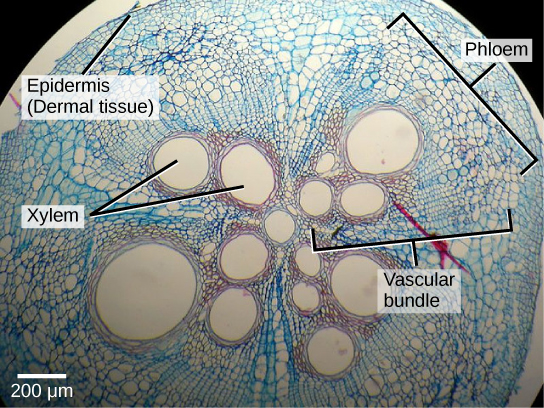
El tejido del xilema transporta agua y minerales desde las raíces a diferentes partes de la planta. Las células conductoras del xilema se denominan elementos traqueares. Las células del parénquima también se encuentran en el xilema, y a veces están presentes fibras de esclerénquima y esclereidas.
Hay dos tipos de elementos traquearios: elementos vasculares y traqueides (Figura\(\PageIndex{7}\)). Ambos tipos celulares que están muertos en la madurez y tienen paredes celulares secundarias engrosadas. Estas celdas se conectan entre sí y permiten que el agua sea transportada a través de ellas. Estructuralmente, los elementos del vaso son más anchos que las traqueidas y contienen placas de perforación entre elementos de vasos adyacentes (Figura\(\PageIndex{7-8}\)). Las amplias aberturas (hendiduras o poros) en las placas de perforación permiten que el agua fluya verticalmente entre los elementos del recipiente, formando un tubo continuo. Ambos tipos de elementos traquearios contienen hoyos, huecos en sus paredes celulares secundarias. Las celdas adyacentes tienen fosas en las mismas ubicaciones, formando pares de fosas, que permiten que el agua y los minerales fluyan entre celdas adyacentes a través de la membrana del foso (las paredes celulares primarias delgadas restantes en estas regiones; Figura\(\PageIndex{9-10}\)). Por lo tanto, el agua fluye a través de placas de perforación y pares de fosas en elementos de vasos pero solo a través de pares de fosas en traqueides Si bien el agua puede moverse más rápidamente a través de los elementos del recipiente, son más susceptibles a las burbujas de aire. Una burbuja de aire interrumpe la cohesión en la columna de agua que sube por el tubo de los elementos del recipiente impidiendo el uso de esa vía en particular. En las traqueidas, una burbuja de aire solo desmantelaría una sola traqueidea en lugar de una columna completa de elementos de vaso. Los elementos vasculares se encuentran solo en las angiospermas, pero las traqueides se encuentran tanto en angiospermas como en gimnospermas.





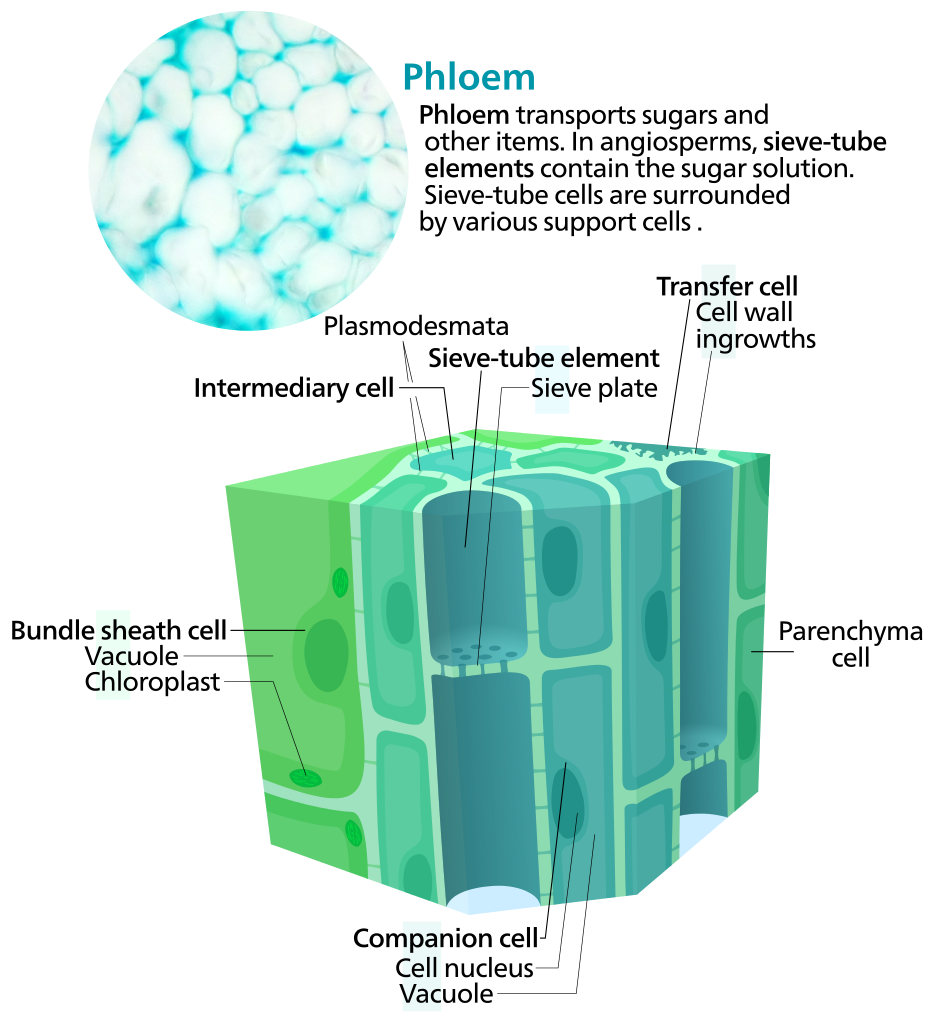
