
12.3A: Moléculas del Complejo Mayor de Histocompatibilidad (MHC)
- Page ID
- 55447

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Indicar qué células del cuerpo muestran moléculas de superficie MHC-I y qué células normalmente muestran moléculas de superficie MHC-II.
- Definir antígeno endógeno y antígeno exógeno y establecer qué clase de molécula MHC se une principalmente a cada uno.
- Indicar qué tipo de linfocito T reconoce epítopos de antígenos proteicos en moléculas de MHC-I y qué tipo reconoce epítopos de antígenos proteicos en moléculas de MHC-II.
- Indicar el papel de los proteasomas en la unión de péptidos de antígenos endógenos por moléculas de MHC-I.
- Indicar el papel de los lisosomas en la unión de péptidos de antígenos exógenos por moléculas MHC-II.
El papel de las moléculas del MHC en las respuestas inmunitarias adaptativas
Las moléculas MHC permiten que los linfocitos T reconozcan epítopos de antígenos y se discriminen a sí mismos de los no propios. A diferencia de los receptores de células B en linfocitos B que son capaces de unirse directamente a epítopos en antígenos, los receptores de células T (TCR) de los linfocitos T solo pueden reconocer epítopos -típicamente cadenas cortas de aminoácidos llamados péptidos- después de que se unan a moléculas del MHC (Figura\(\PageIndex{1}\)).

Los genes MHC son los genes más polimórficos del genoma humano, poseyendo muchos alelos por cada gen. Los genes MHC se expresan de manera co-dominante para que un individuo exprese los alelos heredados de cada progenitor. De esta manera, se maximiza el número de moléculas MHC que se unen al péptido para su presentación a los linfocitos T. Además, cada molécula MHC es capaz de unirse a una amplia variedad de péptidos diferentes, tanto autopéptidos como péptidos extraños. Existen dos clases de moléculas MHC: MHC-I y MHC-II.
- Las moléculas de MHC-I presentan epítopos a linfocitos T8.
- Las moléculas MHC-II presentan epítopos a linfocitos T4.
La expresión de las moléculas del MHC es incrementada por las citocinas producidas tanto durante las respuestas inmunitarias innatas como las respuestas inmunes adaptativas. Las citocinas como el interferón alfa, interferón beta, interferón gamma, factor de necrosis tumoral aumentan la expresión de las moléculas MHC-I, mientras que el interferón gamma es la citocina principal para aumentar la expresión de las moléculas de MHC-II.
Moléculas MHC-I
Las moléculas MHC-I están diseñadas para permitir que el cuerpo reconozca las células infectadas y las células tumorales y las destruya con linfocitos T citotóxicos o CTL. Los CTL son células de defensa efectoras derivadas de linfocitos T8 sin tratamiento previo. Las moléculas MHC-I son:
- Elaborado por todas las células nucleadas del cuerpo.
- Poseer un surco profundo que pueda unirse a epítopos peptídicos, típicamente de 8-11 aminoácidos de longitud, típicamente de antígenos endógenos.
- Presentar complejos MHC-I/péptido a linfocitos T8 ingenuos y linfocitos T citotóxicos que poseen un receptor de células T de forma complementaria o TCR.
- A través del proceso de presentación cruzada, algunas células dendríticas presentadoras de antígeno pueden presentar de manera cruzada epítopos de antígenos exógenos a moléculas MHC-I para su eventual presentación a linfocitos T8 ingenuos.
Los antígenos endógenos son proteínas que se encuentran dentro del citosol de las células humanas. Los ejemplos de antígenos endógenos incluyen:
a. Proteínas virales producidas durante la replicación viral;
b. Proteínas producidas por bacterias intracelulares como Rickettsias y Chlamydias durante su replicación;
c. Proteínas que han escapado al citosol desde el fagosoma de fagocitos como las células presentadoras de antígeno;
d. Antígenos tumorales producidos por células cancerosas; y
e. Autopéptidos de proteínas celulares hospedadoras.
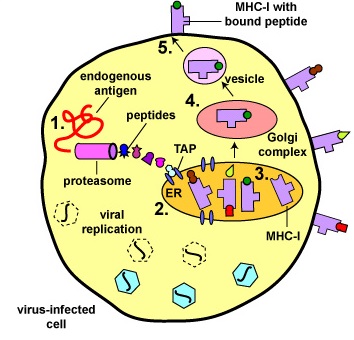
Durante la replicación de virus y bacterias intracelulares dentro de su célula hospedadora, así como durante la replicación de células tumorales, las proteínas virales, bacterianas o tumorales se degradan en una variedad de epítopos peptídicos por orgánulos cilíndricos llamados proteasomas. Las proteínas citosólicas propias del cuerpo también son degradadas en péptidos por los proteasomas.
