
4.4: Ecología Comunitaria
- Page ID
- 52713

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Por lo general, las poblaciones no viven aisladas de otras especies. Las poblaciones que interactúan dentro de un hábitat dado forman una comunidad. El número de especies que ocupan el mismo hábitat y su abundancia relativa se conoce como la diversidad de la comunidad. Las áreas con baja diversidad de especies, como los glaciares de la Antártida, aún contienen una amplia variedad de organismos vivos, mientras que la diversidad de las selvas tropicales es tan grande que no se puede evaluar con precisión. Los científicos estudian la ecología a nivel comunitario para comprender cómo las especies interactúan entre sí y compiten por los mismos recursos.
Depredación y Herbivoría
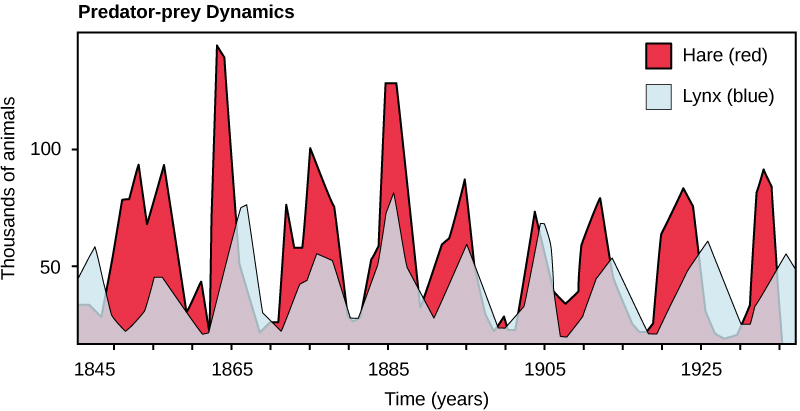
Quizás el ejemplo clásico de interacción entre especies es la relación depredador-presa. La definición más estrecha de depredación describe individuos de una población que matan y luego consumen a los individuos de otra población. Los tamaños de población de depredadores y presas en una comunidad no son constantes a lo largo del tiempo, y pueden variar en ciclos que parecen estar relacionados. El ejemplo más citado de dinámica poblacional depredador-presa se observa en el ciclo del lince (depredador) y la liebre raqueta (presa), utilizando 100 años de datos de captura de América del Norte (Figura\(\PageIndex{1}\)). Este ciclo de tamaños de población de depredadores y presas tiene un período de aproximadamente diez años, con la población depredadora rezagada de uno a dos años por detrás de la población de presas. Una explicación aparente de este patrón es que a medida que aumentan los números de liebre, hay más alimentos disponibles para el lince, lo que permite que la población de lince aumente también. Cuando la población de lince crece a un nivel umbral, sin embargo, matan a tantas liebres que el número de liebre comienza a disminuir, seguido de una disminución en la población de lince debido a la escasez de alimentos. Cuando la población de lince es baja, el tamaño de la población de liebre comienza a aumentar debido, en parte, a la baja presión de depredación, iniciando de nuevo el ciclo.
Mecanismos de Defensa contra la Depredación y Herbivoría
La depredación y la evitación de depredadores están fuertemente influenciadas por la selección natural. Cualquier carácter heredable que permita a un individuo de una población de presas evadir mejor a sus depredadores se representará en mayor número en generaciones posteriores. De igual manera, los rasgos que permiten a un depredador localizar y capturar de manera más eficiente a su presa conducirán a un mayor número de crías y a un aumento en la comunalidad del rasgo dentro de la población. Tales relaciones ecológicas entre poblaciones específicas conducen a adaptaciones que son impulsadas por respuestas evolutivas recíprocas en esas poblaciones. Las especies han desarrollado numerosos mecanismos para escapar de la depredación (incluyendo la herbivoría, el consumo de plantas como alimento). Las defensas pueden ser mecánicas, químicas, físicas o conductuales.

