
13.5: Las reacciones dependientes de la luz
- Page ID
- 58536

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Relacionar la longitud de onda, la energía y el tipo de radiación electromagnética (y el color de la luz visible).
- Explicar cómo las plantas absorben la energía de la luz solar.
- Detallar los pasos de las interacciones dependientes de la luz.
¿Cómo se puede usar la luz para hacer alimentos? Cuando una persona enciende una lámpara, la energía eléctrica se convierte en energía luminosa. Como todas las demás formas de energía cinética, la luz puede viajar, cambiar de forma y ser aprovechada para hacer el trabajo. En el caso de la fotosíntesis, la energía luminosa se convierte en energía química, que los fotoautótrofos utilizan para construir moléculas de carbohidratos (Figura\(\PageIndex{1}\)). Sin embargo, los fotoautótrofos solo utilizan algunos componentes específicos de la luz solar.

¿Qué es la energía lumínica?
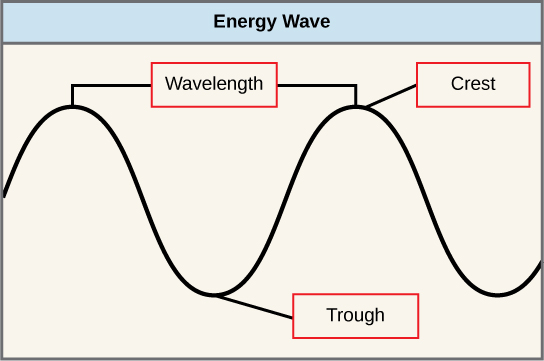
El sol emite una enorme cantidad de radiación electromagnética (energía solar). Los humanos solo pueden ver una fracción de esta energía, porción que, por lo tanto, se conoce como “luz visible”. La manera en que viaja la energía solar se describe como ondas. Los científicos pueden determinar la cantidad de energía de una onda midiendo su longitud de onda, la distancia entre puntos consecutivos de una onda. Una sola onda se mide desde dos puntos consecutivos, como de cresta a cresta o de valle a valle (Figura\(\PageIndex{2}\)). La diferencia entre longitudes de onda se relaciona con la cantidad de energía transportada por ellas.
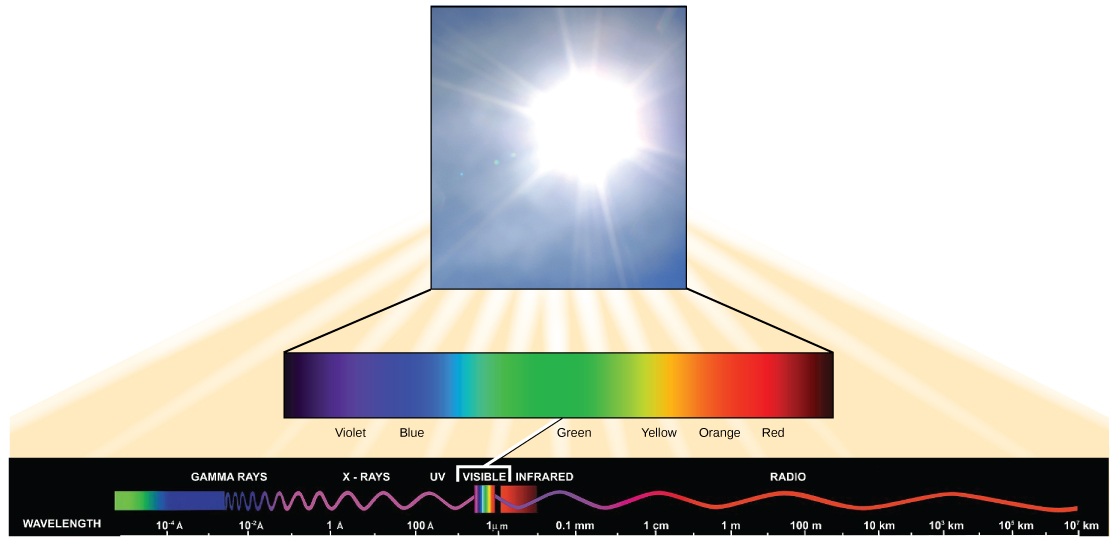
La luz visible constituye sólo uno de los muchos tipos de radiación electromagnética emitida por el sol y otras estrellas. Los científicos diferencian los diversos tipos de energía radiante del sol dentro del espectro electromagnético. El espectro electromagnético es el rango de todas las frecuencias posibles de radiación (Figura\(\PageIndex{3}\)). Varios tipos de radiación electromagnética se originan en el sol, incluidos los rayos X y los rayos ultravioleta (UV). Las ondas de mayor energía pueden penetrar los tejidos y dañar las células y el ADN, explicando por qué tanto los rayos X como los rayos UV pueden ser perjudiciales para los organismos vivos.
