Explicar las tendencias básicas de la evolución humana incluyendo el bipedalismo y la encefalización.
Describir los homínidos y lo que distingue a los homínidos de otros miembros del orden de los Primates.
Describir la categoría de características que distingue a los proto-homínidos de los homínidos.
Describir, incluyendo rangos de fechas aproximados, la evolución del género Homo, incluyendo especies e arly Homo y humanos modernos.
Explicar cómo la cultura material nos ayuda a informar sobre la psicología del Homo sapiens ancestral y otras especies de homínidos.
Visión general
Tendencias: Hay una serie de tendencias en la evolución de los proto-homínidos (especies primitivas parecidas a simios consideradas posiblemente ancestrales para los humanos modernos) al Homo sapiens moderno. Estos rasgos no ocurren todos a la vez, sino a lo largo de millones de años.
Proto-homínidos y homínidos: Al determinar qué características fósiles debe tener un espécimen para ser clasificado como homínido (el término utilizado para los humanos y sus antepasados después de la división con chimpancés y bonobos), se examinan muchas características diferentes, entre ellas las relacionadas con el bipedalismo y la odontología características relacionadas con la masticación. Los simios tienen un espacio entre morder los dientes y rechinar los dientes donde el canino superior encaja cuando las mandíbulas se cierran. También se consideran otras características como el tamaño cerebral y corporal.
Homo Género: La aparición del género Homo (nuestro género) marca el advenimiento de cerebros más grandes, el surgimiento de la cultura material (por ejemplo, herramientas de piedra) y la eventual colonización del mundo fuera de África.
Cultura material: La evidencia más temprana de la cultura material se encuentra en forma de herramientas de piedra encontradas en sitios que datan de hace 2.4 millones de años (ver Contenido Complementario, Capítulo 18, Cultura Material).
Figura\(\PageIndex{1}\): El cráneo original completo (sin dientes superiores ni mandíbula) de un ejemplar de Australopithecus africanus de 2.1 millones de años llamado “Mrs. Ples” (número de catálogo STS 5, cueva Sterkfontein, fósil homínido número 5), descubierto en Sudáfrica. Colección del Museo Transvaal, Northern Flagship Institute, Pretoria, Sudáfrica. (CC BY-SA 4.0; José Braga; Didier Descouens).
Caminos evolutivos humanos y simios: divergencia de ancestros comunes
Antes de comenzar nuestra discusión sobre la evolución humana, será útil echar un vistazo al panorama general. Las dos figuras a continuación junto con la Figura 3.7.16, mucho más abajo, representan colectivamente 20 millones de años de evolución que llevaron a nuestra especie, el Homo sapiens. La Figura 3.7.2 muestra los últimos 10 millones de años de evolución humanoide. La Figura 3.7.3 muestra los últimos 20 millones de años de evolución humanoide y así es inclusiva de la Figura 3.7.2, que muestra mayor detalle comenzando con la filogenia de los homínidos (homínidos africanos incluidos los humanos), 10 Mya para presentar. Ambas figuras muestran nuestro género Homo en la parte superior izquierda de cada diagrama. La Figura 3.7.16 muestra una representación ampliada y mucho más detallada del género Homo incluyendo las relaciones entre Homo sapiens y neandertales y el hecho de que estas dos especies alguna vez coexistieron al mismo tiempo fuera de África hace unos 30,000-40,000 años. En las dos primeras cifras inmediatamente abajo, notamos la gran divergencia en un número sorprendentemente grande de especies a lo largo de millones de años. Observe también lo pequeño y lo reciente que es el género Homo en comparación, y cómo una especie muy reciente que surgió durante el Pleistoceno, los humanos, ha llegado tan rápidamente a dominar la Tierra, al menos por ahora. Se espera que al examinar de cerca estas cifras, puedas organizar mejor los detalles que siguen en esta sección y que las puedas entender dentro del contexto más amplio de la evolución de los primates.
Figura\(\PageIndex{2}\): Hominini (incluye Homo y Pan, pero excluye gorilas) y Homininae (homínidos africanos incluyendo humanos) filogenia, 10 Mya para presentar; la rama humana que comienza con Homo, arriba a la izquierda, se agranda y se muestra con mayor detalle en la Figura 3.7.16 más cerca de la parte inferior de esta página. La línea ancestral Hominin (humanos modernos, especies humanas extintas y todos nuestros antepasados inmediatos, incluidos los miembros de los géneros Homo, Australopithecus, Paranthropus y Ardipithecus) y la línea de simios, incluidos los gorilas, divergieron unas de otras unas 8 Mya (hace millones de años) y las líneas de Hominin y Pan (chimpancé) divergen unas de otras alrededor de 6 Mya. Homo sapiens se muestran en la parte superior izquierda. Para retroceder 20 Mya, véase la Figura 3.7.3 a continuación. Ahí se puede ver que Pongo (orangutanes) divergió de la línea Hominin mucho antes, hace unos 14 millones de años (Imagen de Wikimedia Commons; Archivo:Hominini lineage.svg; https://commons.wikimedia.org/wiki/F...ni_lineage.svg; por Dbachmann; licenciada bajo la CreativeCommons Reconocimiento-Compartir Igual 4.0 Licencia internacional).
Figura\(\PageIndex{3}\): Filogenia de Hominoidea, 20 Mya para presentar. Un hominoide, a veces llamado simio, es miembro de la superfamilia Hominoidea: los miembros existentes son los gibones (simios menores, familia Hylobatidae) y los homínidos. Un homínido es un miembro de la familia Hominidae, los grandes simios: orangutanes (Ponginae; Pongo), gorilas (Gorillini), chimpancés (Pan) y humanos. Obsérvese que la división entre la línea Homo, que lleva a los humanos, de los linajes chimpancé y gorila ocurrió hace unos seis millones y ocho millones de años, respectivamente. (Imagen de Wikimedia Commons; Archivo:Hominoidea lineage.svg; https://commons.wikimedia.org/wiki/F...ea_lineage.svg; por Dbachmann; licenciado bajo la licencia Creative CommonsReconocimiento-Compartir Igual 4.0 Internacional).
Tendencias en la evolución humana
Una serie de preguntas sobre la evolución humana son:
¿Por qué nuestros primeros antepasados se pusieron de pie?
¿Por qué algunas especies se extinguieron hasta que solo quedó una especie, el Homo sapiens?
¿Cuándo, dónde y por qué evolucionaron los humanos modernos?
¿Cuál fue el papel de los neandertales?
¿Qué nos hace humanos?
Si bien las hipótesis sugieren respuestas, la investigación continúa refinando nuestra comprensión de la evolución humana.
Esta sección proporciona una visión general de los principales hallazgos y tendencias en la evolución de los homínidos. (Para obtener información adicional, consulte la sección “Para mayor exploración” al final de esta página).
Figura\(\PageIndex{4}\): Un hombre contemplando su evolución.
Definición de homínidos
Es a través de nuestro estudio de nuestros antepasados y parientes homínidos que estamos expuestos a un mundo de “podría haber sido” —de otros caminos no tomados por nuestra especie, de otras formas de ser humanos. Pero para entender mejor estas diferentes trayectorias evolutivas, primero debemos definir los términos que estamos utilizando. Si se trazara una línea imaginaria entre nosotros y nuestros parientes más cercanos, los grandes simios, el bipedalismo (habitualmente caminando erguidos sobre dos pies) es donde se trazaría esa línea. Hominin, entonces, significa todos en “nuestro” lado de esa línea definida por el bipedalismo. Así, los homínidos son humanos y todos nuestros ancestros bípedos y parientes extintos desde nuestra divergencia del último ancestro común (ACV) que compartimos con los chimpancés. Como se muestra anteriormente en las Figuras 3.3.2 y 3.3.3, muchos científicos creen que esta división entre linajes humanos y chimpancé tuvo lugar hace unos 6 millones de años. sin embargo, Los datos del ADN que utilizan tasas de mutación estimadas sugieren que la división entre los linajes humano y simio puede haber ocurrido mucho antes: la divergencia entre las líneas humanas y de chimpancé tuvo lugar alrededor de 7 a 8 millones y posiblemente hasta 13 millones de años atrás, mientras que la divergencia entre humanos y Las líneas de gorila pueden haber ocurrido hace tanto tiempo como hace 8 a 19 millones de años (Gibbons, 2012). Estas diferentes estimaciones pueden resultar confusas para los estudiantes. Sin embargo, esta amplia gama de estimaciones sirve como un buen ejemplo de cómo nuestro conocimiento es incompleto y a veces cambia a medida que surgen nuevos descubrimientos y métodos. En este contexto, es mejor tener presente que “[...] cuando todo se dice y se hace una taxonomía es solo una hipótesis; no está escrita en tablillas de piedra” (Wood, 2010, p. 8908).
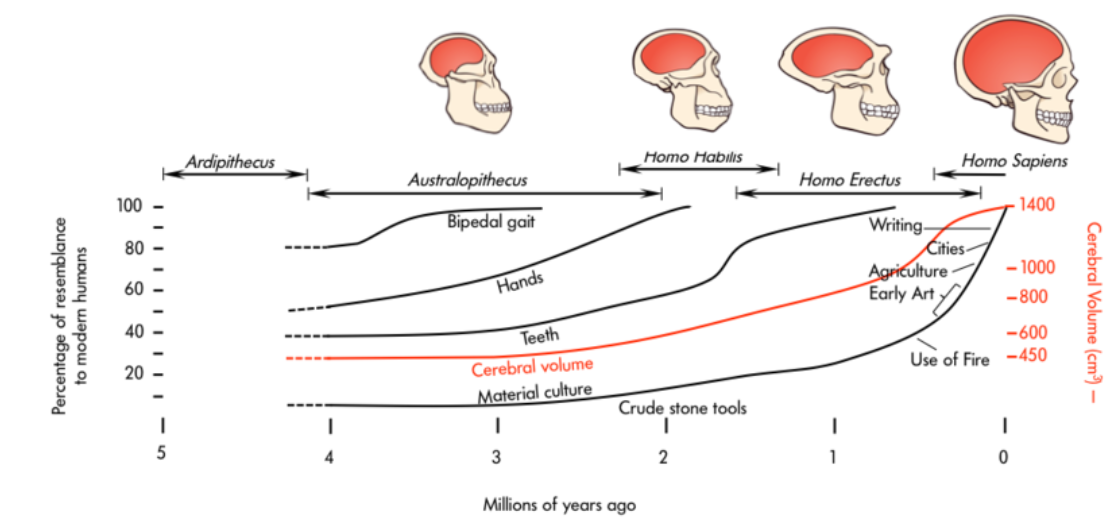
Tendencias morfológicas en la evolución humana
Hay una serie de tendencias en la evolución del linaje humano desde los proto-homínidos (especies ancestrales humanas tempranas) hasta el Homo sapiens moderno. Estos rasgos no ocurren todos a la vez, sino a lo largo de millones de años. En general, las tendencias incluyen:
el movimiento hacia adelante del foramen magnum (el agujero en la parte inferior del cráneo donde la médula espinal ingresa al cráneo para conectarse con el cerebro): relacionado con la postura erguida
una reducción en el tamaño de los caninos
un aumento en el tamaño de los molares
un aumento en la capacidad craneal
aplanamiento de la cara
redondeo del cráneo
Nuevamente, no todos estos rasgos ocurren al mismo tiempo y hay variación entre las diversas especies de homínidos, pero todas estas características morfológicas se dan en la línea evolutiva del Homo sapiens. Otras dos tendencias son especialmente importantes en la evolución de los homínidos:
bipedalismo, y
encefalización del cerebro.
Estos se discuten con más detalle a continuación.
Bipedalismo
El bipedalismo, o caminar erguido, fue el primer rasgo morfológico en el camino hacia la humanidad. El bipedalismo humano es la forma primaria de moverse (esto se llama bipedalismo habitual). Otros primates practican la locomoción bípeda temporal u ocasional, por ejemplo, los primates como los chimpancés pueden caminar bipedalmente mientras llevan algo en sus manos. Pocos otros animales son bípedos habituales como los humanos, por ejemplo, aves y canguros.
Existen numerosos cambios anatómicos que evolucionaron para hacer que los homínidos locomotoras bípedas eficientes, incluyendo cambios en la posición del foramen magnum (ver arriba), la columna vertebral de la columna vertebral, la pelvis, fémur y tibia, articulación de la rodilla y estructura del pie, muchos de estos cambios están relacionados con la mejora equilibrio y absorción de impactos (eLucy 2007).
Los cambios morfológicos asociados al bipedalismo tardan millones de años en evolucionar. Primero aparecen hace 6.0-7.0 millones de años (mya), pero no están completamente en su lugar hasta alrededor de 4.0 mya. Estos cambios físicos continúan refinándose hasta que los vemos como lo hacemos hoy en día en el Homo sapiens moderno (Jurmain et al. 2013).
Hipótesis sobre la evolución del bipedalismo
A lo largo del siglo pasado se han propuesto varias hipótesis para explicar la evolución de los homínidos. Como el bipedalismo es el primer rasgo en el camino hacia los humanos modernos, estas hipótesis se centran en el surgimiento del bipedalismo habitual. Muchos han sido refutados a medida que se descubren nuevos datos. La primera hipótesis fue la hipótesis de caza propuesta por Charles Darwin. La hipótesis de la caza afirma que la clave de la evolución humana fue el cambio de una vida arbórea a una terrestre. Predijo que los primeros homínidos se encontrarían en África basándose en las similitudes que vio entre humanos y simios africanos. Sugirió que el bipedalismo otorgaba una ventaja a los primeros homínidos ya que liberaba sus manos para portar armas utilizadas para cazar animales. Darwin también sugirió que cerebros más grandes precedieron al bipedalismo ya que se necesitaba inteligencia para hacer las herramientas. Ahora sabemos que el bipedalismo habitual es anterior a los cerebros grandes por lo que la hipótesis de Darwin ya no se considera una explicación adecuada. Con el descubrimiento de nuevos datos, se han propuesto otras hipótesis incluyendo las hipótesis de bosque de parches y aprovisionamiento.
La hipótesis del bosque irregular sugiere que el entorno emergente de mosaico que comenzó a finales del Mioceno hizo que el bipedalismo fuera ventajoso. La frase ambiente mosaico en este caso se refiere a un ambiente que tenía bosque irregular intercalado con pastizales que finalmente se convirtieron en las sabanas africanas de hoy. Esto provocó que los recursos alimentarios se extendieran por el paisaje. Para viajar largas distancias, el bipedalismo es más eficiente energéticamente que el cuadrupedalismo (caminar a cuatro patas). Viajar bipedalmente liberó las manos para llevar provisiones y los primeros homínidos podrían haberse alimentado fácilmente de recursos terrestres y arbóreos.