Las defensas mecánicas, como la presencia de armaduras en animales o espinas en plantas, desalientan la depredación y la herbivoría al desalentar el contacto físico (Figura\(\PageIndex{2}\) a). Muchos animales producen u obtienen defensas químicas de las plantas y las almacenan para evitar la depredación. Muchas especies vegetales producen compuestos secundarios de plantas que no sirven para la planta excepto que son tóxicos para los animales y desalientan el consumo. Por ejemplo, la dedalera produce varios compuestos, entre ellos la digital, que son extremadamente tóxicos cuando se comen (Figura\(\PageIndex{2}\) b). (Los científicos biomédicos han reutilizado el químico producido por la dedalera como un medicamento para el corazón, lo que ha salvado vidas durante muchas décadas).
Muchas especies utilizan la forma y coloración de su cuerpo para evitar ser detectadas por depredadores. El bastón tropical es un insecto con la coloración y forma corporal de una ramita, lo que hace que sea muy difícil de ver cuando está estacionario sobre un fondo de ramitas reales (Figura\(\PageIndex{3}\) a). En otro ejemplo, el camaleón puede cambiar su color para que coincida con su entorno (Figura\(\PageIndex{3}\) b).

Algunas especies utilizan la coloración como una forma de advertir a los depredadores de que son de mal gusto o venenosos. Por ejemplo, la oruga mariposa monarca secuestra venenos de su alimento (plantas y algodoncillo) para hacerse venenosa o desagradable para los depredadores potenciales. La oruga es de color amarillo brillante y negro para anunciar su toxicidad. La oruga también es capaz de pasar las toxinas secuestradas al monarca adulto, que también es dramáticamente coloreado de negro y rojo como advertencia para posibles depredadores. Los sapos de vientre de fuego producen toxinas que los hacen de mal gusto para sus depredadores potenciales (Figura\(\PageIndex{4}\)). Tienen una coloración roja o naranja brillante en sus vientres, que muestran a un depredador potencial para anunciar su naturaleza venenosa y desalentar un ataque. La coloración de advertencia solo funciona si un depredador usa la vista para localizar presas y puede aprender—un depredador ingenuo debe experimentar las consecuencias negativas de comer una antes de evitar a otros individuos de color similar.

Si bien algunos depredadores aprenden a evitar comer ciertas presas potenciales debido a su coloración, otras especies han desarrollado mecanismos para imitar esta coloración para evitar ser comidos, a pesar de que ellos mismos pueden no ser desagradables de comer o contener químicos tóxicos. En algunos casos de mimetismo, una especie inofensiva imita la coloración de advertencia de una especie dañina. Asumiendo que comparten los mismos depredadores, esta coloración protege entonces a los inofensivos. Muchas especies de insectos imitan la coloración de las avispas, que son picantes, insectos venenosos, desalentando así la depredación (Figura\(\PageIndex{5}\)).

En otros casos de mimetismo, múltiples especies comparten la misma coloración de advertencia, pero todas ellas en realidad tienen defensas. La comunalidad de la señal mejora el cumplimiento de todos los depredadores potenciales. La figura\(\PageIndex{6}\) muestra una variedad de mariposas de mal sabor con coloración similar.

Principio de exclusión competitiva
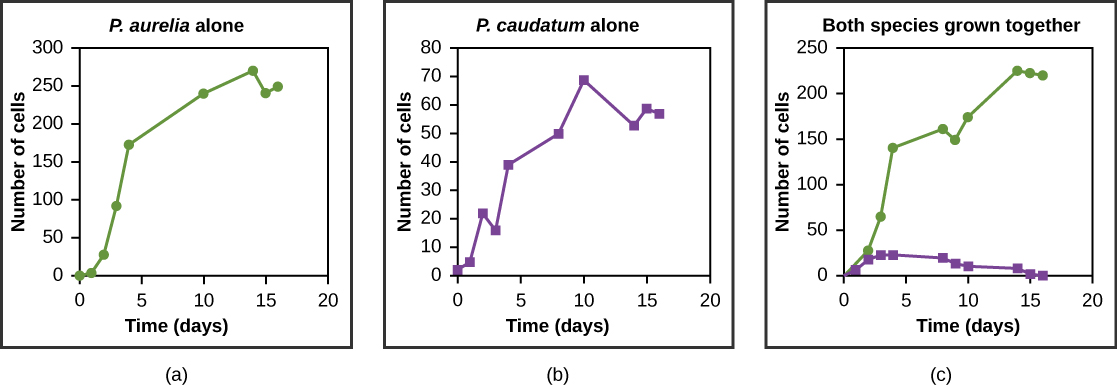