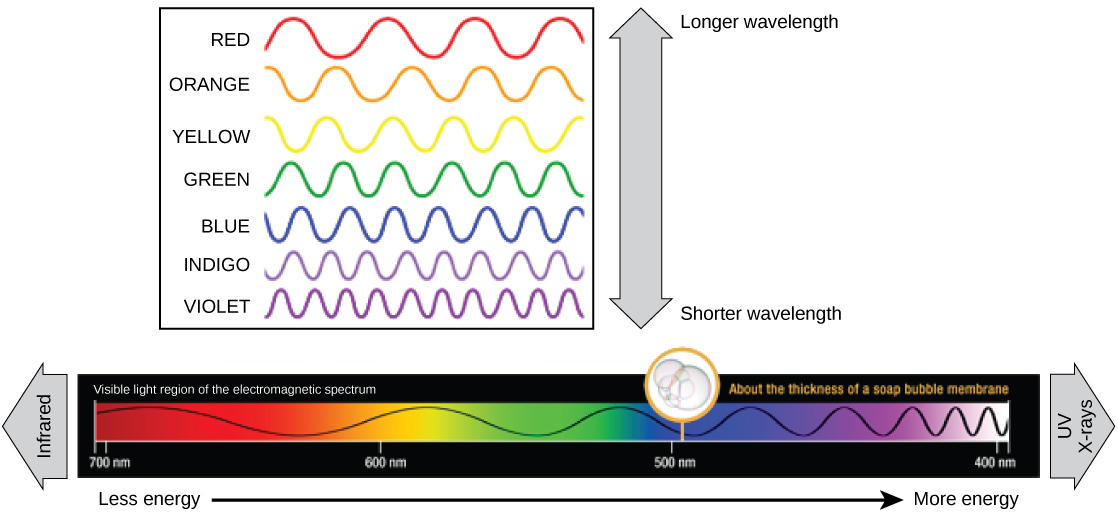
Cada tipo de radiación electromagnética viaja a una longitud de onda particular. Cuanto más larga es la longitud de onda (o cuanto más estirada aparece en el diagrama), menos energía se transporta. Las olas cortas y apretadas llevan la mayor cantidad de energía. Esto puede parecer ilógico, pero piénsalo en términos de un trozo de mover una cuerda pesada. Se necesita poco esfuerzo de una persona para mover una cuerda en olas largas y anchas. Para hacer que una cuerda se mueva en olas cortas y apretadas, una persona necesitaría aplicar significativamente más energía.
Absorción de Luz
La energía lumínica inicia el proceso de fotosíntesis cuando los pigmentos absorben la luz. Los pigmentos orgánicos, ya sea en la retina humana o en el tilacoide del cloroplasto, tienen un estrecho rango de niveles de energía que pueden absorber. Los niveles de energía inferiores a los representados por la luz roja son insuficientes para excitar electrones en los pigmentos retinianos. Los niveles de energía superiores a los de la luz azul desgarrarán físicamente estas moléculas, llamadas blanqueamiento. Por lo que los pigmentos retinianos solo pueden “ver” (absorber) la luz de 700 nm a 400 nm, que por lo tanto se llama luz visible. Por las mismas razones, las moléculas de pigmento vegetal absorben solo la luz en el rango de longitud de onda de 700 nm a 400 nm; los fisiólogos de plantas se refieren a este rango para las plantas como radiación fotosintéticamente activa.
La luz visible vista por los humanos como luz blanca existe realmente en un arco iris de colores. Ciertos objetos, como un prisma o una gota de agua, dispersan la luz blanca para revelar los colores al ojo humano. La porción de luz visible del espectro electromagnético muestra el arco iris de colores, con violeta y azul teniendo longitudes de onda más cortas, y por lo tanto mayor energía. En el otro extremo del espectro hacia el rojo, las longitudes de onda son más largas y tienen menor energía (Figura\(\PageIndex{4}\)).