La hipótesis de aprovisionamiento establece que tener las manos libres para transportar alimentos permitió a los machos abastecer a las hembras y crías. Dado que gran parte de la energía femenina se destinó a la crianza de los hijos, la capacidad de un macho para abastecerla a ella y a su descendencia habría sido una cualidad atractiva. Aquellos machos que pudieran caminar de manera más eficiente bipedalmente mientras transportaban alimentos habrían sido material de primer mate, permitiendo que tanto el macho como la hembra se reproduzcan con éxito. Sin embargo, especies como los lobos proveen a sus hembras y crías a pesar de que son cuadrúpedos al consumir carne y regurgitarla a los cachorros.
La verdad del asunto es que los orígenes del bipedalismo siguen siendo turbios. Es de esperar que más investigaciones nos ayuden a acercarnos a una determinación de por qué evolucionó el bipedalismo, y de ahí nuestros primeros antepasados. Mientras tanto, se pueden explorar otras hipótesis sobre los orígenes del bipedalismo en el sitio web de NOVA: http://www.pbs.org/wgbh/nova/evoluti...ipedalism.html [opcional].
Características dentales
Los simios tienen un complejo de masticación que es bueno para cortar y triturar alimentos. Con el tiempo, los homínidos pierden esta característica dental, ya que el canino disminuye de tamaño y los molares aumentan de tamaño (Larsen 2014).
Evolución del Cerebro
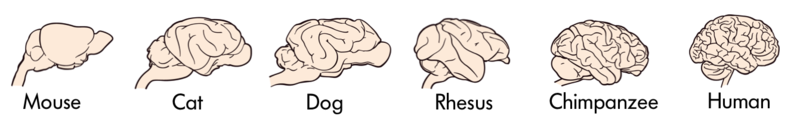
En relación con otros mamíferos, los primates tienen un cerebro más expandido y elaborado, incluyendo la expansión de la corteza cerebral. Compara la complejidad del cerebro humano de la izquierda con el cerebro de gato de la derecha (las fotos no son a escala).
Figura\(\PageIndex{6}\): Cerebro Humano.Figura\(\PageIndex{7}\): Cerebro de Gato.
Claramente, se han producido cambios anatómicos significativos durante la evolución del cerebro en primates, en otros mamíferos y en animales en general. ¿Hay generalizaciones que se puedan hacer sobre la evolución de los cerebros animales?
Striedter (2006) ha identificado una serie de principios generales de evolución cerebral aplicables en una amplia gama de especies (es decir, no solo primates o mamíferos).
1) Los cerebros embrionarios entre especies son más similares que los cerebros adultos, porque los cerebros tienden a diversificarse más a medida que crecen hacia la forma adulta;
2) el tamaño del cerebro relativo al tamaño corporal en los vertebrados (animales con esqueletos) ha tendido a aumentar más a menudo que disminuir con el tiempo evolutivo;
3) parece que los aumentos en el tamaño relativo del cerebro fueron generalmente acompañados por aumentos en la complejidad de forrajeo social o alimentario;
4) la mayoría de los aumentos en el tamaño relativo del cerebro fueron acompañados por aumentos en el tamaño corporal absoluto;
5) los aumentos en el tamaño absoluto del cerebro requieren cambios en las conexiones internas del cerebro que implican una mayor modularidad o especialización (módulos de procesamiento especializados que incrementan la “división del trabajo” en lugar de que todo el cerebro realice todas las clases de procesamiento por igual) de la anatomía y funcionamiento del cerebro;
6) la evolución generalmente agranda los cerebros al extender el período de desarrollo cerebral antes y después del nacimiento mientras conserva (mantiene el mismo) el “orden de nacimiento” de diferentes regiones cerebrales, de modo que los animales de cerebro grande tienden a tener regiones “nacidas” tardías (de desarrollo tardío) desproporcionadamente más grandes (“tarde es igual a grande” ), como la corteza cerebral, lo que lleva a una corteza cerebral desproporcionadamente mayor (aumento de la corticalización) en mamíferos de cerebro grande (los no mamíferos no tienen corteza cerebral); sin embargo hay excepciones a esta regla, por ejemplo, en cualquier tamaño cerebral absoluto dado, hay más corteza cerebral en los simios que en prosimios, y en los loros hay un telencéfalo inusualmente grande (prosencéfalo) que no es contabilizado por la regla;
7) los cambios en las proporciones de tamaño de las áreas cerebrales, aunque “automáticos” dentro de las reglas de escalado (alométricas) anteriores, aún pueden ser adaptativos y someterse a selección natural;
8) a medida que las regiones cerebrales aumentan en tamaño absoluto o proporcional, tienden a laminarse, organizarse en láminas de neuronas, permitiendo conexiones punto por punto correspondientes entre mapas sensoriales y motores con cableado axonal y dendrítico mínimo, ahorrando espacio y energía metabólica;
9) a medida que aumenta el tamaño del cerebro, ocurren más subdivisiones regionales a partir de partes ancestrales que se subdividen en nuevas partes, como en el tálamo dorsal (ubicado debajo de la corteza cerca del centro del cerebro), o, como en el caso del neocórtex, se agregó una nueva parte a un conjunto ancestral de cerebro conservado (retenido sobre evolución) partes;
10) un principio conocido como regla de Deacon es que “grande es igual bien conectado”, lo que significa que a medida que aumenta el tamaño relativo de una estructura cerebral tiende a recibir más conexiones y a proyectar (enviar) más salidas a otras estructuras.
Striedter (2006) agrega una serie de generalizaciones adicionales sobre los cerebros de mamíferos y primates, incluyendo cerebros humanos:
11) la neocorteza mamífera de seis capas (que se encuentra solo en mamíferos como nosotros) probablemente evolucionó a partir de un precursor reptil de 3 capas llamado corteza dorsal (algo así como el que se encuentra en las tortugas) mediante la adición de varias capas de corteza;
12) aparte del neocórtex, el cerebro de los mamíferos es similar al cerebro reptil (que también tiene hipocampo, por ejemplo) pero incluso con un “esquema fundamental” de regiones cerebrales y circuitos, muchos cambios menores en el cableado pueden cambiar drásticamente la forma en que la información fluye a través de un cerebro y, por lo tanto, cómo funciona, el cerebro del mamífero no es solo una versión exclusiva del cerebro reptil;
13) el aumento de la corticalización en los mamíferos no puede explicarse en términos de las reglas de escalado (alométricas) anteriores e involucró cambios altamente especializados en la anatomía cerebral presumiblemente debido a la selección natural que expandió las regiones corticales sensoriales y motoras
14) procerebros de aves evolucionaron a lo largo de un camino muy diferente con expansión de su cresta ventricular dorsal (DVR), la región sensorimotora principal del telencéfalo aviar, muy similar en función al neocórtex de mamíferos, haciendo “muchas aves al menos tan inteligentes como la mayoría de los mamíferos”.
Striedter agrega una serie de puntos sobre el cerebro humano en un intento de identificar características que lo hacen especial en comparación con los cerebros de otros mamíferos.
15) En los seis millones de años transcurridos desde que los simios bípedos (homínidos) divergieron de otros simios, el tamaño absoluto del cerebro aumentó radicalmente (alrededor de cuatro veces), no gradualmente, sino en estallidos— desde que el género Homo evolucionó por primera vez, el tamaño absoluto del cerebro se duplicó de 400 a 800 centímetros cúbicos, luego permaneció relativamente estable en Homo erectus durante los siguientes 1.5 millones de años, pero luego volvió a explotar en la transición al Homo sapien hasta hace unos 100 mil años, momento en el que el tamaño cerebral absoluto alcanzó su valor actual de alrededor de 1,200 a 1,800 centímetros cúbicos. El primer salto en el tamaño del cerebro Homo probablemente estuvo relacionado con el cambio en la dieta que implica la transición a la carne y posteriormente la cocción de la carne. El segundo salto tal vez fue estimulado por la competencia entre humanos por compañeros y otros recursos;
16) el principio de “tarde es igual a grande” predice un neocórtex grande en humanos (la relación entre el neocórtex humano y la médula es el doble que la de los chimpancés);
17) el principio de “grandes iguales bien conectados” es consistente con el conocido número ampliado de proyecciones desde el neocórtex humano a las neuronas motoras en la médula (ubicada justo por encima de la médula espinal) y la médula espinal permitiendo una mayor precisión de control sobre los músculos que sirven las manos, los labios, la lengua, la cara, la mandíbula, los respiratorios músculos y cuerdas vocales, requeridos para el desarrollo del lenguaje humano hace unos 50 mil a 100 mil años;
18) una vez que apareció el lenguaje humano, se hicieron posibles cambios dramáticos en el comportamiento humano sin mayores aumentos en el tamaño del cerebro;
19) el aumento en el tamaño del cerebro tiene algunas desventajas: incluyendo el aumento de los costos metabólicos porque el cerebro utiliza tanta energía metabólica (20% de la energía metabólica humana a pesar de que es solo 2% del peso corporal humano; siendo tan metabólicamente caros los aumentos en el tamaño del cerebro deben ser pagados por una dieta mejorada o reducción de otras demandas de energía metabólica); disminución de la conectividad tal vez haciendo que los dos hemisferios sean más independientes entre sí, y quizás explicar por qué los dos hemisferios cerebrales se especializaron funcionalmente (realizando diferentes funciones cognitivas); y límites de tamaño del cerebro neonatal debido a limitaciones impuestas por el tamaño de la pelvis y canal de parto de la madre humana. Según Striedter, estos costos pueden explicar por qué el tamaño del cerebro humano se estabilizó hace unos 100 mil años;
20) dentro del neocórtex (que se encuentra solo en mamíferos), la corteza prefrontal lateral (ubicada hacia adelante de sus sienes) se ha agrandado relativamente en el cerebro humano, probablemente aumentando su papel en el comportamiento (ver Capítulo 14 para una discusión adicional de la corteza prefrontal lateral y funciones cognitivas superiores tales como pensamiento, planeación, etc.);
21) algunos cambios evolutivos clave en la estructura cerebral no fueron causados por aumentos en el tamaño cerebral absoluto o relativo, como la evolución del neocórtex de los mamíferos, sino que requieren una explicación adicional; comparar especies distantemente relacionadas con el tamaño cerebral absoluto por sí solo pierde factores importantes, por ejemplo, cerebros de algunas ballenas grandes pesan 5 veces más que el cerebro humano, pero los cerebros de ballenas tienen un neocórtex poco laminado y delgado; en casos de especies relacionadas distantemente, las comparaciones del tamaño cerebral relativo son más útiles, por ejemplo, las ballenas humanas y algunas dentas (por ejemplo, orcas) tienen tamaños cerebrales relativos significativamente mamíferos más grandes que el promedio de tamaño corporal similar;
22) dos hipótesis generales sobre la evolución cerebral son que los sistemas cerebrales individuales evolucionan independientemente por selección natural (la hipótesis del mosaico) o alternativamente que los componentes de tales sistemas evolucionan juntos debido a restricciones funcionales (la hipótesis concertada o de restricción), siendo una tercera visión que toda la evolución cerebral es simultáneamente a la vez mosaico y concertada.
Un problema con el enfoque de Striedter es que no explica las fuerzas de la selección natural que pueden dar cuenta de características más específicas de la evolución cerebral en especies específicas, incluida la nuestra. Admite que la evolución del neocórtex no puede explicarse por la evolución de cerebros más grandes y que el neocórtex evolucionó independientemente del tamaño cerebral absoluto. Sin embargo, además de reafirmar la teoría de que la vida social compleja en humanos ancestrales, junto con la competencia entre humanos por los recursos, estimuló la evolución de aumentos en el tamaño y complejidad del neocórtex, ofrece poca comprensión sobre el papel que jugó la selección natural en la evolución de la neocórtex de mamíferos, o en la evolución cerebral en general. Como afirma Adkins-Regan (2006, p.12-13) en su crítica a la obra de Striedter, “Hay relativamente poca discusión de pruebas de hipótesis sobre las presiones selectivas responsables del origen y mantenimiento de los rasgos. El autor parecería estar experimentando síntomas de incomodidad con el concepto de adaptación... Dado que los mecanismos cerebrales son productos de selección natural, una estrategia central en la neurociencia debería ser utilizar los métodos de la biología evolutiva, los cuales han tenido tanto éxito en ayudarnos a comprender los mecanismos y el diseño de los organismos en general”. En el capítulo 14 de este texto, sobre Inteligencia y Cognición, consistente con la crítica de Adkins-Regan, se encuentra una extensa discusión sobre el papel de la selección natural en la evolución de los sistemas cerebrales involucrados en la inteligencia y el pensamiento.
Encefalización del Cerebro
La encefalización del cerebro se refiere a un par de cosas: 1) el aumento del tamaño del cerebro a lo largo del tiempo y 2) el tamaño del cerebro en relación con la masa corporal total. La relación entre el tamaño del cerebro y la masa corporal no cambia tanto en los homínidos a pesar de los presuntos aumentos en la inteligencia, sin embargo, los aumentos en el tamaño cerebral se vuelven significativos comenzando con la especie Homo temprana. Los estudios de especies de mamíferos vivos sugieren que el aumento del número de neuronas y el aumento de la densidad de empaquetamiento de neuronas en la cavidad craneal pueden ser un factor clave en la evolución del aumento de la inteligencia entre las especies. Sin embargo, si bien hay un aumento gradual en el tamaño del cerebro en todo el linaje australopitecino, no es hasta principios del Homo que hay un aumento significativo en la capacidad craneal, aproximadamente un aumento de 20% sobre los australopitecinos. Más significativo es el aumento de aproximadamente 50% en el tamaño cerebral de Homo erectus y las especies de Homo anteriores. No es solo el tamaño del cerebro lo que es importante. Durante este proceso de encefalización, se ha especulado, sin evidencia fósil, que también hubo un recableado del cerebro que coincide con el surgimiento de la cultura material como las herramientas de piedra. No es hasta que esto ocurre que los homínidos abandonan África, posibilitada en gran medida por los avances culturales.
Los cerebros de primates no humanos son simétricos al igual que los de los primeros homínidos. Con la aparición del Homo vemos la lateralización del cerebro —se vuelve asimétrico (cerebro derecho, cerebro izquierdo). Esto lo sabemos por los endocasts. Los endocastos se forman cuando los minerales reemplazan la materia cerebral dentro del cráneo durante el proceso de fosilización. Estos endocastos permiten a los investigadores estudiar los pliegues corticales del cerebro y compararlo con los humanos modernos. A partir de los endocasts, los investigadores determinaron que tres áreas del cerebro comenzaron a cambiar en Homo: el cerebelo, que maneja las actividades motoras aprendidas, el sistema límbico, que procesa la motivación, la emoción y la comunicación social, y el corteza cerebral, que es responsable de experiencias sensoriales, memoria y funciones mentales complejas como el lenguaje, la cognición, la planificación, la imaginación y la inteligencia (véanse los capítulos 14 y 15 sobre Cognición e Inteligencia y Lenguaje). Son estos cambios los que pudieron haber permitido a los primeros miembros de nuestro género, Homo, desarrollar adaptaciones culturales a las presiones ambientales.