Los recursos suelen ser limitados dentro de un hábitat y múltiples especies pueden competir para obtenerlos. Los ecologistas han llegado a entender que todas las especies tienen un nicho ecológico: el conjunto único de recursos que utiliza una especie, que incluye sus interacciones con otras especies. El principio de exclusión competitiva establece que dos especies no pueden ocupar exactamente el mismo nicho en un hábitat. Es decir, diferentes especies no pueden coexistir en una comunidad si están compitiendo por todos los mismos recursos. Es importante señalar que la competencia es mala para ambos competidores porque desperdicia energía. El principio de exclusión competitiva funciona porque si hay competencia entre dos especies por los mismos recursos, entonces la selección natural favorecerá rasgos que disminuyan la dependencia del recurso compartido, reduciendo así la competencia. Si alguna de las especies es incapaz de evolucionar para reducir la competencia, entonces la especie que explote más eficientemente el recurso conducirá a la otra especie a la extinción. Un ejemplo experimental de este principio se muestra en la Figura\(\PageIndex{7}\) con dos especies protozoarias: Paramecium aurelia y Paramecium caudatum. Cuando se cultivan individualmente en el laboratorio, ambos prosperan. Pero cuando se colocan juntos en el mismo tubo de ensayo (hábitat), P. aurelia supera a P. caudatum por los alimentos, lo que lleva a la eventual extinción de este último.
Simbiosis
Las relaciones simbióticas son interacciones cercanas a largo plazo entre individuos de diferentes especies. Las simbiosis pueden ser comensales, en las que una especie se beneficia mientras que la otra no es perjudicada ni beneficiada; mutualistas, en las que ambas especies se benefician; o parasitarias, en las que la interacción perjudica a una especie y beneficia a la otra.

El comensalismo ocurre cuando una especie se beneficia de una interacción estrecha y prolongada, mientras que la otra no se beneficia ni se ve perjudicada. Las aves que anidan en árboles proporcionan un ejemplo de una relación comensal (Figura\(\PageIndex{8}\)). El árbol no se ve perjudicado por la presencia del nido entre sus ramas. Los nidos son ligeros y producen poca tensión en la integridad estructural de la rama, y la mayoría de las hojas, que el árbol utiliza para obtener energía por fotosíntesis, están por encima del nido por lo que no se ven afectadas. El ave, por otro lado, se beneficia enormemente. Si el ave tuviera que anidar al aire libre, sus huevos y crías serían vulnerables a los depredadores. Muchas relaciones comensales potenciales son difíciles de identificar porque es difícil demostrar que una pareja no obtiene algún beneficio de la presencia del otro.
Un segundo tipo de relación simbiótica se llama mutualismo, en el que dos especies se benefician de su interacción. Por ejemplo, las termitas tienen una relación mutualista con los protistas que viven en el intestino del insecto (Figura\(\PageIndex{9}\) a). La termita se beneficia de la capacidad de los protistas para digerir celulosa. Sin embargo, los protistas son capaces de digerir la celulosa solo por la presencia de bacterias simbióticas dentro de sus células que producen la enzima celulasa. La termita en sí no puede hacer esto; sin los protozoos, no podría obtener energía de su alimento (celulosa de la madera que mastica y come). Los protozoos se benefician al tener un ambiente protector y un suministro constante de alimentos de las acciones de masticación de madera de las termitas. A su vez, los protistas se benefician de las enzimas proporcionadas por sus endosimbiontes bacterianos, mientras que las bacterias se benefician de un ambiente doblemente protector y una fuente constante de nutrientes de dos huéspedes. Los liquen son una relación mutualista entre un hongo y algas fotosintéticas o cianobacterias (Figura\(\PageIndex{9}\) b). La glucosa producida por las algas proporciona alimento para ambos organismos, mientras que la estructura física del liquen protege a las algas de los elementos y hace que ciertos nutrientes en la atmósfera estén más disponibles para las algas. Las algas de los líquenes pueden vivir independientemente dado el ambiente adecuado, pero muchos de los socios fúngicos no pueden vivir solos.