El tejido del floema transporta compuestos orgánicos como azúcares desde el sitio de la fotosíntesis hasta el resto de la planta (Figura\(\PageIndex{11-12}\)). Las células conductoras del floema se llaman elementos de tamiz. En comparación con los elementos traquearios, los elementos de tamiz solo tienen paredes celulares primarias (y, por lo tanto, paredes celulares más delgadas en general) y están vivos en la madurez; sin embargo, carecen de ciertos orgánulos, incluido un núcleo. Los elementos de tamizo-tubo son los elementos de tamiz que se encuentran solo en angiospermas mientras que las células de tamiz se encuentran solo en gimnospermas mientras que. Ambos tipos de elementos de tamiz tienen poros en sus paredes celulares (áreas de tamiz) que permiten la transferencia de materiales entre celdas adyacentes, pero estos se concentran en placas de tamiz en elementos de tubo de tamiz y se distribuyen uniformemente en celdas de tamiz. Debido a que carecen de orgánulos esenciales, los elementos de tamiz dependen de células de parénquima especializadas para apoyarlos. Las células compañeras soportan elementos de tubo de tamiz en angiospermas, y las células albuminosas soportan células de tamiz en gimnospermas. Adicionalmente, también se encuentran células de parénquima y células de esclerénquima (fibras de floema) en el floema.

La siguiente tabla resume las diferencias entre xilema y floema:
| Xilema | Floema | |
|---|---|---|
| Contiene mayormente | Células muertas | Células vivas |
| Transportes | Agua y Minerales | Azúcar |
| Dirección | Arriba | Arriba y Abajo |
| Biomasa | Big | Pequeño |
Tejido Meristemático
Los meristemas producen células que se diferencian rápidamente o se especializan, y se convierten en tejido permanente. Dichas células asumen roles específicos y pierden su capacidad de dividirse aún más. Se diferencian en tres tipos principales: tejido dérmico, vascular y tejido molido. El tejido dérmico cubre y protege la planta, y el tejido vascular transporta agua, minerales y azúcares a diferentes partes de la planta. El tejido molido sirve como sitio para la fotosíntesis, proporciona una matriz de soporte para el tejido vascular y ayuda a almacenar agua y azúcares.
Atribuciones
Comisariada y autoría de Kammy Argel y Melissa Ha utilizando las siguientes fuentes:
- 30.1 El cuerpo de la planta y 30.2 proviene de la biología 2e por OpenStax (licenciado CC-BY). Accede gratis en openstax.org.
- 5.1 Tejidos de Introducción a la Botánica de Alexey Shipunov (dominio público)