Figura\(\PageIndex{8}\): Comparación de capacidades craneales de primates vivos. Cráneos de primates proporcionados por cortesía del Museo de Zoología Comparada de la Universidad de Harvard. (Imagen de Wikimedia Commons; Archivo:Serie de cráneo de primate con legend.png; https://commons.wikimedia.org/wiki/F...ith_legend.png; por Christopher Walsh, Escuela de Medicina de Harvard; licenciado bajo la licencia Creative CommonsReconocimiento-Compartir Igual 2.5 Genérica).
¿Por qué cambió el cerebro a principios del Homo?
La pregunta que enfrentaron los investigadores fue por qué cambió el cerebro. Los cerebros grandes tienen algunas desventajas:
requieren mucha energía metabólica; aproximadamente 25-30% de la energía metabólica de un ser humano es consumida por el cerebro aunque comprende solo 2% del peso corporal total
los cerebros grandes requieren que los bebés nazcan en un estado inmaduro con la cabeza y el tamaño del cerebro lo suficientemente pequeños como para permitir el parto a través de un canal de parto relativamente pequeño, lo que resulta en un período más largo de dependencia infantil (el cerebro infantil promedio al nacer es solo aproximadamente 1/3 del tamaño de un cerebro adulto con gran parte del desarrollo cerebral y el desarrollo conductual y mental ocurriendo durante un período de años después del nacimiento)
la dependencia infantil más prolongada es un mayor drenaje de la energía materna; la madre debe tener una nutrición adecuada no solo para ella sino para el lactante lactante
se ha sugerido que los cerebros más grandes disminuyen la eficiencia bípeda de las hembras porque deben tener una pelvis y canal de parto más anchos para dar a luz a un lactante de cerebro grande
Entonces, para que los cerebros grandes se fijaran evolutivamente en la población Homo, las ventajas tuvieron que superar las desventajas enumeradas anteriormente.
Una posible explicación incorpora la interacción de tres variables diferentes: tamaño del grupo, patrones de subsistencia complejos (como forrajeo para alimentos o domesticación de animales y plantas) y el valor nutricional de la carne (Campbell y Loy 2000:318). Primero abordemos el tamaño del grupo.
La investigación sugiere que el tamaño del cerebro y el tamaño de los grupos sociales se correlacionan positivamente entre los primates vivos, lo que implica que los cerebros grandes ayudaron a los individuos a realizar un seguimiento de información social compleja como jerarquías de dominancia, alianzas, enemigos, etc.
Segundo, un cerebro grande permite a los primates realizar un seguimiento de los grandes territorios de subsistencia y permite a los omnívoros desarrollar estrategias para recolectar una amplia variedad de alimentos.
Tercero, comer carne es una manera relativamente fácil de obtener la nutrición necesaria para ejecutar un cerebro grande, que, como se mencionó anteriormente, en los humanos modernos toma alrededor de 1/4 a 1/3 de nuestra energía metabólica diaria. Sin embargo, la carne cruda requiere mucha energía para digerir, por lo que la invención de cocinar carne a fuego puede haber jugado un papel en la evolución de cerebros más grandes porque la carne cocida es mucho más fácil, y requiere menos energía metabólica, para digerir. Así, la cocción de carne proporcionó una rica fuente de energía metabólica, apoyando la evolución de cerebros más grandes.
El argumento a favor de la hipótesis del cerebro social es presentado por Robin Dunbar (1998). Dunbar también afirma que fueron los cambios en el neocórtex, la capa superior de 2-4 mm de espesor de los hemisferios cerebrales, los que fueron críticos en la “homininzación” (desarrollo de habilidades cognitivas humanas) de nuestros antepasados (ver capítulo 14 para obtener ventajas adaptativas adicionales asociadas con la encefalización y aumentos en inteligencia).
Una perspectiva relacionada sobre la evolución del cerebro humano enfatiza el papel de la cultura en el aumento del tamaño del cerebro. Como afirma Geertz (2013, p. 180-181):
El cerebro australopitecino tenía aproximadamente el mismo tamaño que el de los chimpancés modernos (unos 400— 600 cm 3). El cerebro del Homo sapiens mide aproximadamente 1200—1700 cm 3. Pero la expansión del cerebro había comenzado ya con la aparición del Homo habilis, la primera especie homínea que salió de la línea Australopitecina hace unos 2.5 millones de años (con un tamaño cerebral de 500-800 cm 3) y se volvió aún más espectacular con la aparición de hace 1.5 millones de años del artista de herramientas de piedra Homo erectus cuyo cerebro se acercó en tamaño a los humanos modernos a 750—1250 cm 3. Hoy es asumido por arqueólogos y paleontólogos que la producción y uso de herramientas fue incremental a la expansión del cerebro. Debido a que el uso de herramientas es mucho más que simplemente golpear piedras y huesos juntos, y de hecho, depende mucho de las recetas, reglas e instrucciones de los patrones culturales, hoy en día es ampliamente aceptado que la cultura impulsó la expansión del cerebro (junto con otras cosas, por supuesto, como comer carne). La cadena causal, una vez más, sin embargo, es la cultura primero, la expansión cerebral después. 'Debido a que la fabricación de herramientas le da prioridad a la habilidad manual y a la previsión, su introducción debe haber actuado para cambiar las presiones de selección para favorecer el rápido crecimiento del prosencéfalo como, con toda probabilidad, lo hicieron los avances en la organización social, la comunicación y la regulación moral que hay razón para creen que también ocurrieron durante este periodo de solapamiento entre el cambio cultural y biológico. Tampoco fueron tales cambios del sistema nervioso meramente cuantitativos; las alteraciones en las interconexiones entre las neuronas y su forma de funcionamiento pueden haber sido de mayor importancia que el simple aumento de su número”.
El prosencéfalo incluye la corteza cerebral (lóbulos frontal, parietal, temporal y occipital) y una serie de estructuras subcorticales como el tálamo, el hipotálamo, el sistema límbico y los ganglios basales. Diferentes presiones de selección derivadas de la fabricación y uso de herramientas, organización social compleja, comunicación, regulación moral y otros factores sociales y culturales probablemente habrían afectado diferentes áreas del prosencéfalo, e incluso diferentes lóbulos de la corteza cerebral, de diferentes maneras. Esto se deduce del hecho de la organización cerebral que diferentes circuitos y redes cerebrales están especializados para diferentes funciones cognitivas y conductuales (ver Cosmides & Tooby, 2002).
Geología y antecedentes ambientales
El periodo Mioceno (hace aproximadamente 23-5 millones de años) estuvo geológicamente activo en África. Este es el período de la radiación adaptativa de los simios y un período de construcción montañosa que condujo a la formación del Gran Valle del Rift de África. Con el surgimiento de las montañas de la grieta, las lluvias que hasta ahora se habían movido a través del continente desde el Océano Atlántico quedaron bloqueadas (denominadas sombra de lluvia), lo que llevó a la aridificación de África Oriental. El ambiente de sabana que evolucionó en África Oriental fue y es un ambiente mucho más abierto que el entorno boscoso de África Occidental y Central, lo que llevó al surgimiento de nuevas adaptaciones para plantas y animales. Es en este entorno recién emergente donde despega la evolución de los homínidos.
Los datos paleoclimáticos se han correlacionado con eventos de especiación en la evolución de los homínidos, pero no parece dar cuenta de todos los eventos de especiación. Sin embargo, los datos paleoclimáticos sugieren lo siguiente:
Los pastizales se extendieron en África entre 10-5 millones de años atrás durante una fase de enfriamiento y secado. Es durante este marco de tiempo que vivió el ancestro común de los simios africanos y los humanos. El ancestro común se parecía más a un cuadrúpedo que era arbóreo o al menos pasaba una cantidad significativa de tiempo en los árboles. A mediados de este periodo, hace aproximadamente 7-6 millones de años, surgió el primer homínido bípedo, y algunos otros homínidos tempranos son referidos como proto-homínidos en reconocimiento a sus rasgos primitivos, parecidos a simios.
En el periodo Plioceno medio, hace 3-2 millones de años, otra fase de enfriamiento y secado se correlaciona con la radiación adaptativa de los homínidos, incluyendo la aparición de los australopitecinos robustos del género Homo, el mismo género que los humanos modernos.
Cerca del inicio del periodo Pleistoceno, también conocido como la Edad de Hielo, el ambiente siguió secándose. Los hábitats abiertos se propagan en África Oriental. Durante este periodo, el Homo ergaster (Homo erectus) emerge y finalmente abandona el continente africano.
Estos datos tienden a hacernos pensar que la evolución de los homínidos fue impulsada por cambios ambientales; sin embargo, los cambios geológicos, climáticos y ambientales en África durante el Mioceno, Plioceno y Pleistoceno pueden haber tenido poco que ver con la evolución de los homínidos, dejando abierta la posibilidad de que el hipótesis discutidas anteriormente, u otras aún por proponer, pueden estar más causalmente relacionadas con la evolución de los homínidos.
Transiciones clave en la evolución humana
En el camino evolutivo hacia los humanos modernos, algunos científicos han identificado varias “transiciones clave: (1) simio africano a mono bípedo terrestre (alrededor de 4 Ma) [MA=Hace millones de años]; (2) simio bípedo terrestre (australopitecino) a 'Homo temprano' (alrededor de 2 Ma); (3) Homo temprano [especie] a Homo heidelbergensis (1—0. 8Ma); (4) Homo heidelbergensis a Homo de cerebro más grande (de 500 ka) [ka=hace mil años], y (5) Homo de cerebro más grande a H. sapiens (de 200 ka). [Respecto al comportamiento social,] “entre los modelos de base arqueológica, existe un total acuerdo en que el surgimiento de la socialidad homínido basal, como parte de nuestra herencia de primates, debe atribuirse a las primeras etapas evolutivas, posiblemente antes de la emergencia del género Homo” (Anghelinu, 2013, p.13).
Para una mayor exploración
Explora la evolución humana en la impresión
Boyd, Robert y Joan B. Silk. 2009. Cómo evolucionaron los humanos, 5ª edición. Nueva York: W. W. Norton.
Campbell, Bernard G. y James D. Loy. 2000. Humankind Emerging, 8ª edición. Boston: Allyn & Bacon.
Johanson, Donald y Kate Wong. 2010. El legado de Lucy: La búsqueda de los orígenes humanos. Nueva York: Harmony Books.
Stringer, Chris y Peter Andrew. 2006. El mundo completo de la evolución humana. Nueva York: Támesis y Hudson.
Tattersall, Ian. 2008. El rastro fósil: cómo sabemos lo que pensamos que sabemos sobre la evolución humana. Nueva York: Oxford University Press.
Explora la Evolución Humana en la Web
Convertirse en humanos
Charla: Origins Fossil Homínidos
Salón de los Orígenes Humanos
Diario de la Ciencia: Noticias de Evolución Humana
Redescubriendo Biología:Unidad 9 Evolución Humana
BBC: La evolución del hombre
Evolución humana: La evidencia fósil en 3D
Proto-homínidos
El homínido más antiguo descubierto hasta la fecha es de entre 7.2 a 6.8 mya (hace millones de años).
Figura\(\PageIndex{9}\): Sahelanthropus tchadensis. El homínido más antiguo conocido.
El cráneo es una combinación de rasgos parecidos a simios y humanos. Las características similares a los simios incluyen el tamaño del cerebro, la cresta pesada de la frente Sus características humanas incluyen la posición delantera del foramen magnum, los dientes caninos más pequeños y el grosor intermedio del esmalte premolar y molar. Debido a la pronunciada cresta de la ceja, el equipo de Brunet sugiere que el espécimen es masculino.
Existe un debate entre los investigadores sobre si este fósil es un homínido o un simio. Algunos sugieren que el espécimen pertenece al de un simio hembra porque es probable que encuentre caninos usados en las puntas en simios hembra.
homínidos
Si bien los homínidos se presentarán más o menos en orden cronológico, no confundan el orden cronológico con las relaciones evolutivas lineales; algunos homínidos que se presentan no están en la línea directa con los humanos modernos. También es importante tener en cuenta que cada año se hacen nuevos descubrimientos que refinan lo que sabemos sobre la evolución humana.
Descubierto en 1974 en Hadar, Etiopía, Australopithecus afarensis es posiblemente la especie de homínidos fósiles más conocida. Está fechado de 3.7-3.0 mya (Scarre 2014). Se recuperó más del 40% del esqueleto, lo que permitió al equipo reconstruir completamente el esqueleto. Este espécimen fósil, llamado Lucy, aunado a las huellas encontradas en Laetoli, Tanzania, en 1978 por Mary Leakey, confirmó que Au. afarensis era completamente bípedo, aunque no exactamente como los humanos modernos. Las huellas en Laetoli indican que Au. afarensis tuvo un paso corto y una marcha paseante. Desde la década de 1970, se han encontrado cientos de especímenes de Au. afarensis (¡60 individuos al menos solo de Hadar!) en Etiopía, Kenia y Tanzania, permitiendo a los paleoantropólogos hacer “declaraciones definitivas sobre el patrón locomotor y la estatura de” (Jurmain 2013:211) este homínido temprano.
Figura\(\PageIndex{10}\): Reconstrucción del esqueleto fósil de “Lucy” el Australopithecus afarensis
Au. afarensis tiene varias características primitivas o similares a simios, incluyendo un cerebro relativamente pequeño en comparación con Homo, una arcada dental en forma de U, una nariz plana, una frente aplanada (conocida como platicefalia) y una cara prognatica (caracterizada por una mandíbula inferior sobresaliente). Sus caninos, si bien son más grandes que el Homo, son más pequeños que los homínidos anteriores. Si bien su cerebro era más grande que los homínidos anteriores, todavía es pequeño en comparación con el género Homo. Hay evidencia de dimorfismo sexual; los machos Au. afarensis no eran más altos de 5 pies y las hembras alrededor de 3-4 pies, similares en proporción a los simios africanos modernos. Au. afarensis tiene un conjunto de rasgos derivados asociados con el bipedalismo; es decir, pelvis en forma de cuenco, curva en S de la columna vertebral y anatomía de la rodilla. Sin embargo, la curvatura de los dedos de manos y pies y la proporción de brazos a piernas sugieren a algunos investigadores que Au. afarensis pasó algún tiempo en los árboles.
En 2006, un niño Au. afarensis de 3.3 millones de años fue descubierto a menos de 4 km de donde se encontró a Lucy en 1974. No sólo es el fósil juvenil más antiguo jamás encontrado, también es el fósil de homínidos más completo encontrado hasta la fecha. Selam, como el fósil fue apodado (o el bebé de Lucy o el bebé de Dikika), confirmó sugerencias anteriores de que Au. afarensis era bípedo, pero pasaba tiempo en los árboles. La estructura del hombro con sus articulaciones de hombro apuntando hacia arriba y la cresta ósea que corre a lo largo de los omóplatos son como la de los simios, lo que habría facilitado el movimiento arbóreo aunque no fueran tan capaces como los chimpancés de moverse en los árboles.