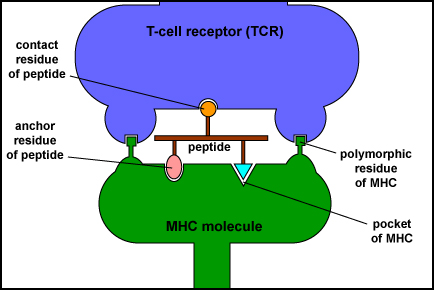
Estos epítopos peptídicos se unen luego a un surco de moléculas MHC-I que luego se transportan a la superficie de esa célula donde pueden ser reconocidos por un receptor de células T de forma complementaria (TCR) y una molécula CD8, un correceptor, en la superficie de un linfocito T8 ingenuo o un linfocito T citotóxico ( CTL). Los TCR reconocen tanto el antígeno peptídico extraño como la molécula MHC (Figura\(\PageIndex{2}\)). Los TCR, sin embargo, no reconocerán los autopéptidos unidos a MHC-I. En consecuencia, las células normales no son atacadas y asesinadas.

Las células dendríticas unen epítopos de antígenos endógenos a moléculas de MHC-I y los presentan a linfocitos T8 vívidos para activar estos linfocitos T8 vívidos.
- Los antígenos son engullidos por células dendríticas y colocados en un fagosoma. Algunas de las proteínas escapan del fagosoma al citosol de la célula dendrítica donde se convierten en antígenos endógenos.
- Estos antígenos endógenos pasan a través de proteasomas donde se degradan en una serie de péptidos.
- Los péptidos son transportados al retículo endoplásmico rugoso (ER) por una proteína transportadora llamada TAP.
- Los péptidos luego se unen a los surcos de moléculas MHC-I recién sintetizadas.
- El retículo endoplásmico transporta las moléculas MHC-I con péptidos unidos al complejo de Golgi.
- El complejo de Golgi, a su vez, transporta los complejos MHC-I/péptido a través de una vesícula exocítica a la membrana citoplásmica donde se anclan. Aquí, el péptido y los complejos MHC-I/péptido pueden ser reconocidos por linfocitos T8 vívidos por medio de TCR y moléculas CD8 que tienen una forma complementaria.
A través del proceso de presentación cruzada, algunas células dendríticas presentadoras de antígeno pueden presentar de manera cruzada epítopos de antígenos exógenos a moléculas MHC-I para su eventual presentación a linfocitos T8 ingenuos.
La molécula MHC-I con péptido unido en la superficie de células dendríticas presentadoras de antígeno (Figura\(\PageIndex{3}\)) puede ser reconocida por un TCR/CD8 de forma complementaria en la superficie de un linfocito T8 sin tratamiento previo para iniciar la inmunidad mediada por células (Figura\(\PageIndex{4}\)). (Ciertas células dendríticas, como se discutirá más adelante, también pueden presentar antígenos exógenos a moléculas de MHC-I).
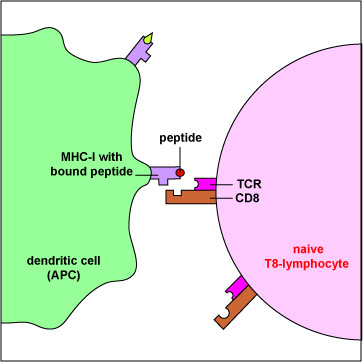
La molécula MHC-I con péptido unido en la superficie de células infectadas y células tumorales (Figura\(\PageIndex{5}\)) puede ser reconocida por un TCR/CD8 de forma complementaria en la superficie de un linfocito T citotóxico o CTL para iniciar la destrucción de la célula que contiene el antígeno endógeno (Figura\(\PageIndex{6}\)). (Los CTL son células efectoras derivadas de linfocitos T8 sin tratamiento previo).
Los linfocitos T citotóxicos (CTL) son capaces entonces de reconocer complejos péptido/MHC-I por medio de sus receptores de células T (TCR) y moléculas CD8 y matar las células a las que se unen.
- Durante la replicación viral dentro de la célula hospedadora, los antígenos endógenos, como las proteínas virales, pasan a través de proteasomas donde se degradan en una serie de péptidos.