Un parásito es un organismo que se alimenta de otro sin matar inmediatamente al organismo del que se está alimentando. En el parasitismo, el parásito se beneficia, pero el organismo del que se alimenta, el huésped, se ve perjudicado. El hospedador suele ser debilitado por el parásito ya que sifona recursos que el huésped normalmente utilizaría para mantenerse. Los parásitos pueden matar a sus huéspedes, pero generalmente hay selección para ralentizar este proceso y permitir que el parásito complete su ciclo reproductivo antes de que éste o sus crías puedan propagarse a otro huésped. El parasitismo es una forma de depredación.
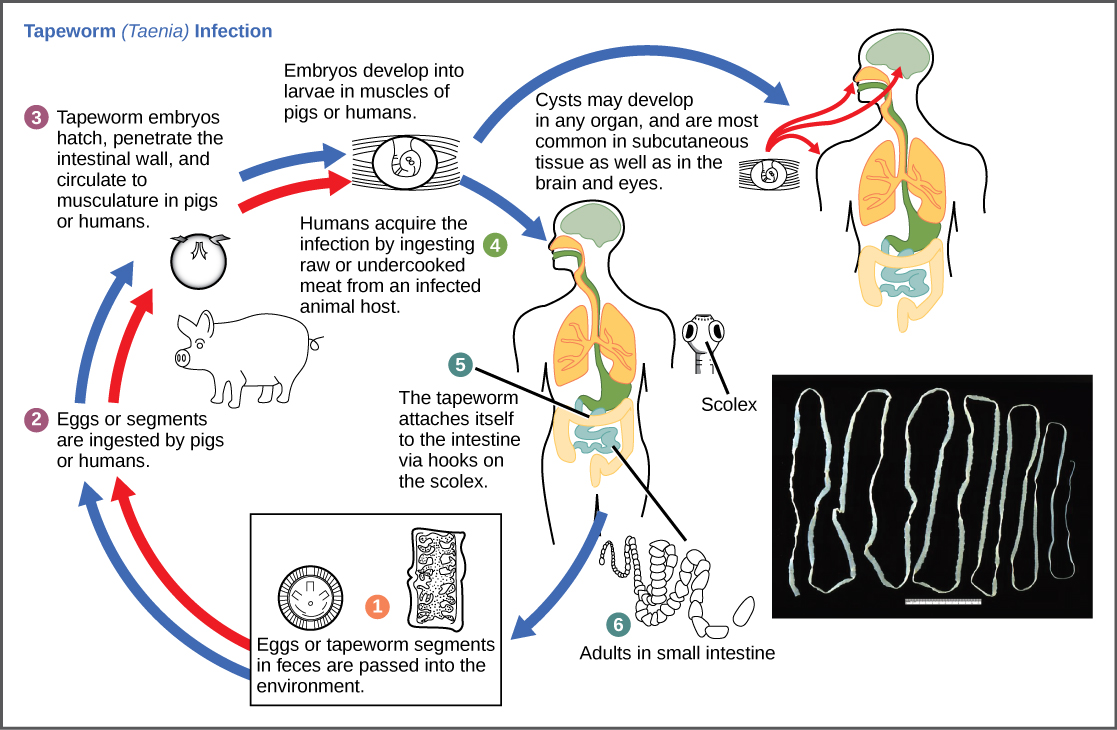
Los ciclos reproductivos de los parásitos suelen ser muy complejos, requiriendo a veces más de una especie hospedadora. Una tenia causa enfermedad en humanos cuando se consume carne contaminada y poco cocida como cerdo, pescado o carne de res (Figura\(\PageIndex{10}\)). La tenia puede vivir dentro del intestino del huésped durante varios años, beneficiándose de la comida del huésped, y puede crecer hasta alcanzar más de 50 pies de largo al agregar segmentos. El parásito se traslada de una especie hospedadora a una segunda especie hospedadora para completar su ciclo de vida.
Características de las Comunidades
Las comunidades son sistemas complejos que se pueden caracterizar por su estructura (el número y tamaño de las poblaciones y sus interacciones) y dinámicas (cómo cambian los miembros y sus interacciones con el tiempo). Comprender la estructura y dinámica de la comunidad nos permite minimizar los impactos en los ecosistemas y manejar comunidades ecológicas de las que nos beneficiamos.