Los datos paleoecológicos indican que Au. afarensis vivió tanto en pastizales (sabanas) como en entornos boscosos.
Australopithecus africanus (ver Figura 3.7.2, anterior) ha sido fechado en 3-2 mya (Scarre 2014). En comparación con Au. afarensis, Au. africanus tiene incisivos más pequeños y molares más grandes; los caninos ya no tienen el aspecto puntiagudo y triangular visto en simios y Au. afarensis; sin embargo, Au. africanus aún exhibe alguna protrusión de la mandíbula inferior y tiene un cerebro pequeño como el de Au. afarensis. Existe un consenso general de que Au. africanus es descendiente directo de Au. afarensis. Su relación con el Homo es menos clara.
Australopithecus garhi
Encontrado en Bouri, Etiopía en 1997, Au. garhi (ver Figura 3.7.2, anterior) data de 2.5 mya. Se han encontrado pocos especímenes fósiles y los que tienen son relativamente fragmentarios. Se encontraron un cráneo y otros fragmentos de cráneo que sirven como base para la identificación de especies. El tamaño y la forma de sus dientes molares sugieren a algunos investigadores que Au. garhi está relacionado con Paranthropus aethiopicus (ver abajo), pero sus otras características, por ejemplo, el cerebro, la cara y otros dientes, se parecen más al género Homo. Ante esto, algunos investigadores sostienen que es ancestral al Homo.
Figura\(\PageIndex{12}\): Australopithecus garhi.
Australopithecus sediba
Australopithecus sediba (Figura 3.3.13) fue encontrado en 2008 en la cueva de Malapa, Sudáfrica, que data de 1.95-1.75 mya, tiene un mosaico de características que sugieren que puede ser transicional de los australopitos al género Homo. Sin embargo, esta afirmación es polémica ya que las fechas más tempranas para Homo son anteriores a Au. sediba en unos 500,000 años (Becoming Human 2008).
Las características que vinculan a Au. sediba con Homo incluyen la forma de la pelvis, el caso cerebral más vertical, los pómulos más pequeños y la forma molar.
Australopitos Robustos
Tres especies robustas de homínidos surgieron en el periodo Plio-Pleistoceno: Paranthropus aethiopicus, Paranthropus boisei y Paranthropus robustus (ver Figuras 3.3.14-17). Tienen características morfológicas que sugieren que estaban bien adaptados para comer alimentos duros que necesitaban molienda, lo que llevó a que fueran identificados como “robustos”.
Nota
Existe debate sobre si las diferencias mencionadas anteriormente califican a los australopitos robustos para estar en un género separado de los australopitecinos. En algunas obras antropológicas se utiliza el género Australopithecus. En otras, como esta obra, se utiliza Paranthropus.
No hay evidencia que sugiera que P. boisei sea ancestral de cualquier homínido posterior.
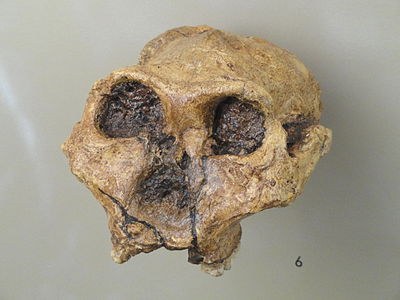
Figura\(\PageIndex{16}\). Paranthropus boisei; modelo de macho adulto (Museo Smithsonian de Historia Natural).
Figura\(\PageIndex{17}\). Paranthropus robustus.
Homo Género
Arriba describimos a nuestros ancestros humanos más antiguos, principalmente miembros del género Australopithecus que vivieron entre 2 millones y 4 millones de años atrás. Aquí presentamos los primeros miembros del género Homo, enfocándose en las especies Homo habilis y Homo erectus (ver Figura 3.3.19 y Figura 3.3.20, a continuación).
Definiendo el género Homo
Al agrupar especies en un género común, los biólogos considerarán criterios como las características físicas (morfología), evidencia de ascendencia común reciente y estrategia adaptativa (uso del ambiente). Sin embargo, existe desacuerdo sobre cuál de esos criterios debe priorizarse, así como cómo deben interpretarse fósiles específicos a la luz de los criterios. Existe acuerdo general en que las especies clasificadas como Homo deben compartir características ampliamente similares a las de nuestra especie. Estos incluyen los siguientes:
• un tamaño cerebral relativamente grande, lo que indica un alto grado de inteligencia;
• una cara más pequeña y plana;
• mandíbulas y dientes más pequeños; y
• mayor dependencia de la cultura, particularmente el uso de herramientas de piedra, para explotar una mayor diversidad de ambientes (zona adaptativa).
Algunos investigadores incluirían un tamaño corporal general más grande y proporciones de extremidades (piernas más largas/brazos más cortos) en esta lista. También hay una aparente disminución en el dimorfismo sexual (diferencias de tamaño corporal entre hombres y mujeres). Si bien estos criterios parecen relativamente claros, evaluarlos en el registro fósil ha resultado más difícil, particularmente para los primeros miembros del género. Hay varias razones para ello. Primero, muchos especímenes fósiles que datan de este periodo de tiempo están incompletos y mal conservados, lo que dificulta su evaluación. Segundo, los primeros fósiles de Homo aparecen bastante variables en el tamaño del cerebro, los rasgos faciales y el tamaño de los dientes y el cuerpo, y aún no hay consenso sobre cómo darle mejor sentido a esta diversidad.
En esta sección, tomaremos varias vías para examinar el origen y evolución del género Homo. Primero, exploraremos las condiciones ambientales de la época del Pleistoceno en la que evolucionó el género Homo. A continuación examinaremos la evidencia fósil de las dos especies principales tradicionalmente identificadas como Homo temprano: Homo habilis y Homo erectus. Luego utilizaremos datos de fósiles y sitios arqueológicos para reconstruir el comportamiento de los primeros miembros del Homo, incluyendo la fabricación de herramientas, prácticas de subsistencia, patrones migratorios y estructura social. Finalmente, los consideraremos juntos en un intento de caracterizar las estrategias adaptativas clave del Homo temprano y cómo pusieron a nuestros primeros antepasados en la trayectoria que condujo a nuestra propia especie, el Homo sapiens.
Clima
Las primeras especies de homínidos cubiertas anteriormente, como Ardipithecus ramidus y Australopithecus afarensis, evolucionaron durante la época tardía del Plioceno. El Plioceno (hace 5.3 millones a 2.6 millones de años) estuvo marcado por condiciones más frías y secas, con casquetes de hielo formándose permanentemente en los polos. Aún así, el clima de la Tierra durante el Plioceno era considerablemente más cálido y húmedo que en la actualidad.
La posterior época del Pleistoceno (2.6 millones de años a 11.000 años atrás) marcó el comienzo de un importante cambio ambiental. El Pleistoceno se conoce popularmente como la Edad de Hielo. Dado que el término “Edad de Hielo” tiende a evocar imágenes de glaciares y mamuts lanudos, naturalmente se asumiría que se trataba de un período de clima uniformemente frío alrededor del globo. Pero en realidad no es así. En cambio, el clima se volvió mucho más variable, ciclando abruptamente entre ciclos cálido/húmedo (interglacial) y frío/seco (glacial).
En África, la investigación paleoclimática ha determinado que los pastizales se expandieron y encogieron varias veces durante este período, incluso cuando se expandieron a largo plazo, volviéndose cada vez más comunes durante el Pleistoceno. Una solución adoptada por algunos homínidos fue especializarse en alimentarse de los nuevos tipos de plantas que crecen en este paisaje. Los robustos australopitecinos probablemente desarrollaron sus dientes molares grandes con esmalte grueso para explotar este nicho dietético particular.
Miembros del género Homo tomaron una ruta diferente. Ante el inestable clima africano y el paisaje cambiante, desarrollaron cerebros más grandes que les permitieron confiar en soluciones culturales como la elaboración de herramientas de piedra que abrieron nuevas oportunidades de búsqueda de alimento. Esta estrategia de flexibilidad conductual les sirvió bien durante este tiempo impredecible y condujo a nuevas innovaciones como el aumento del consumo de carne, la caza cooperativa y la explotación de nuevos entornos fuera de África, incluyendo Europa y Asia.
El surgimiento del Homo (nuestro género)
La aparición del género Homo marca el advenimiento de cerebros más grandes, el surgimiento de la cultura material (al menos la cultura material que sobrevive en el registro arqueológico) y la eventual colonización del mundo fuera de África. Las primeras especies de Homo son contemporáneas con varios australopitos: Au. africanus, Au. garhi, Au. sediba y todas las especies de Paranthropus. África estaba al ras de homínidos (Figura\(\PageIndex{16}\)). Hay varias tendencias que vemos en la evolución de las primeras especies de Homo a humanos, Homo sapiens:
Redondeo del cráneo
Agrandamiento y recableado del cerebro (juzgado a partir de los endocastos)
Caras y dientes más pequeños
Prognatismo decreciente
Tallness
Diversidad de rasgos culturales
Al igual que con la visión general de los homínidos tempranos, continuaremos tomando el enfoque “grumper”, una tendencia a agrupar especímenes en menos especies en lugar de tender a diferenciar e hipótesis de un mayor número de especies. Varias especies de Homo no serán discutidas en detalle, pero pueden mencionarse de pasada, por ejemplo, Homo rudolfensis, Homo ergaster, Homo gautengensis, Homo antecessor, Homo cepranensis, Homo rhodesiensis, Homo tsaichangensis. Primero examinaremos las características morfológicas de diversas especies de Homo. Los rasgos culturales se abordarán por separado.
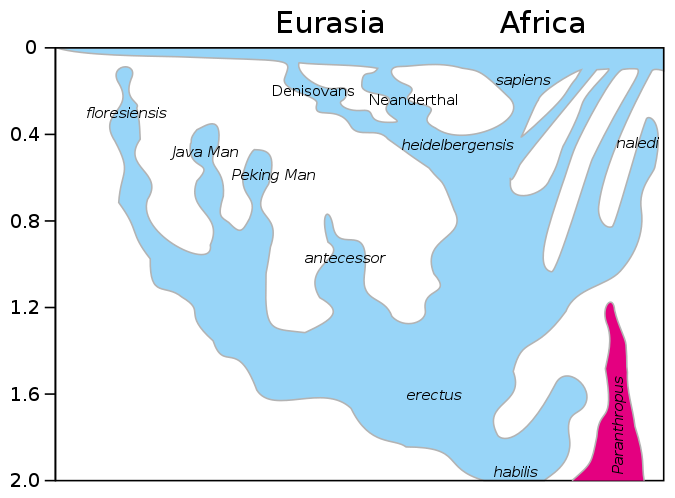
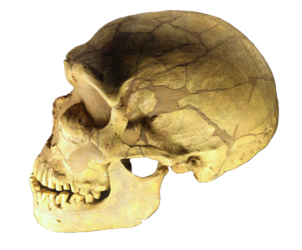
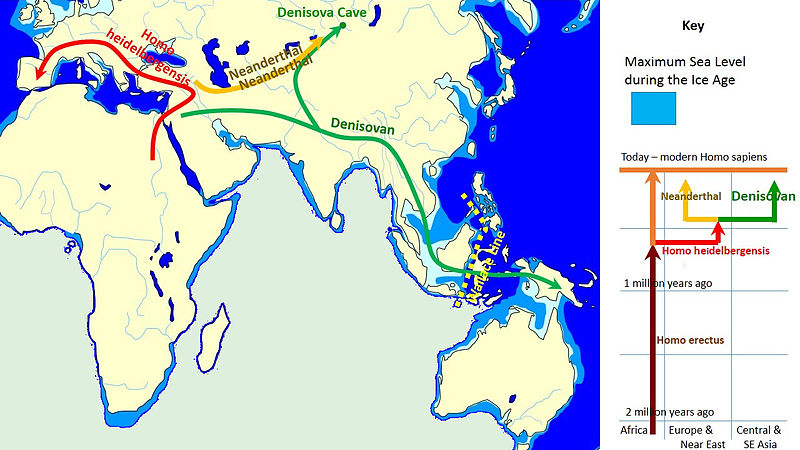
Figura\(\PageIndex{18}\): Un modelo de la evolución del género Homo en los últimos 2 millones de años (millones de años, Mya, está en el eje vertical). La rápida expansión “Fuera de África” de H. sapiens se indica en la parte superior del diagrama por la expansión lateral del azul a lo largo de la parte superior del diagrama, con mezcla indicada con neandertales, denisovanos y homínidos africanos arcaicos no especificados. La supervivencia tardía de australopitecinos robustos (Paranthropus) junto al Homo hasta 1.2 Mya se indica en púrpura.
H. heidelbergensis se muestra como el vínculo entre neandertales, denosiovanos y H. sapiens
División de H. erectus asiático en Java Man y Peking Man
H. antecessor mostrado como una rama de H. erectus llegando a Europa
Después de que H. sapiens emergieron de África unos 60 kya se extendieron por todo el mundo y se cruzaron con otros descendientes de H. heidelbergensis, y neandertales, denisovanos
Homo Habilis: Los primeros miembros de nuestro género
El Homo habilis ha sido tradicionalmente considerado como la especie más temprana colocada en el género Homo. Sin embargo, como veremos, existe un desacuerdo sustancial entre los paleoantropólogos sobre los fósiles clasificados como Homo habilis, incluyendo si provienen de una sola o múltiples especies, o incluso si deberían formar parte del género Homo en absoluto.
En comparación con los australopitecinos, el Homo habilis tiene un tamaño cerebral algo mayor, un promedio de 650 centímetros cúbicos (cc) en comparación con menos de 500 cc para Australopithecus. Adicionalmente, el cráneo es más redondeado y la cara menos prognatica. Sin embargo, los restos postcraneales muestran un tamaño corporal y proporciones similares a Australopithecus. Las fechas conocidas para los fósiles identificados como Homo habilis van desde hace aproximadamente 2.5 millones de años hasta hace 1.7 millones de años.
Tamaño del Cerebro: 650 cc promedio (rango de 510 cc a 775 cc)
Dentición: Dientes más pequeños con esmalte más delgado comparado con Australopithecus; forma de arcada dental parabólica
Características craneales: cráneo más redondo y menos prognatismo facial que Australopithecus
Características postcraneales: Estatura pequeña; plano corporal similar al Australopithecus
Cultura: Herramientas Oldowan; el más antiguo del tipo conocido de herramienta de piedra; raspadores, choppers; indicación de habilidades cognitivas cambiantes (ver Módulo 3.8)
Es casi seguro que las herramientas de piedra fueron anteriores al Homo habilis (posiblemente por Australopithecus garhi o la especie responsable de las herramientas de Kenia que datan de hace 3.7 millones de años). Sin embargo, las herramientas de piedra se vuelven más frecuentes en sitios que datan de hace aproximadamente 2 millones de años, la época del Homo habilis (Roche, Blumenschine y Shea 2009). Esto sugiere que estos homínidos dependían cada vez más de herramientas de piedra para ganarse la vida.