Comprensión de los pigmentos
Existen diferentes tipos de pigmentos, y cada uno ha evolucionado para absorber solo ciertas longitudes de onda (colores) de luz visible. Los pigmentos reflejan o transmiten las longitudes de onda que no pueden absorber, haciéndolas aparecer en el color correspondiente.
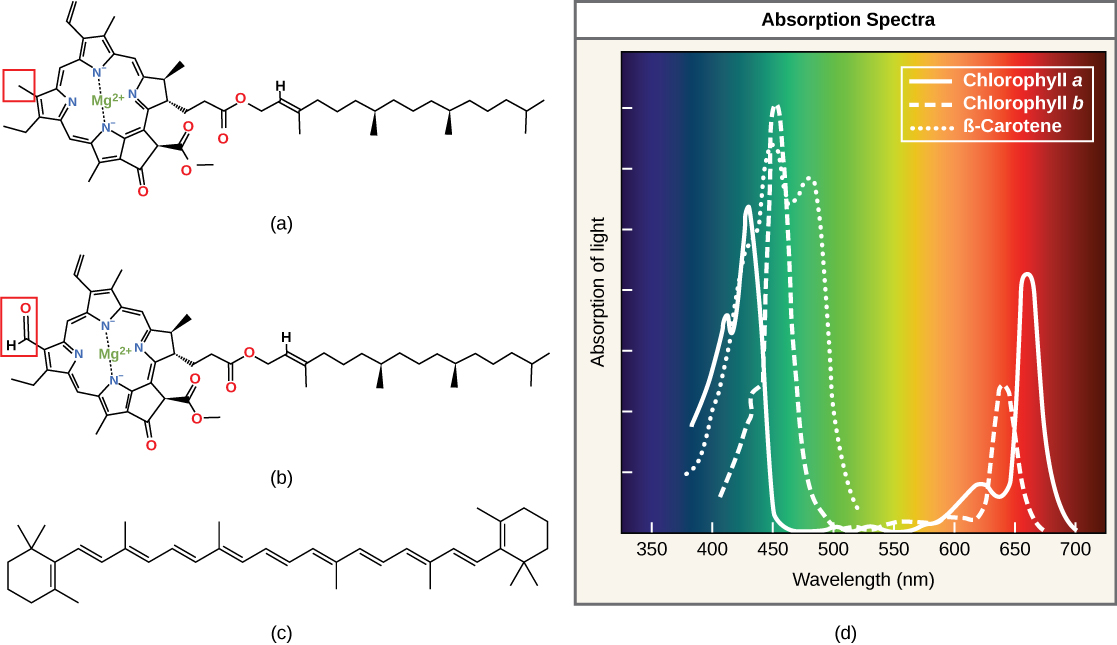
Las clorofilas y carotenoides son las dos clases principales de pigmentos fotosintéticos que se encuentran en plantas y algas; cada clase tiene múltiples tipos de moléculas de pigmento. Hay cinco clorofilas principales: a, b, c y d y una molécula relacionada que se encuentra en procariotas llamada bacterioclorofila. La clorofila a y la clorofila b se encuentran en cloroplastos vegetales y serán el foco de la siguiente discusión.
Con docenas de formas diferentes, los carotenoides son un grupo mucho más grande de pigmentos. Los carotenoides que se encuentran en la fruta, como el rojo del tomate (licopeno), el amarillo de las semillas de maíz (zeaxantina) o la naranja de una cáscara de naranja (β-caroteno), se utilizan como anuncios para atraer dispersores de semillas. En la fotosíntesis, los carotenoides funcionan como pigmentos fotosintéticos que son moléculas muy eficientes para la eliminación del exceso de energía. Cuando una hoja se expone a pleno sol, se requieren las reacciones dependientes de la luz para procesar una enorme cantidad de energía; si esa energía no se maneja adecuadamente, puede causar daños significativos. Por lo tanto, muchos carotenoides residen en la membrana tilacoidea, absorben el exceso de energía y disipan esa energía de manera segura como calor.