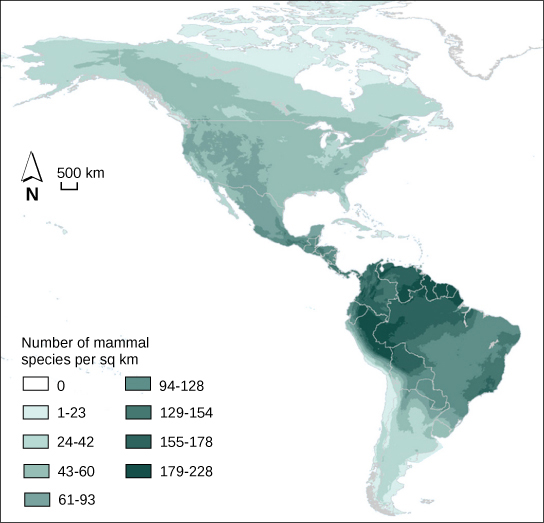
Los ecologistas han estudiado ampliamente una de las características fundamentales de las comunidades: la biodiversidad. Una medida de la biodiversidad utilizada por los ecologistas es el número de especies diferentes en un área en particular y su abundancia relativa. El área en cuestión podría ser un hábitat, un bioma o toda la biosfera. La riqueza de especies es el término utilizado para describir el número de especies que viven en un hábitat u otra unidad. La riqueza de especies varía en todo el mundo (Figura\(\PageIndex{11}\)). La riqueza de especies está relacionada con la latitud: la mayor riqueza de especies ocurre cerca del ecuador y la menor riqueza ocurre cerca de los polos. Las razones exactas de esto no se entienden claramente. Otros factores además de la latitud también influyen en la riqueza de especies. Por ejemplo, los ecologistas que estudiaban islas encontraron que la biodiversidad varía según el tamaño de la isla y la distancia del continente.
La abundancia relativa es el número de individuos en una especie en relación con el número total de individuos en todas las especies dentro de un sistema. Las especies de fundación, que se describen a continuación, suelen tener la mayor abundancia relativa de especies.
Las especies de cimentación son consideradas la “base” o “lecho rocoso” de una comunidad, teniendo la mayor influencia en su estructura general. Suelen ser productores primarios, y suelen ser un organismo abundante. Por ejemplo, las algas marinas, una especie de algas pardas, son una especie de cimentación que forma la base de los bosques de algas marinas frente a la costa de California.
Las especies fundacionales pueden modificar físicamente el ambiente para producir y mantener hábitats que beneficien a los demás organismos que las utilizan. Los ejemplos incluyen las algas descritas anteriormente o las especies arbóreas que se encuentran en un bosque. Los corales fotosintéticos del arrecife de coral también proporcionan estructura al modificar físicamente el ambiente (Figura\(\PageIndex{12}\)). Los exoesqueletos de coral vivo y muerto conforman la mayor parte de la estructura del arrecife, que protege a muchas otras especies de las olas y las corrientes oceánicas.

Una especie clave es aquella cuya presencia tiene una influencia desmesurada en el mantenimiento de la prevalencia de diversas especies en un ecosistema, la estructura de la comunidad ecológica y, a veces, su biodiversidad. Pisaster ochraceus, la estrella de mar intermareal, es una especie clave en la porción noroeste de los Estados Unidos (Figura\(\PageIndex{13}\)). Los estudios han demostrado que cuando este organismo es removido de las comunidades, las poblaciones de mejillones (su presa natural) aumentan, lo que altera completamente la composición de la especie y reduce la biodiversidad. Otra especie clave es el tetra bandeado, un pez en arroyos tropicales, que abastece casi todo el fósforo, nutriente inorgánico necesario, al resto de la comunidad. El tetra bandeado se alimenta en gran medida de insectos del ecosistema terrestre y luego excreta fósforo en el ecosistema acuático. Las relaciones entre las poblaciones de la comunidad, y posiblemente la biodiversidad, cambiarían drásticamente si estos peces se extinguieran.