A las herramientas de piedra se les asigna mucha importancia en el estudio de los orígenes humanos. El estudio de la forma de las herramientas, las materias primas seleccionadas y la forma en que fueron elaboradas y utilizadas puede proporcionar información sobre los procesos de pensamiento de los primeros humanos y cómo modificaron su entorno para sobrevivir (ver Contenido Complementario, Capítulo 18, Cultura material). Los paleoantropólogos tradicionalmente han clasificado colecciones de herramientas de piedra en industrias, en función de su forma y modo de fabricación. No existe una correspondencia exacta entre una industria de herramientas y una especie de homínidos; sin embargo, se pueden hacer algunas asociaciones generales entre industrias de herramientas y homínidos particulares, ubicaciones y períodos de tiempo. Los nombres de las cuatro principales industrias de herramientas en la evolución humana (de la más antigua a la más reciente) son: Oldowan, Acheulean, Mousterian y Upper Paleolítico. La industria de herramientas de piedra más antigua es Oldowan, que lleva el nombre del sitio de Olduvai Gorge donde se descubrieron las herramientas por primera vez. Generalmente se considera que el periodo de tiempo del Oldowan dura de aproximadamente 2.5 mya a 1.6 mya. Las herramientas de esta industria se describen como herramientas de “escamas y picadoras”, las picadoras que consisten en adoquines de piedra con algunas escamas las golpearon. Para un observador casual, es posible que estas herramientas no se vean muy diferentes de las rocas rotas al azar. Sin embargo, son más difíciles de hacer de lo que sugiere su apariencia cruda. La roca seleccionada como núcleo debe ser golpeada por la roca que sirve como piedra de martillo en el ángulo recto para que se eliminen una o más escamas planas. Esto requiere seleccionar rocas que se fracturarán de manera predecible en lugar de fragmentarse, así como la capacidad de planificar con anticipación e imaginar los pasos necesarios para crear el producto terminado. El proceso deja tanto el núcleo como las escamas con filos cortantes afilados que pueden ser utilizados para una variedad de propósitos.
¿Qué hacían los homínidos con las herramientas? Una actividad clave parece haber sido la matanza de animales. Los huesos de animales con marcas de corte comienzan a aparecer en sitios con herramientas Oldowan. Los estudios de huesos animales en el sitio muestran que los huesos de las piernas a menudo están agrietados, lo que sugiere que estaban extrayendo la médula de las cavidades óseas. Es interesante considerar si los homínidos cazaban a estos animales o los adquirieron por otros medios. Los huesos masacrados provienen de una variedad de mamíferos africanos, ¡desde pequeños antílopes hasta animales tan grandes como ñus y elefantes! Es difícil imaginar el Homo habilis lento y de cuerpo pequeño con sus herramientas Oldowan que derriba animales tan grandes. Una posibilidad es que los homínidos estuvieran secuestrando cadáveres de leones y otros gatos grandes. Independientemente de cómo estuvieran adquiriendo la carne, todas estas actividades sugieren un importante cambio dietético de la forma en que comían los australopitecinos. Los fabricantes de herramientas Oldowan estaban explotando un nuevo nicho ecológico que les proporcionaba más proteínas y calorías. En general, el uso creciente de herramientas de piedra permitió a los homínidos ampliar su nicho ecológico y ejercer un mayor control sobre su entorno. Como veremos en breve, este patrón continuó y se hizo más pronunciado con Homo erectus.
Descubrimiento de Homo habilis
Homo habilis fue descubierto por primera vez por Louis y Mary Leakey en Olduvai Gorge, Tanzania, en 1960. Asociados con herramientas de piedra (Oldowan), los Leakeys nombraron a su descubrimiento “hombre práctico”. Se han encontrado fósiles de H. habilis en Tanzania, Kenia, Etiopía y Sudáfrica, aunque existe cierto debate sobre si los especímenes de Sudáfrica deben incluirse en la especie. Algunos investigadores sostienen que hubo otra especie de Homo temprana, Homo rudolfensis, que data de 2.4-2.5 mya. Los fósiles de H. rudolfensis son ligeramente más grandes que los de H. habilis, lo que lleva a algunos investigadores a sugerir que H. habilis exhibió dimorfismo sexual y lo que estamos viendo son especímenes masculinos y femeninos de H. habilis. Otros afirman que las diferencias de tamaño son lo suficientemente significativas como para garantizar las dos designaciones de especies (O'Neil 1999-2012). En 2013 se descubrió una mandíbula Homo en el área de investigación Ledi-Geraru, Afar, Etiopía. Con una fecha de 2.8 a 2.75mya, la mandíbula presenta una barbilla similar a Australopithecus y dientes similares a Homo (Villmoare et al. 2015). Si bien aún es temprano en el proceso de investigación, este descubrimiento y otras investigaciones pueden retrasar la fecha del origen del Homo y ayudar a resolver el debate entre los fósiles de H. rudolfensis y H. habilis. Para nuestro propósito, los consideraremos todos H. habilis, haciendo el rango de fechas aproximado para este homínido 2.5 a 1.4mya o menos conservadoramente a 1.7mya.
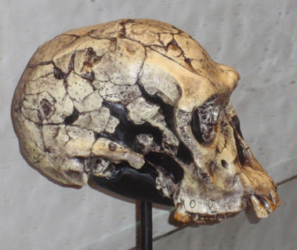
Figura\(\PageIndex{19}\): Homo habilis.
Morfológicamente, H. habilis tiene un cerebro más grande que los australopitos, aproximadamente 35% más grande (O'Neil c1999-2012). Recordarás de la sección sobre tendencias en la evolución humana que se especula que el cerebro también comenzó a recablear en este punto. H. erectus presenta menos prognatismo (protrusión de la mandíbula inferior) y platicefalia (aplanamiento de la parte posterior de la cabeza) que los homínidos tempranos. La cresta de la ceja también es más pequeña. Todos estos rasgos juntos hacen que el rostro sea más pequeño que los australopitos. Postcranealmente, H. habilis exhibe una mezcla de rasgos primitivos y derivados. Los rasgos primitivos que lo conectan con un antepasado australopital son los antebrazos más largos y el tamaño de los huesos de los dedos junto con la forma en que los tendones se unen a los huesos de la muñeca. Las puntas de los huesos de los dedos son anchas como los humanos. Los dientes más pequeños, una arcada dental en forma de arco parabólico, la morfología del pie y un cráneo más redondeado completan los rasgos humanos. El microanálisis del desgaste dental indica que H. habilis fue omnívoro.
Cuadro 3.3.2
Localizaciones clave de los fósiles de Homo habilis y los correspondientes fósiles y fechas(adaptado por Kenneth A. Koenigshofer, PhD, Chaffey College, de Exploraciones: Una invitación abierta a la antropología biológica;https://explorations.americananthro.....php/chapters/; Capítulo 10, “Primeros miembros del género Homo” de Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, y Lara Braff, Eds.; licenciado bajo una Licencia Creative Commons Reconocimiento-NoComercial 4.0 Internacional, excepto donde se indique otra cosa).
Ubicación de Fósiles Fechas (mya = hace millones de años) Descripción
Ledi-Gararu, Etiopía 2.8 mya Mandíbula inferior parcial con evidencia de rasgos tanto de Australopithecus como de Homo; tentativamente considerada evidencia fósil de Homo Temprano más antigua.
Garganta de Olduvai, Tanzania 1.7 mya a 1.8 mya. Varios especímenes diferentes se clasificaron como Homo habilis, incluyendo el espécimen tipo encontrado por Leakey, un pie relativamente completo, y un cráneo con una capacidad craneal de aproximadamente 600 cc.
Koobi Fora, Cuenca del lago Turkana, Kenia 1.9 mya. Varios fósiles de la cuenca del lago Turkana muestran considerables diferencias de tamaño, lo que lleva a algunos expertos a clasificar al espécimen más grande como una especie separada, Homo rudolfensis.
Sterkfontein y otras posibles cuevas sudafricanas de aproximadamente 1.7 mya han arrojado restos fragmentarios identificados como Homo habilis, pero los sitios de cuevas sudafricanas aseguran fechas y faltan detalles sobre los fósiles.
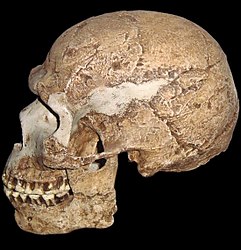
Homo erectus
Alrededor de 1.9 mya, apareció una nueva especie de Homo. Conocido como Homo erectus, la visión científica predominante era que esta especie se parecía mucho más a nosotros. Estos homínidos estaban equipados con cerebros más grandes y cuerpos grandes con proporciones de extremidades similares a las nuestras. Quizás lo más importante es que su forma de vida es ahora reconociblemente humana, con herramientas más avanzadas, caza, uso del fuego y colonizando nuevos ambientes fuera de África.
Comparado con Homo habilis, Homo erectus tuvo un tamaño cerebral mayor (promedio de alrededor de 900 cc en comparación con 650 cc a 750 cc para habilis). En lugar de tener una forma redondeada como la que tienen nuestros cráneos, el cráneo erecto era largo y bajo como una pelota de fútbol, con una frente retrocedida, y una cresta horizontal llamada toro occipital que le daba a la parte posterior del cráneo una apariencia cuadrada. Los huesos craneales son más gruesos que los de los humanos modernos, y algunos cráneos de Homo erectus tienen un ligero engrosamiento a lo largo de la sutura sagital llamada quilla sagital. Grandes crestas de cejas parecidas a un estante cuelgan sobre los ojos. Como se señaló anteriormente, el clima era cada vez más árido y el dosel del bosque en algunas partes de África estaba siendo reemplazado por un entorno de pastizales más abierto, resultando en una mayor exposición al sol para nuestros antepasados. En comparación con los australopitecinos anteriores, los miembros del género Homo también estaban desarrollando cuerpos y cerebros más grandes, comenzando a obtener carne cazando o recogiendo cadáveres, y elaborando sofisticadas herramientas de piedra. Para fines de enfriamiento, H. erectus pudo haber tenido poco vello corporal acompañado de piel oscura para proteger contra la exposición al sol. En general se acepta que el Homo erectus fue el primer homínido en migrar fuera de África y colonizar Asia y más tarde Europa (aunque los descubrimientos recientes en Asia pueden desafiar esta visión).
Fósiles de Homo erectus
Con base en datos fósiles actuales, el Homo erectus existió entre 1.9 millones (mya) a hace 25 mil años (Jurmain 2013). H. erectus, literalmente “humano erectus”, se han encontrado fósiles en Java (Indonesia), África, China, Europa e Israel. Con base en diferencias morfológicas en el cráneo, algunos científicos identifican dos especies, H. erectus en Asia y H. ergaster en África, siendo los especímenes africanos más pequeños que los asiáticos; sin embargo, utilizaremos la designación H. erectus para ambas.
En 1984, se encontró un esqueleto de H. erectus casi completo a lo largo de un río en el norte de Kenia. La datación por potasio-argón coloca a “Turkana Boy” entre 1.64 y 1.33 millones de años atrás (mya). El envejecimiento y el sexado de los restos fósiles indican que el individuo era un macho de aproximadamente ocho años de edad. Él medía alrededor de 5'3” de altura. Estudios recientes indican que Turkana Boy siguió el patrón de crecimiento de los simios, por lo que habría estado cerca de su estatura adulta al momento de su muerte (Jurmain et al. 2013).
Homo erectus tiene una larga historia en Indonesia; los fósiles de Java fueron fechados por argón que datan de aproximadamente 1.6 millones a 1.8 millones de años. Los fósiles de H. erectus del sitio de Ngandong en Java han arrojado fechas muy recientes de 43 mil años, aunque un estudio más reciente con diferentes métodos de datación concluyó que eran mucho mayores, entre 140 mil y 500 mil años de antigüedad.
El patrón de aumento del tamaño cerebral continuó con H. erectus; su cerebro es hasta 50% más grande que su predecesor, H. habilis (O'Neil c1999-2012). Este gran cerebro estaba apoyado por una dieta pesada en carne y otras proteínas. Sus características distintivas incluyen su quilla sagital (un engrosamiento del hueso que va de adelante hacia atrás en la parte superior del cráneo; probable para la unión muscular) crestas masivas de la frente (toro supraorbital) y prominencias óseas en la parte posterior del cráneo. Postcranealmente, los huesos de H.erectus son más gruesos que H. habilis, al igual que sus mandíbulas y huesos faciales, y la proporción de brazos a piernas es como la de los humanos modernos, haciendo que algunos sugieran que su andar bípedo era como el nuestro. La longitud de los huesos de sus piernas indica que H. erectus habría sido un eficiente corredor de larga distancia, permitiéndoles “atropellar animales de caza pequeños e incluso medianos en las sabanas tropicales de África Oriental” (O'Neil c1999-2012). Si es así, como se mencionó anteriormente, es probable que H. erectus tuviera mucho menos vello corporal que sus predecesores, ya que habrían necesitado poder sudar eficientemente para enfriar el cuerpo. Es posible que H. erectus tuviera poco vello corporal (NOVA 2011). Durante el Pleistoceno, el clima era cada vez más árido y el dosel del bosque en algunas partes de África estaba siendo reemplazado por un entorno de pastizales más abierto, lo que resultó en una mayor exposición al sol para nuestros antepasados.
En comparación con los australopitecinos anteriores, los miembros del género Homo también estaban desarrollando cuerpos y cerebros más grandes, comenzando a obtener carne cazando o recogiendo cadáveres, y elaborando sofisticadas herramientas de piedra. Según Nina Jablonski, experta en la evolución de la piel humana, la pérdida de vello corporal y el aumento de la capacidad de sudoración forman parte del paquete de rasgos que caracterizan al género Homo. Si bien cerebros más grandes y cuerpos de patas largas hicieron posible que los humanos cubrieran largas distancias mientras buscaban comida, esta nueva forma corporal tuvo que enfriarse efectivamente para manejar un estilo de vida más activo. Prevenir que el cerebro se sobrecaliente fue especialmente crítico. La capacidad de mantenerse frescos también pudo haber permitido que los homínidos se alimentaran durante la parte más calurosa del día, dándoles una ventaja sobre los depredadores de sabanas, como los leones, que normalmente descansan durante el calor del día.
Como se señaló anteriormente, los científicos generalmente coinciden en que H. erectus fue el primer homínido en salir de África. Como se mencionó anteriormente, se han encontrado fósiles en África, el norte de China, Indonesia, Europa e Israel. En la República de Georgia, se encontraron fósiles en estratos fechados en 1.7mya, lo que sugiere que H. erectus abandonó África poco después de su evolución. Un reporte reciente (Dembo et al. 2015) postula que H. habilis fue el primer homínido en salir de África, no H. erectus. Si esta contención se apoyara con más datos, todavía se puede argumentar que H. erectus tuvo bastante éxito en colonizar el Viejo Mundo (África, Europa y Asia), ayudado, sin duda, por sus comportamientos culturales avanzados.
