
17.1.3: Teoría de Cohesión-Tensión
- Page ID
- 58364

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Explicar cómo el agua se mueve hacia arriba a través de una planta según la teoría de cohesión-tensión.
- Proporcionar evidencia experimental para la teoría de la cohesión-tensión.
En 1895, los fisiólogos irlandeses H. H. Dixon y J. Joly propusieron que el agua es levantada por la planta por tensión (presión negativa) desde arriba. Como hemos visto, el agua se pierde continuamente de las hojas por la transpiración. Dixon y Joly creían que la pérdida de agua en las hojas ejerce un tirón sobre el agua en los conductos del xilema y atrae más agua hacia la hoja. Pero incluso la mejor bomba de vacío puede tirar el agua hasta una altura de solo 10.4 m (34 pies) más o menos. Esto se debe a que una columna de agua que alta ejerce una presión de 1.03 MPa apenas compensada por la presión de la atmósfera. ¿Cómo se puede atraer el agua hasta la cima de una secuoya, la más alta tiene 113 m (370 pies) de altura? Teniendo en cuenta todos los factores, probablemente se necesite un tirón de al menos ~1.9 MPa.
La respuesta al dilema radica en la cohesión de las moléculas de agua; esa es la propiedad de que las moléculas de agua se aferren a cada una a través de los enlaces de hidrógeno que forman (Figura\(\PageIndex{1}\)). Cuando el agua ultrapura se limita a tubos de muy pequeño calibre, la fuerza de cohesión entre las moléculas de agua imparte una gran fuerza a la columna de agua. Se ha reportado que se necesitan tensiones de hasta 21 MPa para romper la columna, alrededor del valor necesario para romper alambres de acero del mismo diámetro. En cierto sentido, la cohesión de las moléculas de agua les da las propiedades físicas de los alambres sólidos.

