
13.7: Fotorespiración y Vías Fotosintéticas
- Page ID
- 58506

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Definir la fotorespiración.
- Explique cómo las plantas C3, C4 y CAM reducen la fotorespiración.
- Esbozar la vía C 4 y comparar su uso por plantas C 4 y plantas CAM.
Diferentes especies de plantas tienen adaptaciones que les permiten hacer diferentes variaciones de las reacciones independientes de la luz. A estas se les llama vías fotosintéticas. Las plantas se clasifican como C 3, C 4 o CAM dependiendo de su uso de estas vías, pero tenga en cuenta que algunas plantas pueden cambiar vías fotosintéticas dependiendo de las condiciones ambientales. El proceso para las reacciones independientes de la luz descrito en la sección anterior fue la vía C 3: el compuesto formado durante la fijación (3-PGA) tiene tres átomos de carbono. Antes de discutir los detalles de la vía C 4, es importante comprender las circunstancias que llevaron a estas adaptaciones.
Fotorespiración
Como su nombre indica, la ribulosa-1,5-bisfosfato carboxilasa/oxigenasa (RuBisCO) cataliza dos reacciones diferentes. El primero es agregar CO 2 a la ribulosa-1,5- bisfosfato (RuBP), la actividad de la carboxilasa. El segundo es agregar O 2 a RuBP, la actividad oxigenasa.
La actividad oxigenasa de RuBisCO forma la molécula de tres carbonos 3-fosfoglicerato (3-PGA), al igual que en las reacciones independientes de la luz, y la molécula de dos carbonos glicolato. El glicolato ingresa a los peroxisomas, donde usa O 2 para formar intermedios que ingresan a las mitocondrias donde se descomponen en CO 2. Por lo que este proceso usa O 2 y libera CO 2 como lo hace la respiración celular aeróbica, razón por la cual se llama fotorespiración. Deshace el trabajo de la fotosíntesis, que es construir azúcares.
Qué acción de RubiSco predomina depende de las concentraciones relativas de O 2 y CO 2 con CO 2 alto, O 2 bajo favoreciendo la acción de carboxilasa y O 2 alto, CO bajo 2 favoreciendo la acción oxigenasa. Las reacciones lumínicas de la fotosíntesis liberan oxígeno, y más oxígeno se disuelve en el citosol de la célula a temperaturas más altas. Por lo tanto, las altas intensidades de luz y las altas temperaturas (por encima de ~ 30°C) favorecen la segunda reacción y dan como resultado la fotorespiración.
C 3 Plantas
Una solución a la fotorespiración es que las plantas abran sus estomas para liberar O 2 y obtener CO 2. Sin embargo, si las condiciones son calientes o secas, esto provocará demasiada pérdida de agua (transpiración). Por esta razón, las plantas C 3, que solo realizan la vía C 3 y no utilizan la vía C 4 para prevenir la fotorespiración (ver abajo), funcionan mejor en áreas frescas y húmedas. El arroz y las papas son ejemplos de plantas C 3.
C 4 Plantas
Muchas angiospermas han desarrollado adaptaciones que minimizan las pérdidas a la fotorespiración. Todos ellos utilizan un método suplementario de captación de CO 2 que inicialmente forma una molécula de cuatro carbonos en comparación con las dos moléculas de tres carbonos que se forman inicialmente en la vía C 3. De ahí que estas plantas se llamen plantas C 4. Tenga en cuenta que las plantas C 4 eventualmente conducirán las reacciones independientes de la luz (vía C 3), pero primero forman una molécula de cuatro carbonos.
Las plantas C 4 presentan cambios estructurales en su anatomía foliar de manera que sintetizar el azúcar de cuatro carbonos (la vía C 4) y reanudar las reacciones independientes de la luz (vías C 3) se separan en diferentes partes de la hoja con RubisCO secuestró donde el nivel de CO 2 es alto y el nivel de O 2 bajo. Después de ingresar por los estomas, el CO 2 se difunde en una célula mesófila (Figura\(\PageIndex{1}\)). Al estar cerca de la superficie foliar, estas células están expuestas a altos niveles de O 2, pero no tienen RubiSco por lo que no pueden iniciar la fotorespiración (ni las reacciones independientes de la luz).
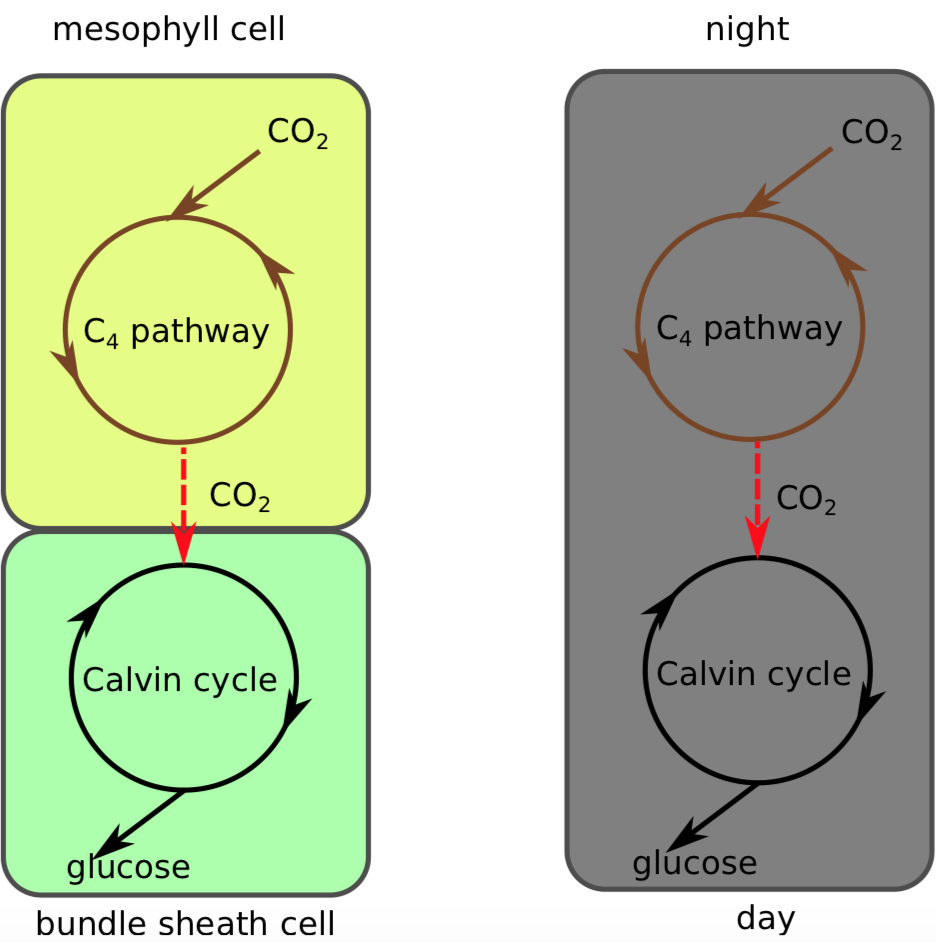
En cambio, el CO 2 se inserta en un compuesto de tres carbonos llamado ácido fosfoenolpirúvico (PEP) formando el compuesto de cuatro carbonos ácido oxaloacético. El ácido oxaloacético se convierte en ácido málico o ácido aspártico (ambos tienen 4 carbonos), el cual se transporta a través de plasmodesmas a una célula de vaina de haz. Las células de vaina de haz están profundas en la hoja, por lo que el oxígeno atmosférico no puede difundirse fácilmente hacia ellas (Figura\(\PageIndex{2}\)). Adicionalmente, a menudo tienen tilacoides con complejos reducidos de fotosistema II (el que produce O 2). Ambas características mantienen bajos los niveles de oxígeno en las células de vaina de haz, que es donde el compuesto de cuatro carbonos se descompone en dióxido de carbono, que ingresa a las reacciones independientes de la luz (vía C 3) para formar azúcares y ácido pirúvico, que se transporta de regreso a una célula mesófila donde se encuentra convertido de nuevo en PEP.