Ahora podemos abordar la pregunta de por qué Homo erectus viajó distancias tan vastas a estas regiones remotas. Para ello, tenemos que considerar lo que hemos aprendido sobre la biología, la cultura y las circunstancias ambientales del Homo erectus. El mayor tamaño cerebral y corporal del Homo erectus fueron alimentados por una dieta que consistía en más carne, y las patas más largas y poderosas permitieron caminar y correr distancias más largas para adquirir alimentos. Dado que estaban comiendo más arriba en la cadena alimentaria, era necesario que ampliaran su rango hogareño para encontrar suficiente caza. Los desarrollos culturales, incluyendo mejores herramientas de piedra y nuevas tecnologías como el fuego, les dieron mayor flexibilidad para adaptarse a diferentes ambientes. Por último, el gran cambio climático del Pleistoceno discutido anteriormente en el capítulo ciertamente jugó un papel. Los cambios en la temperatura del aire, la precipitación, el acceso a fuentes de agua y otras alteraciones del hábitat tuvieron efectos de gran alcance en las comunidades de animales y plantas; esto incluyó Homo erectus. Si los homínidos confiaran más en la caza, los patrones de migración de sus presas podrían haberlos llevado a distancias cada vez más largas.
Hay evidencia de Homo erectus en China de varias regiones y periodos de tiempo. Los fósiles de Homo erectus del norte de China, conocidos colectivamente como “Hombre de Pekín”, son algunos de los fósiles humanos más famosos del mundo, fechados alrededor de 400,000 a 700,000 años atrás. El descubrimiento de herramientas de piedra de China que datan de 2.1 millones de años, más antiguas que cualquier fósil de Homo erectus en cualquier lugar, abre la intrigante posibilidad de que los homínidos antes que el Homo erectus pudieran haber migrado fuera de África.
En este momento, los investigadores no están de acuerdo en si los primeros europeos pertenecían al Homo erectus propiamente dicho o a una especie descendiente posterior.
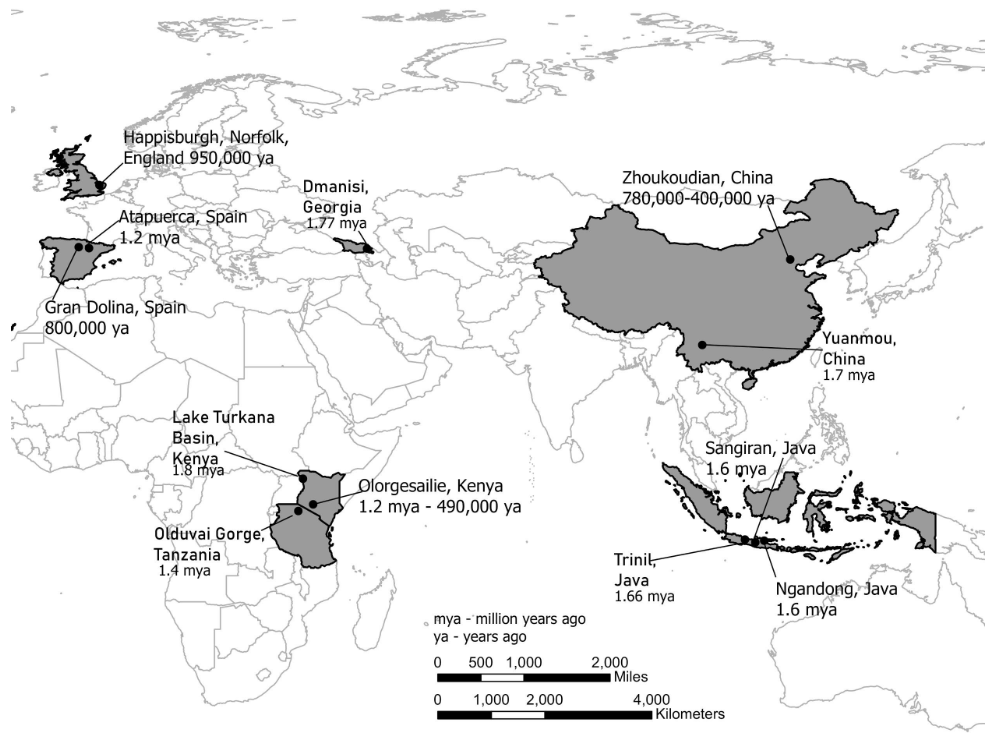
Figura\(\PageIndex{21}\): Mapa que muestra las ubicaciones de los fósiles de Homo erectus alrededor de África y Eurasia (adaptado por Kenneth A. Koenigshofer, PhD, Chaffey College, de Exploraciones: Una invitación abierta a la antropología biológica;https://explorations.americananthro.....php/chapters/; Capítulo 10, “Primeros miembros del género Homo” de Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, y Lara Braff, Eds.; licenciado bajo una Licencia Creative Commons Reconocimiento-NoComercial 4.0 Internacional, excepto donde se indique otra cosa).
Homo erectus Cultura
Homo erectus muestra importantes innovaciones culturales en dieta, tecnología, historia de vida, ambientes ocupados, y quizás incluso organización social, algunos que probablemente reconocerás como más “parecidos a los humanos” que cualquiera de los homínidos previamente cubiertos. Hace aproximadamente 1.5 millones de años, algunas poblaciones de Homo erectus comenzaron a hacer diferentes formas de herramientas (conocidas como “cultura material”; ver Sección 3.14). Estas herramientas, clasificadas juntas como constitutivas de la industria de herramientas achelana, son más complejas en su forma y más consistentes en su fabricación. A diferencia de las herramientas Oldowan, que eran adoquines modificados al golpear algunas escamas, los fabricantes de herramientas achelenses moldearon cuidadosamente ambos lados de la herramienta. Este tipo de técnica, conocida como descamación bifacial, requiere más planeación y habilidad por parte del fabricante de herramientas; él o ella necesitaría ser consciente de los principios de simetría a la hora de elaborar la herramienta. Una de las formas de herramienta más comunes de este periodo fue el handaxe. Además de las mandaxes, en los sitios del Homo erectus están presentes formas como raspadores, raspadoras y herramientas en escamas. Un aspecto llamativo de las herramientas achelanas es su uniformidad. Están más estandarizados en forma y modo de fabricación que las herramientas Oldowan anteriores. Por ejemplo, los mandaxes mencionados varían en tamaño, pero son notablemente consistentes en cuanto a su forma y proporciones. También fueron una forma de herramienta increíblemente estable a lo largo del tiempo, durando más de un millón de años con pocos cambios.
Recientemente, los métodos más nuevos, incluido el análisis microscópico de rocas y huesos quemados, han revelado evidencia clara del uso del fuego en Koobi Fora, Kenia, que data de hace 1.5 millones de años (Hlubik et al. 2017).
Existe un consenso general de que H. erectus evolucionó a partir de H. habilis y Homo heidelbergensis evolucionó de H. erectus en África, suplantando eventualmente poblaciones de H. erectus en el Viejo Mundo: África, Asia y Europa (Figura\(\PageIndex{18}\)).
Uso de herramientas y habilidades cognitivas del Homo erectus
¿Qué (si acaso) nos dicen las herramientas achelanas sobre la mente del Homo erectus? Claramente, tomaron una buena cantidad de habilidad para fabricar. Aparte de la configuración real de la herramienta, otras decisiones tomadas por los fabricantes de herramientas pueden revelar su uso de la previsión y la planificación. ¿Simplemente escogieron las rocas más convenientes para hacer sus herramientas, o buscaron una materia prima en particular que sería ideal para una herramienta en particular? El análisis de las herramientas de piedra achelana sugiere que en algunos sitios, los fabricantes de herramientas seleccionaron cuidadosamente sus materias primas, viajando a afloramientos rocosos particulares a piedras de cantera y tal vez incluso quitando grandes losas de roca en las canteras para obtener el material más deseable. Tales actividades complejas requerirían una planeación avanzada. También probablemente requirieron cooperación y comunicación con otros individuos, ya que tales acciones serían difíciles de llevar a cabo en solitario. Sin embargo, otros sitios de Homo erectus carecen de evidencia de tal selectividad; en lugar de viajar incluso una distancia corta para obtener una mejor materia prima, los homínidos tendían a usar lo que estaba disponible en su área inmediata (Shipton et al. 2018). En contraste con las herramientas de Homo erectus, las herramientas del Homo sapiens moderno temprano durante el Paleolítico Superior muestran una enorme diversidad entre regiones y períodos de tiempo. Adicionalmente, las herramientas y artefactos del Paleolítico Superior comunican información como el estado y la pertenencia al grupo. Tal innovación y señalización social parecen haber estado ausentes en el Homo erectus, lo que sugiere que tenían una relación diferente con sus herramientas que la del Homo sapiens (Coolidge y Wynn 2017). Algunos científicos afirman que estos contrastes en la forma y fabricación de herramientas pueden significar diferencias cognitivas clave entre las especies, como la capacidad de usar un lenguaje complejo.
Cuadro 3.3.3. Características deHomo erectus. (adaptado por Kenneth A. Koenigshofer, PhD, Chaffey College, de Exploraciones: una invitación abierta a la antropología biológica;https://explorations.americananthro.....php/chapters/; Capítulo 10, “Primeros miembros del género Homo” por Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, y Lara Braff, Eds.; licenciado bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional, excepto cuando se indique otra cosa).
Híminin
Homo erectus
Fechas
Hace 1.8 millones de años hasta hace unos 200,000 años
Región (es)
África Oriental y Sudafricana; Eurasia Occidental; China y Sudeste Asiático
Descubrimientos famosos
Lago Turkana, Olorgesailie, Kenia; Zhoukoudian, China; Dmanisi, República de Georgia
Tamaño del Cerebro
Promedio 900 cc; rango entre 650 cc y 1,100 cc
Dentición
Dientes más pequeños que Homo habilis
Características craneales
Cráneo largo y bajo con características robustas que incluyen huesos gruesos de la bóveda craneal y cresta grande de la ceja, quilla sagital y toro occipital
Características postcraneales
Tamaño corporal más grande comparado con Homo habilis; proporciones corporales (piernas más largas y brazos más cortos) similares a Homo sapiens
Cultura
Herramientas achelanas (en África) (ver Módulo 3.8); evidencia de aumento de la caza y el consumo de carne; uso del fuego; migración fuera de África
EL GRAN FOTO DEL HOMO TEMPRANO
Estamos descubriendo que la evolución del género Homo es más compleja de lo que se pensaba anteriormente. La visión predominante anterior de una simple progresión de Australopithecus a Homo habilis a Homo erectus como etapas claramente delineadas en la evolución humana simplemente no se sostiene más.
Variabilidad en el Registro Fósil del Homo Temprano
Como se desprende de la información aquí presentada, existe una tremenda variabilidad durante este tiempo. Si bien los fósiles clasificados como Homo habilis muestran muchas de las características del género Homo, como la expansión cerebral y el menor tamaño de los dientes, el tamaño corporal pequeño y los brazos largos son más parecidos a los australopitecinos. También hay una tremenda variabilidad dentro de los fósiles asignados al Homo habilis, por lo que no hay consenso sobre si se trata de una especie única o múltiple de Homo, un miembro del género Australopithecus, o incluso un nuevo género aún por definir.
¿Qué significa esta diversidad para cómo deberíamos ver el Homo temprano? En primer lugar, no hay una ruptura abrupta entre Australopithecus y Homo habilis o incluso entre Homo habilis y Homo erectus. Las características que definimos como Homo no aparecen como un paquete unificado; aparecen en el registro fósil en diferentes momentos. Esto se conoce como evolución del mosaico.
Podemos considerar varias explicaciones para la diversidad que vemos dentro del Homo temprano desde hace unos 2.5 millones a 1.5 millones de años atrás. Una posibilidad es la existencia de múltiples especies contemporáneas de Homo temprano durante este período. A la luz del patrón de inestabilidad ambiental discutido anteriormente, no debería ser sorprendente ver fósiles de diferentes partes de África y Eurasia mostrar una tremenda variabilidad. Múltiples formas de homínidos también podrían evolucionar en una misma región, ya que se diversificaron para ocupar diferentes nichos ecológicos. Sin embargo, incluso la presencia de múltiples especies de homínidos no impide que interactúen y se crucen entre sí.
La diversidad de tamaños cerebrales y corporales también podría reflejar la plasticidad del desarrollo: adaptaciones a corto plazo dentro de la vida (Anton, Potts y Aiello, 2014). Estos tienen la ventaja de ser más flexibles que la selección genética natural, que sólo podría ocurrir a lo largo de muchas generaciones. Por ejemplo, entre las poblaciones humanas de hoy en día, se piensa que diferentes tamaños corporales son adaptaciones a diferentes climas o ambientes nutricionales.
Tendencias en el Comportamiento del Homo Temprano
Nuevos descubrimientos también cuestionan viejas suposiciones sobre el comportamiento de Homo habilis y Homo erectus. Así como la evidencia fósil no separa perfectamente Australopithecus y Homo, la evidencia de las formas de vida del Homo temprano muestra una diversidad similar. Por ejemplo, se pensó que una de las líneas divisorias tradicionales entre Homo y Australopithecus eran herramientas de piedra: Homo las hizo; Australopithecus no lo hizo. sin embargo, el reciente descubrimiento de herramientas de piedra de Kenia que data de hace 3.3 millones de años desafía este punto de vista. De igual manera, la creencia de que el Homo erectus fue la primera especie en asentarse fuera de África puede ahora cuestionarse con el reporte de herramientas de piedra de 2.1 millones de años de antigüedad provenientes de China. Si este hallazgo está respaldado por evidencia adicional, puede provocar una reevaluación del Homo erectus siendo el primero en salir. En cambio, podría haber habido múltiples migraciones anteriores de homínidos como Homo habilis o incluso especies de Australopithecus.
En lugar de demarcaciones obvias entre las especies y sus correspondientes avances de comportamiento, ahora parece que muchos comportamientos fueron compartidos entre las especies. A pesar de la nebulosidad que domina la narrativa del Homo temprano, podemos identificar algunas tendencias generales para el período de un millón de años asociado al Homo temprano. Estas tendencias incluyen expansión cerebral, reducción del prognatismo facial, menor tamaño de mandíbula y diente, mayor tamaño corporal y evidencia de bipedalismo terrestre completo. Estos rasgos están asociados con un cambio conductual clave que enfatiza la cultura como una estrategia flexible para adaptarse a circunstancias ambientales impredecibles. En este repertorio se incluyen la creación y el uso de herramientas de piedra para procesar la carne obtenida mediante la recolección y posterior caza, la utilización del fuego y la cocción, y las raíces del patrón de la historia de vida humana de la infancia prolongada, la cooperación en la crianza de los hijos y la práctica de técnicas de forrajeo experto. De hecho, es evidente que las innovaciones culturales están impulsando los cambios biológicos, y viceversa, alimentando un ciclo de retroalimentación que continuará durante las últimas etapas de la evolución humana.