- Los péptidos son transportados al retículo endoplásmico rugoso (ER) por una proteína transportadora llamada TAP.
- Los péptidos luego se unen a los surcos de moléculas MHC-I recién sintetizadas.
- El retículo endoplásmico transporta las moléculas MHC-I con péptidos unidos al complejo de Golgi.
- El complejo de Golgi, a su vez, transporta los complejos MHC-I/péptido a través de una vesícula exocítica a la membrana citoplásmica donde se anclan. Aquí, el péptido y los complejos MHC-I/péptido pueden ser reconocidos por CTL por medio de TCR y moléculas CD8 que tienen una forma complementaria.
Las moléculas MHC-I están codificadas por tres genes MHC-I, HLA-A, HLA-B y HLA-C. Como se mencionó anteriormente, sin embargo, existen muchos alelos diferentes para cada gen que hereda una persona. De esta manera, se maximiza el número de moléculas MHC-I que unen péptidos para su presentación a linfocitos T-8. La expresión de las moléculas MHC-I en todos los tipos celulares se ve incrementada por las citocinas interferón-alfa (IFN-a) e interferón-beta (IFN-ß).

Ejercicio: Preguntas de Pensar-Par-Compartir
Todas las células nucleadas producen moléculas MHC-I. Las moléculas MHC-I se unen a epítopos peptídicos de antígenos que se encuentran dentro de nuestras células. Los epítopos peptídicos unidos a moléculas de MHC-I son reconocidos por TCR y moléculas CD8 en las superficies de linfocitos T8 vívidos y en linfocitos T citotóxicos (CTL).
¿Por qué es importante que todas las células nucleadas de nuestro cuerpo sean capaces de producir moléculas MHC-I?
Moléculas MHC-II
Las moléculas MHC-II están diseñadas para permitir que los linfocitos T4 reconozcan epítopos de antígenos exógenos y se discriminen a sí mismos de los no propios. Las moléculas MHC-II son:
- Elaborado por células presentadoras de antígeno o APC, como células dendríticas, macrófagos y linfocitos B.
- Posee un surco profundo que puede unirse a epítopos peptídicos, a menudo de 10-30 aminoácidos de largo pero con una longitud óptima de 12-16 aminoácidos, típicamente de antígenos exógenos. Los péptidos interactúan a lo largo de toda su longitud con el surco.
- Presentar complejos MHC-II/péptido a linfocitos T4 ingenuos o linfocitos T4 efectores que tienen un receptor de células T o TCR de forma complementaria.
- A través del proceso de presentación cruzada, algunas células dendríticas presentadoras de antígeno pueden presentar de manera cruzada epítopos de antígenos endógenos a moléculas de MHC-II para su eventual presentación a linfocitos T4 vívidos.
Los antígenos exógenos son antígenos que ingresan desde fuera del cuerpo, como bacterias, hongos, protozoos y virus libres. Estos antígenos exógenos ingresan a macrófagos, células dendríticas y linfocitos B a través de la fagocitosis. Los microbios son engullidos y colocados en un fagosoma que luego se fusiona con lisosomas. Después de esta fusión, el fagolisosoma se acidifica. La acidificación, a su vez, activa las proteasas dentro del fagolisosoma permitiendo que los antígenos proteicos del microbio se degraden en una serie de péptidos cortos. Estos epítopos peptídicos se unen luego a moléculas de MHC-II y luego se transportan a la superficie de la célula presentadora de antígeno (APC) (Figura\(\PageIndex{7}\)). (Ciertas células dendríticas, como se discutirá más adelante, también pueden presentar antígenos endógenos a moléculas de MHC-II).
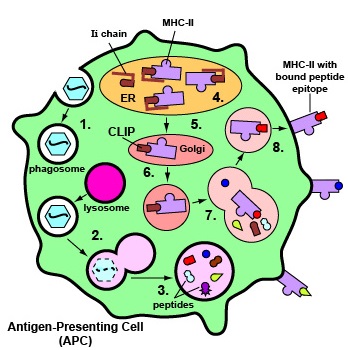
Algunos patógenos, como Mycobacterium tuberculosis, Mycobacterium leprae y Leishmania, son capaces de crecer en las vesículas encíticas de macrófagos sin ser destruidos por los lisosomas. Estos macrófagos pueden, sin embargo, ser activados por linfocitos efectores T4 llamados células T H 1 y posteriormente usar proteasas intravesiculares para degradar las proteínas de estos patógenos en péptidos para su presentación a moléculas de MHC-II que pasan a través de su camino a la superficie celular.