Según la teoría de la cohesión-tensión, la transpiración es el principal impulsor del movimiento del agua en el xilema. Crea presión negativa (tensión) equivalente a —2 MPa en la superficie foliar. El agua de las raíces es finalmente levantada por esta tensión. El potencial hídrico negativo atrae agua del suelo hacia los pelos de la raíz, luego hacia el xilema radicular. La cohesión y la adhesión arrastran el agua hasta el xilema. La transpiración extrae agua de la hoja a través del estoma. El potencial hídrico se vuelve cada vez más negativo desde las células radiculares hasta el tallo hasta las hojas más altas, y finalmente a la atmósfera (Figura\(\PageIndex{2}\)).
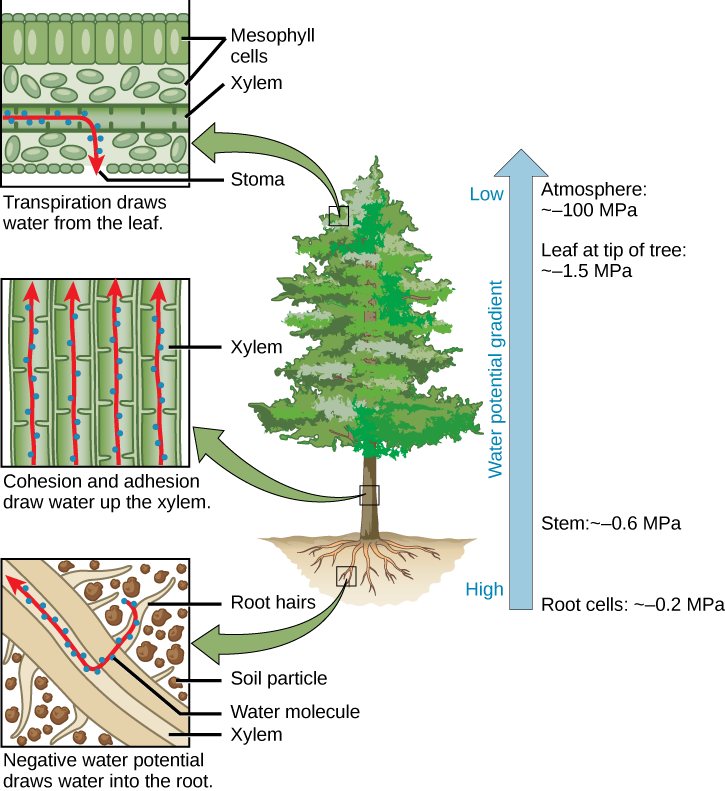
El potencial hídrico en la superficie foliar varía mucho dependiendo del déficit de presión de vapor, que puede ser insignificante a alta humedad relativa (HR) y sustancial a baja HR. Por la noche, cuando los estomas suelen cerrarse y la transpiración se detiene, el agua se mantiene en el tallo y la hoja por la adhesión del agua a las paredes celulares de los vasos del xilema y las traqueidas, y la cohesión de las moléculas de agua entre sí.
El mecanismo de la teoría de la cohesión-tensión se basa en fuerzas puramente físicas debido a que los vasos del xilema y las traqueidas no viven en la madurez. La evaporación del agua en los espacios intercelulares de aire crea una mayor tensión sobre el agua en las células mesófilas, aumentando así la atracción sobre el agua en los vasos del xilema. Los vasos del xilema y las traqueides están estructuralmente adaptados para hacer frente a grandes cambios de presión. Los anillos en los recipientes mantienen su forma tubular, al igual que los anillos en la manguera de una aspiradora mantienen la manguera abierta mientras está bajo presión. Pequeñas perforaciones entre los elementos del vaso reducen el número y tamaño de las burbujas de gas que se pueden formar a través de un proceso llamado cavitación. La formación de burbujas de gas en el xilema interrumpe la corriente continua de agua desde la base hasta la parte superior de la planta, provocando una ruptura denominada embolia en el flujo de savia de xilema. Cuanto más alto es el árbol, mayores son las fuerzas de tensión necesarias para tirar del agua, y más eventos de cavitación. En árboles más grandes, las embolias resultantes pueden taponar los vasos del xilema, haciéndolos no funcionales.
La fuerza de tracción debida a la transpiración es tan poderosa que permite que algunos árboles y arbustos vivan en el agua de mar. El agua de mar es marcadamente hipertónica al citoplasma en las raíces del manglar rojo (Rhizophora mangle), y podríamos esperar que el agua salga de las células resultando en una pérdida de turgencia y marchitamiento. Sin embargo, las tensiones notablemente altas en el xilema (~3 a 5 MPa) pueden atraer agua a la planta contra este gradiente osmótico. Los manglares literalmente desalientan el agua de mar para satisfacer sus necesidades.
La evidencia experimental apoya la teoría de la cohesión-tensión. Hace más de un siglo, un botánico alemán que aserró un roble de 21 m (70 pies) y colocó la base del tronco en un barril de solución de ácido pícrico. La solución se elaboró por el tronco, matando a los tejidos cercanos a medida que avanzaba. Si las raíces fueran la fuerza impulsora, el movimiento ascendente del agua se habría detenido tan pronto como el ácido mató a las raíces. Sin embargo, la solución llegó a la cima del árbol. Cuando el ácido llegó a las hojas y las mató, cesó el movimiento del agua, demostrando que la transpiración en las hojas estaba causando al agua el movimiento ascendente del agua.
Según la teoría de cohesión-tensión, el agua en el xilema está bajo tensión debido a la transpiración. Consistente con esta predicción, el diámetro de los pinos Monterey disminuye durante el día, cuando las tasas de transpiración son mayores (Figura\(\PageIndex{3}\)). Debido a que la columna de agua está bajo tensión, las paredes del xilema son arrastradas por adherencia.
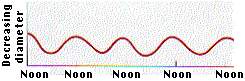
Los árboles más altos
Al hilar ramas en una centrífuga, se ha demostrado que el agua en el xilema evita la cavitación a presiones negativas superiores a ~1.6 MPa. Y el hecho de que las secuoyas gigantes (Sequoia sempervirens, Figura\(\PageIndex{4}\)) puedan levantar con éxito el agua 109 m (358 pies), lo que requeriría una tensión de ~1.9 MPa, lo que indica que se evita la cavitación incluso a ese valor. Sin embargo, tales alturas pueden estar acercándose al límite para el transporte de xilema. Las mediciones cercanas a la cima de uno de los árboles de secuoyas gigantes vivientes más altos, 112.7 m (~370 pies), muestran que las altas tensiones necesarias para transportar el agua han dado como resultado estomas más pequeños, causando menores concentraciones de CO 2 en las agujas, menor fotosíntesis y crecimiento reducido ( células más pequeñas y agujas mucho más pequeñas; Koch et al. 2004). Los límites en el transporte por agua limitan así la altura máxima que los árboles pueden alcanzar. El árbol vivo más alto es una secoya gigante de 115.9-m, y el árbol más alto jamás medido, un abeto Douglas, fue 125.9 m.

Referencia: Koch, G., Sillett, S., Jennings, G. et al. Los límites a la altura del árbol. Naturaleza 428, 851—854 (2004). https://doi.org/10.1038/nature02417
Lectura suplementaria
Woodward, I. Plantas altas. Naturaleza 428, 807—808 (2004). https://doi.org/10.1038/428807a
Atribuciones
Comisariada y autoría de Melissa Ha utilizando las siguientes fuentes:
- 30.5 Transporte de Agua y Solutos en Plantas de Biología 2e por OpenStax (licenciado CC-BY). Accede gratis en openstax.org.
- 16.2A Xilema de Biología por John. W. Kimball (licencia CC-BY)