BIOLOGÍA EN ACCIÓN
Las especies invasoras son organismos no nativos que, cuando se introducen en un área fuera de su área de distribución nativa, alteran la comunidad que invaden. En Estados Unidos, especies invasoras como la lisimaquia púrpura (Lythrum salicaria) y el mejillón cebra (Dreissena polymorpha) han alterado drásticamente los ecosistemas que invadieron. Algunos animales invasores conocidos incluyen el barrenador esmeralda del fresno (Agrilus planipennis) y el estornino europeo (Sturnus vulgaris). Ya sea disfrutando de una caminata por el bosque, haciendo un viaje en barco de verano o simplemente caminando por una calle urbana, es probable que se haya encontrado con una especie invasora.
Una de las muchas proliferaciones recientes de una especie invasora se refiere a la carpa asiática en Estados Unidos. Las carpas asiáticas fueron introducidas en los Estados Unidos en la década de 1970 por las pesquerías (estanques comerciales de bagres) y por las instalaciones de tratamiento de aguas residuales que utilizaron las excelentes capacidades de alimentación de filtro de los peces para limpiar sus estanques del exceso de plancton. Algunos de los peces escaparon, y en la década de 1980 habían colonizado muchas vías fluviales de la cuenca del río Mississippi, incluidos los ríos Illinois y Missouri.
Alimentadores voraces y reproductores rápidos, la carpa asiática puede superar a las especies nativas por la alimentación y podría conducir a su extinción. Una especie, la carpa herbívora, se alimenta de fitoplancton y plantas acuáticas. Compite con especies nativas por estos recursos y altera los hábitats de vivero para otros peces mediante la eliminación de plantas acuáticas. En algunas partes del río Illinois, la carpa asiática constituye el 95 por ciento de la biomasa de la comunidad. Aunque comestible, el pescado es huesudo y no deseado en Estados Unidos.
Los Grandes Lagos y sus preciadas pesquerías de salmón y trucha de lago están siendo amenazados por la carpa asiática. Las carpas aún no están presentes en los Grandes Lagos, y se está intentando impedir su acceso a los lagos a través del Chicago Ship and Sanitary Canal, que es la única conexión entre el río Mississippi y las cuencas de los Grandes Lagos. Para evitar que la carpa asiática salga del canal, se han utilizado una serie de barreras eléctricas para desalentar su migración; sin embargo, la amenaza es lo suficientemente significativa como para que varios estados y Canadá hayan demandado para que el canal de Chicago se corte permanentemente del lago Michigan. Los políticos locales y nacionales han influido en la manera de resolver el problema. En general, los gobiernos han sido ineficaces en la prevención o ralentización de la introducción de especies invasoras.
Dinámica de la comunidad
La dinámica comunitaria son los cambios en la estructura y composición de la comunidad a lo largo del tiempo, a menudo después de disturbios ambientales como volcanes, terremotos, tormentas, incendios y cambio climático. Se dice que las comunidades con un número relativamente constante de especies están en equilibrio. El equilibrio es dinámico con identidades de especies y relaciones que cambian con el tiempo, pero manteniendo números relativamente constantes. Después de una perturbación, la comunidad puede o no regresar al estado de equilibrio.
La sucesión describe la aparición y desaparición secuencial de especies en una comunidad a lo largo del tiempo después de una perturbación severa. En sucesión primaria, la roca recién expuesta o recién formada es colonizada por organismos vivos. En sucesión secundaria, una parte de un ecosistema se ve perturbada y quedan restos de la comunidad anterior. En ambos casos, hay un cambio secuencial en las especies hasta que se desarrolla una comunidad más o menos permanente.
Sucesión Primaria y Especies Pioneras
La sucesión primaria ocurre cuando se forman nuevas tierras, o cuando se elimina el suelo y toda la vida de la tierra preexistente. Un ejemplo de lo primero es la erupción de volcanes en la Isla Grande de Hawai, lo que resulta en lava que desemboca en el océano y forma continuamente nuevas tierras. De este proceso, se agregan aproximadamente 32 acres de tierra a la Isla Grande cada año. Un ejemplo de la remoción de suelo preexistente es a través de la actividad de los glaciares. El peso masivo del glaciar recorre el paisaje hasta el lecho rocoso mientras el glaciar se mueve. Esto elimina cualquier suelo original y deja la roca expuesta una vez que el glaciar se derrite y se retira.