Aquí las moléculas MHC-II con péptidos unidos pueden ser reconocidas por un receptor de células T de forma complementaria y una molécula CD4, un correceptor, en la superficie de un linfocito T4 (Figura\(\PageIndex{8}\)). Los linfocitos T4 son las células que el cuerpo utiliza para regular tanto la inmunidad humoral como la inmunidad mediada por células.

Las moléculas MHC-II están codificadas por tres genes MHC-II, HLA-DR, HLA-DP y HLA-DQ. Interferón gamma (IFN-? ) aumenta la expresión de moléculas tanto MHC-I como MHC-II.
Preguntas de Think-Pair-Share
Solo las células presentadoras de antígeno como las células dendríticas, los macrófagos y los linfocitos B producen moléculas MHC-II. Los epítopos peptídicos unidos a moléculas MHC-II son reconocidos por TCR y moléculas CD4 en las superficies de linfocitos T4 vívidos y en linfocitos T4 efectores.
¿Por qué no todas las células nucleadas de nuestro cuerpo producen moléculas MHC-II así como moléculas MHC-I?
¿Por qué es importante que las células dendríticas produzcan moléculas tanto MHC-I como MHC-II?
Resumen
- Las moléculas MHC permiten que los linfocitos T reconozcan epítopos y se discriminen a sí mismos de los no propios.
- Los receptores de células T (TCR) de los linfocitos T solo pueden reconocer epítopos -típicamente cadenas cortas de aminoácidos llamados péptidos- después de que se unan a las moléculas del MHC.
- MHC-I presenta epítopos a linfocitos T8; MHC-II presenta epítopos a linfocitos T4.
- Las moléculas MHC-I están diseñadas para permitir que el cuerpo reconozca las células infectadas y las células tumorales y las destruya con linfocitos T citotóxicos o CTL. (Los CTL son células de defensa efectoras derivadas de linfocitos T8 sin tratamiento previo).
- Las moléculas de MHC-I son elaboradas por todas las células nucleadas del cuerpo; se unen a epítopos peptídicos típicamente de antígenos endógenos; presentan complejos MHC-I/péptido a linfocitos T8 vívidos y linfocitos T citotóxicos que poseen un receptor de células T de forma complementaria o TCR.
- A través del proceso de presentación cruzada, algunas células dendríticas presentadoras de antígeno pueden presentar de manera cruzada epítopos de antígenos exógenos a moléculas MHC-I para su eventual presentación a linfocitos T8 ingenuos.
- Los antígenos endógenos son proteínas que se encuentran dentro del citosol de células humanas e incluyen proteínas virales producidas durante la replicación viral, proteínas producidas por bacterias intracelulares, proteínas que han escapado al citosol desde el fagosoma de fagocitos como las células presentadoras de antígeno y antígenos tumorales producido por las células cancerosas.
- Durante la replicación de virus y bacterias intracelulares dentro de su célula hospedadora, así como durante la replicación de células tumorales, las proteínas virales, bacterianas o tumorales se degradan en una variedad de epítopos peptídicos por orgánulos cilíndricos llamados proteasomas. Los epítopos peptídicos resultantes se unen luego a moléculas de MHC-I que luego se transportan a la superficie de esa célula.
- Los antígenos exógenos son antígenos que ingresan desde fuera del cuerpo como bacterias, hongos, protozoos y virus libres.
- Las moléculas de MHC-II se fabrican mediante células presentadoras de antígeno o APC, tales como células dendríticas, macrófagos y linfocitos B; se unen a epítopos peptídicos típicamente de antígenos exógenos; y presentan complejos MHC-II/péptido a linfocitos T4 vívidos o linfocitos T4 efectores que tienen un receptor de células T de forma complementaria o TCR.
- A través del proceso de presentación cruzada, algunas células dendríticas presentadoras de antígeno pueden presentar de manera cruzada epítopos de antígenos endógenos a moléculas de MHC-II para su eventual presentación a linfocitos T4 vívidos.
- Los antígenos exógenos ingresan a macrófagos presentadores de antígeno, células dendríticas y linfocitos B a través de la fagocitosis, y son engullidos y colocados en un fagosoma donde los antígenos proteicos del microbio son degradados por proteasas en una serie de péptidos. Estos péptidos se unen luego a moléculas MHC-II que luego se ponen en la superficie de la APC.