Homo heidelbergensis: el ancestro común del Homo neanderthalensis en Europa y del Homo sapiens en África.
Algunas publicaciones, por ejemplo Larsen 2014, se refieren a H. heidelbergensis como el arcaico Homo sapiens, pero para nuestros fines, usaremos la designación H.heidelbergensis. Otto Schoetensack encontró los primeros fósiles de Homo heidelbergensis en 1907 en Mauer, Alemania. Desde entonces se han encontrado fósiles de H. heidelbergensis en África, Europa y Asia. El rango de fechas para la especie es de 800 kya (hace mil años) a 350 kya. Los rasgos primitivos (rasgos como los ancestros) incluyen su gran toro supraorbital (cresta de la frente), quilla sagital y hueso frontal bajo. Los rasgos derivados (rasgos diferentes al antepasado) incluyen toros supraorbitales separados sobre cada órbita ocular, una bóveda craneal posterior más vertical, huesos parietales anchos en relación con la base craneal y una capacidad craneal mayor que H. erectus (Becoming Human c2008). Adicionalmente, exhiben dimorfismo sexual similar al de los humanos modernos.
Figura\(\PageIndex{22}\): Cráneo de Homo heidelbergensis sobre un soporte de acero espejado que muestra reflejo del paladar y dentición en mandíbula superior.
Existe variación regional en la morfología de H. heidelbergensis. Especímenes europeos encontrados en Atapuerca, (España), Petralona (Grecia), Seinheim (Alemania) y Swancombe (Inglaterra) muestran que tenían cuerpos compactos, lo que podría haber sido una respuesta a vivir en los climas fríos del norte ya que ayudaría a conservar el calor. Adicionalmente, el cráneo es un mosaico de rasgos de H. erectus y rasgos derivados. En Asia, los datos de sitios como Zhoukoudian, Jinniushan y Dalí (China) muestran una mezcla de rasgos de H. erectus y H. sapiens; este último incluye gran capacidad craneal y paredes delgadas del cerebro. Los especímenes africanos de Kabwe (Zambia), Florisbad (Sudáfrica), Laetoli (Tanzania) y Bodo (Etiopía) también muestran una combinación de rasgos de H. erectus y H. sapiens. Comparte el toro supraorbital masivo y el toro occipital prominente con H. erectus y bóveda craneal delgada, occipital menos angulada y base craneal con H. sapiens.
Significativamente, H. heidelbergensis es el ancestro común del Homo neanderthalensis en Europa y del Homo sapiens en África.
Homo neanderthalensis
Numerosos fósiles de neandertales han sido recuperados desde su descubrimiento en 1856 en el Valle de Neander, Alemania. Los neandertales han sido la especulación de los científicos y del público en general desde entonces. Algunos antropólogos clasifican a los neandertales como una subespecie de Homo sapiens, Homo sapiens neanderthalensis, mientras que otros interpretan las diferencias morfológicas como lo suficientemente significativas como para justificar su clasificación como una especie diferente, Homo neanderthalensis. Aquí, utilizaremos esta última designación.
Figura\(\PageIndex{23}\): Homo neanderthalensis.
Los verdaderos neandertales aparecen por primera vez en el registro fósil hace unos 200 mil años (200 kya), con fósiles que exhiben características similares al neandertal apareciendo ya en 400 kya. Investigaciones recientes indican que los neandertales se extinguieron hace apenas unos 40 mil años, entre 41 kya y 39 kya (Higham et al., 2014). La investigación molecular denota que parte del ADN neandertal vive en los humanos modernos, aproximadamente el 2% del ADN de “personas que descienden de europeos, asiáticos y otros no africanos es neandertal” (Callaway, 2014). Los genes neandertales están involucrados en combatir infecciones, lidiar con la radiación ultravioleta (Callaway, 2014), y vivir a gran altura (Callaway, 2015). El ADN neandertal se ha relacionado recientemente con depresión, obesidad y ciertos trastornos de la piel, por ejemplo, lesiones causadas por la exposición al sol (Callaway, 2015).
Los neandertales son el único homínido que originalmente evolucionó en un ambiente glacial, dando lugar a algunas características adaptadas al clima frío. Muchos científicos sostienen que el prognatismo de la mitad de la cara (media cara sobresaliente) permitió agrandamiento de los senos paranasales que funcionaban para calentar y agregar humedad al aire frío y seco antes de ingresar a los pulmones. Pequeños agujeros debajo de las órbitas oculares, llamados foramina infraorbitaria, son más grandes en los neandertales europeos que los humanos modernos, lo que sugiere que los vasos sanguíneos eran más grandes, lo que permitió un mayor flujo sanguíneo a la cara. Esto habría ayudado a mantener la cara más caliente. Son relativamente cortos y robustos, los machos promedian 5' 5” y las hembras 5'1”, con apéndices más cortos, los cuales ayudarían a conservar el calor al proporcionar menos superficie desde la que irradiar calor. Sus huesos de las piernas son gruesos y densos, lo que sugiere que frecuentemente caminaban y corrían, muy probablemente en actividades de adquisición de alimentos. Algunos postulan que algunos neandertales tenían piel pálida que habría ayudado a aumentar la síntesis de vitamina D al aumentar la cantidad de radiación UV para ser absorbida por el cuerpo (O'Neil, c1999-2014). El ancho de sus troncos corporales y las tibias cortas se ajustan a las predicciones para la adaptación al clima frío como propone Christopher Ruff (Larsen, 2013). El tamaño cerebral de los neandertales también puede estar relacionado con el clima frío. Con un promedio de una capacidad craneal entre 1300 y 1400 cm3, tienen los cerebros más grandes de todos los homínidos, incluyendo nuestra propia especie, el Homo sapiens. El tamaño puede estar asociado con una mayor eficiencia metabólica en clima frío, lo cual es similar a los pueblos inuit modernos de hoy que tienen un tamaño cerebral mayor que otras poblaciones humanas (Jurmain et al., 2013).
La presencia de un moño occipital (un bulto prominente en la parte baja de la espalda del cráneo) es una de las características utilizadas para identificar especímenes de H. neanderthalensis, aunque cabe señalar que estas características persisten en un pequeño porcentaje de las poblaciones humanas modernas. El moño occipital puede haber evolucionado para contrarrestar la cara pesada de Neanderthal al correr; evita que la cabeza haga enormes aceleraciones horizontales (NOVA 2002). El moño occipital también alarga el cráneo en comparación con la mayoría de Homo sapiens. Tenían fuertes crestas de cejas como las de H. heidelbergensis. Los cráneos neandertales recuperados de Amud y Tabún en Israel presentan más características craneales similares a H. sapiens, incluyendo falta de moño occipital, órbitas oculares más pequeñas, aberturas nasales altas y anchas y dientes más pequeños (Larsen 2014).
Denisovanos
Los denisovanos eran humanos arcaicos estrechamente relacionados con los neandertales, cuyas poblaciones se superponían geográficamente con los antepasados de los humanos modernos que existían al mismo tiempo. En 2010, los científicos anunciaron el descubrimiento y análisis de ADN de un hueso de dedo y dos dientes encontrados en la cueva Denisova, Siberia (Reich et al., 2010). Los artefactos fueron recuperados de un depósito fechado de 50 kya a 30 kya. Los datos sugieren que los restos eran de un individuo que compartía un origen común con los neandertales, pero no era un neandertal, ni era un humano moderno. Los individuos denisovanos comparten 4-6% de su material genético con pueblos modernos que viven en Nueva Guinea, Islas Bougainville y China. Otros estudios (Cooper y Stringer, 2013) indican que los denisovanos cruzaron Eurasia y se cruzaron con humanos modernos, pero genéticamente los denisovanos estaban más estrechamente relacionados con los neandertales que con los humanos modernos (Meyer et al., 2012). Si bien otros hallazgos arrojarán más luz sobre los denisovanos, es claro que hubo más variabilidad genética durante el Pleistoceno de lo que se pensaba anteriormente (Larsen, 2014), pero que el flujo de genes entre las diversas poblaciones tuvo implicaciones para la emergencia de los humanos modernos (Pääbo, 2015).
Figura\(\PageIndex{24}\): Diseminación y evolución de los denisovanos.
Homo floresiensis
En 2003, un equipo de científicos descubrió fósiles inusuales en la cueva de Liang Bua, Isla Flores, Indonesia. La pequeña estatura del individuo condujo al nombramiento de una nueva especie, Homo floresiensis. Desde entonces se han encontrado doce individuos que van en el tiempo de 74 kya a 17 kya. H. floresiensis tenía un cerebro pequeño y solo tenía cerca de 3.5' de altura cuando era adulto. Tenían frentes retrocedidas, sin barbillas, hombros encogidos hacia adelante y pies grandes en relación con sus piernas cortas. Comparten algunas características con H. sapiens, incluyendo dentición más pequeña, crestas de cejas separadas y una cara no prognática (cara aplanada).
Figura\(\PageIndex{25}\): Homo floresiensis.
Se han propuesto varias hipótesis para explicar la aparición de H. floresiensis. Uno sugiere que estamos viendo enanismo insular, que es un proceso evolutivo resultante del aislamiento a largo plazo en una isla pequeña. Recursos limitados y falta de depredadores seleccionados para individuos de cuerpo más pequeño que necesitan menos recursos que los individuos de cuerpo grande. Otra hipótesis afirma que H.floresiensis no es una especie separada, sino que H. sapiens exhibe microcefalia o alguna otra deficiencia del desarrollo como el cretinismo hipertiroideo (Oxnard et al. 2010) porque las “... características craneales... están dentro del rango moderno de variación observada en poblaciones vivas de la región más grande [de Indonesia]” (Larsen 2014:409), dando apoyo a la segunda hipótesis. La forma en que H. floresiensis encaja en el panorama evolutivo no está claro con base en los datos actuales.
Homo sapiens
En la actualidad, solo una especie de homínidos habita en el planeta Tierra, el Homo sapiens. Todas las demás especies de Homo se extinguieron hace aproximadamente 30,000 a 40,000 años. Muchos científicos creen que estas extinciones fueron impulsadas por la competencia del Homo sapiens. Morfológicamente, las características de H. sapiens incluyen la presencia de un mentón, un cerebro grande, cara plana, cráneo redondeado o globular, y una cresta ceja continua y reducida. Sus huesos son graciles (delgados) en comparación con los homínidos anteriores, aunque los primeros H. sapiens son más robustos que las poblaciones modernas y no muestran ninguna de las adaptaciones de clima frío de los neandertales. La naturaleza más gracile da crédito a la hipótesis de que los humanos modernos evolucionaron primero en África. Las proporciones corporales más delgadas son más adaptativas a los ambientes tropicales africanos ya que habría más superficie corporal para irradiar calor. La evidencia de ADN también respalda un origen africano. Las poblaciones africanas modernas tienen más diversidad genética que cualquier otra población humana moderna, inferyendo que han estado evolucionando más tiempo (Becoming Human c2008).
Figura\(\PageIndex{26}\): Homo sapiens.
Los fósiles de Homo sapiens más antiguos se encontraron en África en Omo Kibish, Etiopía, fechados hace casi 200 mil años (195 kya; kya = hace mil años). Los fósiles de Homo sapiens encontrados en Herto, Etiopía (160 kya—154 kya), la boca del río Klasies y la cueva fronteriza, Sudáfrica (120 kya — 89 kya), Skhūl, Israel (130 kya—100 kya) y Qafzeh, Israel (120 kya—92 kya), agregan más apoyo a la era del Homo sapiens. Los sitios en Asia, por ejemplo, en China y Borneo, Indonesia datan de 40 kya o menos y en Europa no datan antes de 31 kya (Mladeč, República Checa). Los humanos modernos de alguna manera llegaron a Australia sobre mar abierto por 55 kya; sin embargo, no se han encontrado restos humanos en el continente australiano que data antes de 30 kya en el lago Mungo. Estos datos dan poco apoyo a la Hipótesis Multirregional, un modelo que propuso que H. sapiens moderno evolucionó en lugares separados del Viejo Mundo a partir de poblaciones locales arcaicas de H. sapiens. Este modelo sugirió que el flujo de genes entre las poblaciones condujo a la similitud genética de los humanos modernos. La mayoría de los antropólogos apoyan el modelo de origen africano. Hay dos variantes de este modelo. El modelo Out of Africa afirma que los humanos modernos surgieron en África y se extendieron a Europa después de 50 kya, reemplazando a la población no H. sapiens sin flujo génico. Una alternativa es el modelo de Asimilación que también afirmó que el humano moderno surgió por primera vez en África y se extendió a Europa y Asia. La principal diferencia entre los modelos Out of Africa y Asimilation es que este último afirma flujo génico entre H. sapiens y H. neanderthalensis. Teniendo en cuenta los recientes análisis de ADN de neandertales discutidos anteriormente, múltiples líneas de evidencia apoyan el modelo de Asimilación.
El caso de los neandertales en desaparición
Sabemos que algunos genes neandertales están en el genoma de los humanos modernos, por lo que en esencia, alguna parte de los neandertales sobrevive hoy. Sin embargo, no hay neandertales caminando en el mundo moderno. Entonces, ¿qué pasó con los neandertales? Los científicos sugieren que H. sapiens superó a los neandertales con su dieta más diversa y sus “habilidades sofisticadas y cognitivas” (Becoming Human c2008), lo que puede haber incluido una capacidad superior para formar imágenes visuales en la imaginación para pronosticar los resultados de acciones futuras, mejorando así planeación e inventiva (Koenigshofer, 2017). Estos rasgos permitieron a H. sapiens adaptarse fácilmente a las condiciones climáticas rápidamente cambiantes durante el Paleolítico Superior.
Una hipótesis reciente sugiere que los neandertales se extinguieron durante un período de poderosa actividad volcánica en Eurasia occidental. Las excavaciones en la cueva Mezmaiskaya en las montañas del Cáucaso en el sur de Rusia han recuperado una gran cantidad de huesos neandertales, herramientas de piedra y restos de animales presa. Los restos y artefactos neandertales aparecen en estratos por encima de una capa de ceniza volcánica y debajo de una segunda capa de ceniza volcánica. No se han encontrado huesos ni artefactos neandertales por encima del segundo nivel, lo que sugiere que los neandertales ya no ocuparon la zona. Ambas capas de ceniza contienen niveles de polen asociados con climas más fríos y secos. “Las capas de ceniza corresponden cronológicamente a lo que se conoce como la súper erupción campaniana de Ignimbrita, que ocurrió hace alrededor de 40 mil años en la Italia moderna, y una erupción más pequeña que se cree que ocurrió alrededor de la misma época en las montañas del Cáucaso” (University of Chicago Press Journals 2010). El invierno volcánico subsiguiente provocó un cambio climático dramático que llevó a la desaparición de los neandertales. Estos datos se correlacionan con la investigación de Higham et al. (2014) que afirma que los neandertales se extinguieron entre 41 mil y 39 mil años atrás.