En ambos casos, el ecosistema comienza con roca desnuda que está desprovista de vida. El nuevo suelo se forma lentamente a medida que la meteorización y otras fuerzas naturales descomponen la roca y conducen al establecimiento de organismos abundantes, como los líquenes y algunas plantas, que se conocen colectivamente como especies pioneras (Figura\(\PageIndex{14}\)) porque son las primeras en aparecer. Estas especies ayudan a descomponer aún más la roca rica en minerales en suelo donde otras especies menos resistentes pero más competitivas, como pastos, arbustos y árboles, crecerán y eventualmente reemplazarán a las especies pioneras. Con el tiempo la zona alcanzará un estado de equilibrio, con un conjunto de organismos muy diferentes a las especies pioneras.
Sucesión secundaria
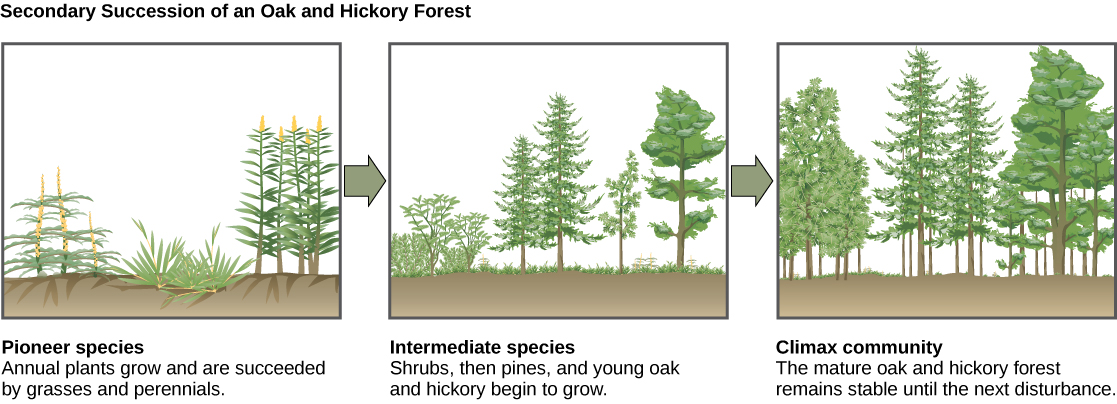
Un ejemplo clásico de sucesión secundaria ocurre en bosques talados por incendios forestales, o por tala de talones claros (Figura\(\PageIndex{15}\)). Los incendios forestales quemarán la mayor parte de la vegetación, y a menos que los animales puedan huir de la zona, son asesinados. Sus nutrientes, sin embargo, son devueltos al suelo en forma de ceniza. Así, aunque la comunidad ha sido dramáticamente alterada, existe un ecosistema de suelo presente que proporciona una base para una rápida recolonización.
Antes del incendio, la vegetación estaba dominada por árboles altos con acceso al principal recurso energético vegetal: la luz solar. Su altura les dio acceso a la luz solar al tiempo que sombreaban el suelo y otras especies bajas. Después del incendio, sin embargo, estos árboles ya no son dominantes. Así, las primeras plantas en volver a crecer suelen ser plantas anuales seguidas en pocos años por pastos de rápido crecimiento y propagación y otras especies pioneras. Debido, al menos en parte, a los cambios en el ambiente provocados por el crecimiento de pastos y forbs, a lo largo de muchos años, los arbustos emergen junto con pequeños árboles. Estos organismos se denominan especies intermedias. Eventualmente, a lo largo de 150 años o más, el bosque alcanzará su punto de equilibrio y se asemejará a la comunidad antes del incendio. A este estado de equilibrio se le conoce como la comunidad clímax, que permanecerá hasta el próximo disturbio. La comunidad clímax es típicamente característica de un clima y geología dados. Aunque la comunidad en equilibrio se ve igual una vez que se alcanza, el equilibrio es dinámico con constantes cambios en la abundancia y en ocasiones en las identidades de especies.
Colaboradores y Atribuciones
- Ecología Comunitaria de OpenStax está licenciado bajo CC BY 4.0. Modificado del original por Matthew R. Fisher.