Estas plantas C 4 están bien adaptadas (y es probable que se encuentren en) hábitats con altas temperaturas diurnas y luz solar intensa. Debido a que utilizan la vía C 4 para prevenir la fotorespiración, no tienen que abrir sus estomas en la misma medida que las plantas C 3 y así pueden conservar el agua. Algunos ejemplos de pasto cangrejo, maíz (maíz), caña de azúcar y sorgo. Aunque comprenden solo ~ 3% de las angiospermas por especie, las plantas C 4 son responsables de ~ 25% de toda la fotosíntesis en tierra.
Plantas CAM
CAM significa c rassulacean a cid m etabolism porque se estudió por primera vez en miembros de la familia de plantas Crassulaceae. Las plantas CAM también realizan la vía C 4. Sin embargo, en lugar de segregar las vías C 4 y C 3 en diferentes partes de la hoja, las plantas CAM las separan en el tiempo (Tabla\(\PageIndex{1}\)). Como resultado, las plantas CAM no necesitan abrir sus estomas durante el día para reducir la fotorespiración porque ya han formado una molécula de cuatro carbonos por la noche que se puede descomponer para liberar dióxido de carbono durante el día.
| Noche | Mañana |
|---|---|
|
|
Por lo tanto, las plantas CAM prosperan en condiciones de altas temperaturas diurnas, luz solar intensa y baja humedad del suelo. Algunos ejemplos de plantas CAM incluyen cactus (Figura\(\PageIndex{3}\)), piñas, todas las bromelias epífitas, sedums y la “planta de hielo” que invade la costa de California.

Atribución
Curada y autoría de Melissa Ha usando 16.2E Photorespiration y C4 Plants from Biology por John W. Kimball (licenciado CC-BY)