Cada tipo de pigmento se puede identificar por el patrón específico de longitudes de onda que absorbe de la luz visible, que es el espectro de absorción. La gráfica de la Figura\(\PageIndex{5}\) muestra los espectros de absorción de clorofila a, clorofila b y un tipo de pigmento carotenoide llamado β-caroteno (que absorbe la luz azul y verde). Observe cómo cada pigmento tiene un conjunto distinto de picos y valles, revelando un patrón de absorción altamente específico. La clorofila a absorbe longitudes de onda de cualquier extremo del espectro visible (azul y rojo), pero no verde. Debido a que el verde se refleja o transmite, la clorofila aparece verde. Los carotenoides se absorben en la región azul de longitud de onda corta y reflejan las longitudes de onda más largas de amarillo, rojo y naranja.
Muchos organismos fotosintéticos tienen una mezcla de pigmentos; utilizándolos, el organismo puede absorber energía de un rango más amplio de longitudes de onda. No todos los organismos fotosintéticos tienen pleno acceso a la luz solar. Algunos organismos crecen bajo el agua donde la intensidad y calidad de la luz disminuyen y cambian con la profundidad (Figura\(\PageIndex{6}\)). Otros organismos crecen en competencia por la luz. Las plantas en el suelo de la selva tropical deben ser capaces de absorber cualquier poco de luz que pase, ya que los árboles más altos absorben la mayor parte de la luz solar y dispersan la radiación solar restante (Figura\(\PageIndex{7}\)).


Al estudiar un organismo fotosintético, los científicos pueden determinar los tipos de pigmentos presentes generando espectros de absorción. Un instrumento llamado espectrofotómetro puede diferenciar qué longitudes de onda de luz puede absorber una sustancia. Los espectrofotómetros miden la luz transmitida y computan a partir de ella la absorción. Al extraer pigmentos de las hojas y colocar estas muestras en un espectrofotómetro, los científicos pueden identificar qué longitudes de onda de luz puede absorber un organismo. Los métodos adicionales para la identificación de pigmentos vegetales incluyen diversos tipos de cromatografía que separan los pigmentos por sus afinidades relativas a las fases sólida y móvil.
Cómo funcionan las reacciones dependientes de la luz
La función general de las reacciones dependientes de la luz es convertir la energía solar en energía química en forma de NADPH y ATP. Esta energía química apoya las reacciones independientes de la luz y alimenta el ensamblaje de moléculas de azúcar. En las reacciones dependientes de la luz, los complejos proteicos y las moléculas pigmentarias trabajan juntos para producir NADPH y ATP (Figura\(\PageIndex{8}\) y Video\(\PageIndex{1}\)).

Video\(\PageIndex{1}\): Aquí hay una animación de las reacciones dependientes de la luz.
Los fotosistemas absorben la energía de la luz
El paso real que convierte la energía luminosa en energía química tiene lugar en un complejo multiproteico llamado fotosistema, dos tipos de los cuales se encuentran incrustados en la membrana tilacoidea, el fotosistema II (PSII) y el fotosistema I ( PSI) (Figura\(\PageIndex{9}\)). Los dos complejos difieren en base a lo que oxidan (es decir, la fuente del suministro de electrones de baja energía) y lo que reducen (el lugar al que entregan sus electrones energizados).
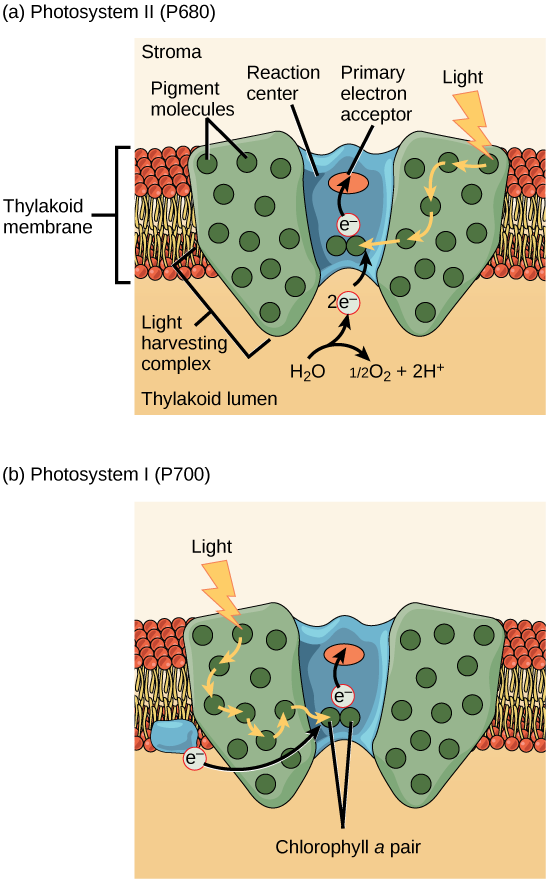
Ambos fotosistemas tienen la misma estructura básica; una serie de proteínas de antena a las que se unen las moléculas de clorofila rodean el centro de reacción, donde tiene lugar la fotoquímica. Cada fotosistema es atendido por un complejo de antenas, que pasa la energía de la luz solar al centro de reacción. El complejo de antenas consiste en múltiples proteínas de antena que contienen una mezcla de 300 a 400 moléculas de clorofila a y b, así como otros pigmentos como los carotenoides. (Técnicamente, los fotosistemas consisten en el centro de reacción y el complejo de antenas, y un fotosítem más complejos de recolección de luz comprende un complejo de fotosistema, pero en la Figura\(\PageIndex{9}\), el complejo de captación de luz se etiqueta como el complejo de antenas). La absorción de un solo fotón, o cantidad distinta o “paquete” de luz, por cualquiera de las clorofilas empuja esa molécula a un estado excitado. En definitiva, la energía de la luz ahora ha sido capturada por moléculas biológicas pero aún no se almacena en ninguna forma útil. La energía se transfiere de la clorofila a la clorofila hasta que finalmente (después de aproximadamente una millonésima de segundo), se entrega al centro de reacción. Hasta este punto, sólo se ha transferido energía entre moléculas, no electrones.
El centro de reacción contiene un par de moléculas de clorofila a con una propiedad especial. Esas dos clorofilas pueden sufrir oxidación tras la excitación; en realidad pueden renunciar a un electrón. Es en este paso en el centro de reacción, este paso en la fotosíntesis, donde la energía luminosa se convierte en un electrón excitado.
Los electrones se mueven hacia abajo en la cadena de transporte de electrones, proporcionando energía para bombear protones al lumen tilacoide
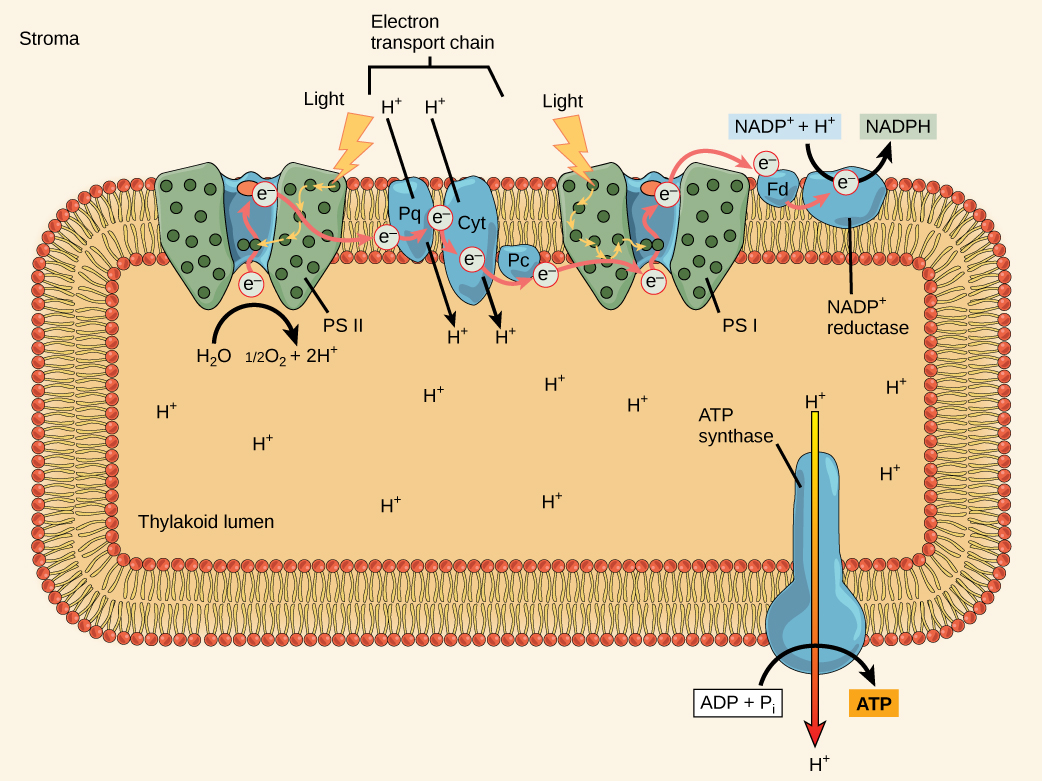
El centro de reacción del PSII (llamado P680) entrega sus electrones de alta energía, uno a la vez, al aceptor de electrones primario, y a través de la cadena de transporte de electrones (Pq al complejo del citocromo b6f a plastocianina) a PSI (Figura\(\PageIndex{10}\)). El complejo citocromo b6f, una enzima compuesta por dos complejos proteicos, transfiere los electrones de la molécula portadora plastoquinona (Pq) a la proteína plastocianina (Pc), facilitando finalmente la transferencia de electrones de PSII a PSI. Durante este proceso, el citocromo b6f bombea protones desde el estroma hacia el lumen tilacoide utilizando la energía de los electrones que se mueven hacia abajo de la cadena de transporte de electrones.
Fotofosforilación
La síntesis de ATP en las reacciones dependientes de la luz se llama fotofosforilación. La acumulación de protones dentro de la luz tilacoidea crea un gradiente de concentración. El movimiento pasivo (difusión facilitada) de protones de alta concentración (en la luz tilacoidea) a baja concentración (en el estroma) se aprovecha para crear ATP. Los protones acumulan energía a altas concentraciones porque todos tienen la misma carga eléctrica, repeliéndose entre sí.
Para liberar esta energía, los protones correrán por cualquier abertura, similar al chorro de agua a través de un agujero en una presa. En el tilacoide, esa apertura es un paso a través de un canal proteico especializado llamado ATP sintasa. La energía liberada por la corriente de protones permite a la ATP sintasa unir un tercer grupo fosfato al ADP, que forma una molécula de ATP (Figuras\(\PageIndex{10-11}\)). El flujo de protones a través de la ATP sintasa se llama quimiósmosis porque los iones se mueven de un área de alta a una zona de baja concentración a través de una estructura semipermeable.

Fotólisis del agua
El electrón faltante de P680 es reemplazado por la extracción de un electrón de baja energía del agua; así, el agua se divide (fotólisis de agua) y el PSII se vuelve a reducir. La división de una molécula H 2 O libera dos electrones, dos protones y un átomo de oxígeno (Figuras\(\PageIndex{10-11}\)). Esos protones, además de los transportados por el citrocromo b6f, se acumulan en la luz tilacoidea y alimentan la fotofosforilación. Se requiere dividir dos moléculas para formar una molécula de gas O 2 diatómico. Alrededor del 10 por ciento del oxígeno es utilizado por las mitocondrias en la hoja para apoyar la respiración celular aeróbica, el proceso de descomponer la glucosa para generar ATP. El resto escapa a la atmósfera donde es utilizado por organismos aeróbicos para apoyar la respiración.
Reducción de NADP +
Debido a que los electrones han perdido energía antes de su llegada a PSI, deben ser re-energizados por PSI, de ahí que otro fotón sea absorbido por la antena PSI. Esa energía se retransmite al centro de reacción PSI (llamado P700). P700 se oxida y envía electrones de alta energía al portador de electrones NADP + para formar NADPH. Así, PSII captura la energía para crear gradientes de protones para hacer ATP, y PSI captura la energía para reducir NADP + en NADPH. Los dos fotosistemas trabajan en concierto, en parte, para garantizar que se generen las proporciones correctas de NADPH y ATP necesarias para las reacciones independientes de la luz. Existen otros mecanismos para afinar esa proporción para que coincida exactamente con las necesidades de energía constantemente cambiantes del cloroplasto.
Atribución
Curada y autoría de Melissa Ha usando 8.2 Las reacciones dependientes de la luz de la fotosíntesis de la biología 2e por OpenStax (con licencia CC-BY). Accede gratis en openstax.org.