Evolución del Cerebro
Muchos científicos (Cajal, 1937; Crick & Koch, 1990; Edelman, 2004) creen que el cerebro humano es la máquina más compleja del universo conocido. Su complejidad apunta a un hecho innegable: que ha evolucionado lentamente con el tiempo a partir de formas más simples. La evolución del cerebro es intrigante no sólo porque podemos maravillarnos con esta complicada estructura biológica, sino también porque revela una larga historia de muchos sistemas nerviosos menos complejos (Figura 3.3.25), lo que sugiere un registro de comportamientos adaptativos en formas de vida distintas a las humanas. Por lo tanto, el estudio evolutivo del sistema nervioso es importante, y es el primer paso para comprender su diseño, su funcionamiento y su interfaz funcional con el entorno.
Figura\(\PageIndex{27}\): Los cerebros de diversos mamíferos (no a escala). Los cerebros mostrados se representan artificialmente como aproximadamente iguales en tamaño. Aunque no se muestran las grandes diferencias en el tamaño real de estos cerebros, todavía es posible ver el grado creciente de plegamiento de la corteza cerebral como lo indica el mayor número de fisuras y giros (valles y colinas en la superficie de la corteza, indicados por el número de líneas) en la superficie del chimpancé y cerebros humanos comparados con los demás. Tenga en cuenta que la corteza del ratón es lisa, no muestra ningún plegamiento, lo que indica menos corticalización: desarrollo cortical. El mayor plegamiento de la corteza cerebral ocurre en humanos revelando una tendencia en la evolución de mamíferos y primates hacia el aumento de la corticalización junto con el aumento del tamaño del cerebro. Para más información sobre las tendencias de la evolución cerebral en mamíferos y primates, ver sección 18.13.
Los cerebros de algunos animales, otros mamíferos, como simios, monos y roedores, son estructuralmente similares a los humanos (Figura 3.3.27), mientras que otros no lo son (por ejemplo, invertebrados como el pulpo). ¿La similitud anatómica de estos cerebros sugiere que los comportamientos que emergen en estas especies también son similares? De hecho, muchos animales muestran comportamientos similares a los humanos, por ejemplo, los simios usan señales de comunicación no verbal con sus manos y brazos que se asemejan a formas de comunicación no verbales en humanos (Gardner & Gardner, 1969; Goodall, 1986; Knapp & Hall, 2009). Si estudiamos respuestas fisiológicas muy simples hechas por neuronas individuales (células nerviosas), las respuestas neuronales de invertebrados como la babosa marina (Kandel & Schwartz, 1982) se ven muy similares a las respuestas neuronales de neuronas individuales en mamíferos, incluyendo humanos, sugiriendo que a partir del tiempo inmemoriales tales funciones básicas de las células nerviosas individuales en el cerebro de muchas formas animales simples se han conservado a lo largo de la evolución de animales más complejos y que de hecho estas características fundamentales de las células nerviosas son la base de comportamientos más complejos en animales que evolucionaron más tarde ( Bullock, 1984).
Aunque los procesos fisiológicos básicos en las células nerviosas son muy similares entre especies, sin embargo incluso a nivel microanatómico, observamos que las neuronas individuales difieren en complejidad en algunos aspectos entre las especies animales. Las neuronas humanas presentan una complejidad más intrincada que las neuronas en otros animales; por ejemplo, los procesos neuronales (dendritas, que reciben entrada de otras neuronas) en humanos tienen muchos más puntos de ramificación, ramas y espinas dendríticas asociadas con conexiones entre neuronas (ver Capítulo 10 sobre Aprendizaje y Memoria).
La complejidad en la estructura del sistema nervioso, tanto a nivel macro como micro, da lugar a comportamientos complejos. Podemos observar movimientos similares de las extremidades, como en la comunicación no verbal, en simios y humanos, pero la variedad y complejidad de los comportamientos no verbales usando las manos en humanos supera a los simios. Esto se relaciona con el hecho de que los humanos tienen más neuronas para el control del movimiento de las manos que los simios. Las personas sordas que usan el lenguaje de señas americano (ASL) se expresan en inglés de manera no verbal; usan este idioma con una gradación tan fina que existen muchos acentos de ASL (Walker, 1987). Complejidad de comportamiento con creciente complejidad del sistema nervioso, especialmente la corteza cerebral, se puede observar en el género Homo (Figuras 3.3.27 y 3.3.28). Si comparamos la sofisticación de la cultura material en Homo habilis (hace 2 millones de años; volumen cerebral ~650 cm3) y Homo sapiens (300,000 años hasta ahora; volumen cerebral ~1400 cm3), la evidencia muestra que Homo habilis utilizó piedra cruda herramientas en marcado contraste con las herramientas modernas utilizadas por Homo sapiens para erigir ciudades, desarrollar lenguajes escritos, crear industrias complejas y embarcarse en viajes espaciales. Todo esto se debe a la creciente complejidad del cerebro.
Figura\(\PageIndex{28}\): Aumento del volumen cerebral y otras características anatómicas y conductuales, incluido el desarrollo de cultivos materiales (incluyendo herramientas de piedra) sobre la evolución de los homínidos. Tenga en cuenta que tanto el volumen cerebral como los avances culturales se aceleran tras el primer uso del fuego. Falta en este diagrama el cerebro de 1.400 centímetros cúbicos de los neandertales, igual o ligeramente más grande que el cerebro de los humanos modernos.
¿Qué llevó a la complejidad del cerebro y del sistema nervioso durante la evolución, y al refinamiento acompañante de la conducta y la cognición? Darwin (1859, 1871) propuso dos fuerzas: la selección natural y la selección sexual como motores de trabajo que impulsan estos cambios. Como se señaló anteriormente, Darwin profetizó, “la psicología se basará en un nuevo fundamento, el de la necesaria adquisición de cada poder y capacidad mental por gradación”. En otras palabras, Darwin predijo que la psicología se basará en la evolución (Rosenzweig, Breedlove, & Leiman, 2002).
Figura\(\PageIndex{29}\): Cronología de los homínidos que destaca las divisiones evolutivas (divergencias) de las líneas genéticas de gorila y chimpancé, las principales innovaciones conductuales y culturales y la migración fuera de África. No se muestra la migración del Homo sapiens a Europa hace unos 45,000 años donde convivieron con los neandertales durante miles de años (Imagen de Wikimedia Commons; Archivo:humantimeline-templateimage-20181222.png; https://commons.wikimedia.org/wiki/F...e-20181222.png; por Drbogdan; licenciado bajo la licencia Creative CommonsReconocimiento-Compartir Igual 4.0 Internacional).
Uso de herramientas, evolución de la inteligencia y cerebro
Como lo atestigua la discusión anterior, la construcción y el uso de herramientas es antiguo en el género Homo que data de millones de años al menos a Australopithecus. Cachel (2012, p. 222) afirma: “El comportamiento de las herramientas humanas es específico de la especie. Sigue siendo una característica diagnóstica de los humanos”. Según Osiurak y Massen (2014, p.1107), “el uso de herramientas también ha sido visto a menudo como un paso importante durante la evolución (van Schaik et al., 1999) o incluso como un marcador de la evolución de la inteligencia humana”. Vaesen (2012, p. 203) sostiene que la creación de herramientas humanas y el uso de herramientas proporcionan evidencia de “capacidad cognitiva superior única” en humanos y marcan “una discontinuidad cognitiva importante” entre humanos y simios. Según Vaesen (p. 203), existen “nueve capacidades cognitivas consideradas cruciales para el uso de herramientas: coordinación ojo-mano mejorada, plasticidad del esquema corporal, razonamiento causal, representación de funciones, control ejecutivo, aprendizaje social, enseñanza, inteligencia social y lenguaje. Dado que las diferencias notables entre humanos y grandes simios se mantienen firmes en ocho de nueve de estos dominios, [él] concluye [s] que el uso de herramientas humanas todavía marca una importante discontinuidad cognitiva entre nosotros y nuestros parientes más cercanos”. También “aborda [es] la evolución de las tecnologías humanas. En particular, [él] muestra [s] cómo los rasgos cognitivos revisados ayudan a explicar por qué la acumulación tecnológica evolucionó tan marcadamente en los humanos, y tan modestamente en los simios”. La acumulación tecnológica se refiere a la acumulación de innovaciones tecnológicas a lo largo de generaciones que conducen a una complejidad tecnológica cada vez mayor. Esto está estrechamente ligado a la gran capacidad de los humanos para la transmisión cultural eficiente de comportamientos aprendidos a lo largo de generaciones sucesivas, una habilidad que ocurre en los simios solo en un grado relativamente limitado. Vaesen (p. 213) explica: “La complejidad de las tecnologías humanas está estrechamente ligada a nuestra notable capacidad de cultivo acumulativo [audaz añadido]: Los humanos han sido capaces de construir sistemas complejos acumulando modificaciones a lo largo de generaciones sucesivas, mejorando gradualmente los logros anteriores”. Esto requiere “mecanismos sofisticados para el aprendizaje social y la enseñanza activa”, en los que el lenguaje ciertamente jugó un papel, pero es “probable que se necesiten agregar otras capacidades cognitivas; por ejemplo,.. el pensamiento causal es parte integral del aprendizaje social humano, y por lo tanto”, también se necesita para “alto- fidelidad transmisión cultural” (p. 214). Vaesen enfatiza aún más este punto: “el pensamiento causal (cada vez que surgió) es una explicación plausible de por qué la cultura acumulativa evolucionó tan marcadamente en los humanos y tan modestamente en los simios” (p.214). Cachel (2012, p. 222) va más allá y afirma que los experimentos demuestran el “gran abismo que existe entre el razonamiento causal humano y las habilidades de razonamiento de los chimpancés. De hecho, los chimpancés no parecen tener una “teoría de cómo funciona el mundo”. Si esto es así, entonces personifica el carácter único del comportamiento de la herramienta humana —que se basa en el conocimiento inconsciente y sofisticado de la energía, el movimiento, los objetos y la interacción de objetos que Povinelli et al. (2000) etiquetan 'física popular'”. El razonamiento causal reduce los costos de aprendizaje individual, haciendo que el aprendizaje de los individuos sea más eficiente, “al hacer que el aprendizaje individual sea razonado en lugar de aleatorio” (Vaesen, 2012, p.215). Vaesen también sostiene que el control ejecutivo que implica la inhibición de las impulsiones actuales y el foco de atención contribuye de manera similar.
Vaesen (2012) describe el trabajo de otros investigadores que han identificado áreas cerebrales involucradas en el uso de herramientas. Vaesen (p.204) escribe “Orban y sus colegas (2006) identificaron un conjunto de regiones funcionales en el surco intraaparietal dorsal (IPS) del cerebro humano que está involucrada en representaciones del campo visual central y en la extracción de la forma tridimensional del movimiento. Fundamentalmente, estas regiones cerebrales no se encontraron en los cerebros de los monos. Las regiones subsirven, conjeturaron los autores, el análisis visual mejorado necesario para la precisión con la que los humanos manipulan las herramientas. En segundo lugar, Stout y Chaminade (2007) encontraron que partes de estas regiones eran efectivamente reclutadas cuando los sujetos humanos modernos se dedicaban a la fabricación de herramientas similares a Oldowan. Es importante destacar que no se observó un aumento de la activación cuando se pidió a los sujetos humanos que golpearan adoquines juntos sin pretender producir escamas. El IPS dorsal humano, por lo tanto, puede permitir una mejor identificación de dianas adecuadas en el núcleo, y como tal, explicar en parte por qué los humanos superan a otros primates en materia de uso de herramientas”.
Los conocimientos adicionales sobre el uso de herramientas y el cerebro provienen del estudio de individuos con daño cerebral. Osiurak y Massen (2014, p. 1107) revisan los resultados de estudios de “pacientes con trastornos por uso de herramientas, también llamados apraxia de uso de herramientas. Cuando se les pide que enciendan una vela, por ejemplo, esos pacientes pueden encenderla correctamente pero luego se la ponen a la boca en un intento de fumarla. Tales observaciones han llevado a los modelos cognitivos tradicionales de apraxia a asumir que el uso de herramientas está respaldado por el conocimiento sensoriomotor sobre la manipulación de herramientas. Esta hipótesis sensitivo-motora asume que el conocimiento de manipulación almacenado dentro de las áreas fronto-parietales inferiores [de la corteza cerebral] es crítico para las habilidades de uso de herramientas”. Este puede ser el caso porque “el lóbulo parietal inferior izquierdo podría más bien apoyar el razonamiento técnico, es decir, la capacidad de razonar sobre las propiedades físicas del objeto... la capacidad de comprender las acciones mecánicas podría ser la especificidad de las porciones anteriores de la lóbulo parietal inferior (particularmente la circunvolución supramarginal) mientras que la corteza parietal posterior podría estar involucrada en la planeación de los componentes de agarre y alcance tanto de acciones de uso de herramientas como de no uso de herramientas”.
Figura\(\PageIndex{30}\): Corteza cerebral derecha humana resaltando partes del lóbulo parietal. Los pliegues en la superficie de la corteza forman colinas (giras, singulares, circunvoluciones) y valles (fisuras o surcos; singular, surco). El surco central divide el lóbulo frontal del lóbulo parietal detrás de él. El lóbulo parietal, especialmente en el lado derecho del cerebro, procesa información sobre el espacio y las relaciones espaciales. Las regiones del lóbulo parietal están involucradas en la construcción de herramientas y el uso de herramientas, importantes en la evolución humana. Ver texto para más detalles. (Imagen de Wikimedia Commons; Archivo:ParietCapts lateral.png; https://commons.wikimedia.org/wiki/F...ts_lateral.png; autor, Sebastian023; licenciado bajo la licencia Creative CommonsReconocimiento-Compartir Igual 3.0 Unported. Leyenda de Kenneth A. Koenigshofer, PhD).
La construcción y el uso de herramientas, aunque de importancia crítica en la evolución humana como lo indica la discusión anterior, son solo un aspecto estrecho del complejo de habilidades que conforman la inteligencia humana. En el Capítulo 14 sobre Inteligencia y Cognición, continuamos nuestra discusión sobre este tema examinando los mecanismos cerebrales que subyacen a otras habilidades cognitivas a las que colectivamente denominamos inteligencia. En la siguiente sección de este capítulo, David Buss de la Universidad de Texas examina las teorías evolutivas en psicología.
Anghelinu, M. (2013). Colocación de la escalera al árbol: Una visión de sentido común sobre la evolución cognitiva del linaje humano del Pleistoceno. Arheologia Moldovei, 36 (1), 7-24.
Bailey SE. (2006). Sahelanthropus tchadensis. En: Enciclopedia de antropología, Vol. 5. Thousand Oaks (CA): SAGE Referencia. p. 2044-2045.
Cachel, S. (2012). El comportamiento de las herramientas humanas es específico de cada especie y sigue siendo único. Ciencias del Comportamiento y del Cerebro, 35 (4), 222.
Callaway E. (2015). Los neandertales tuvieron un efecto enorme en la biología humana. Naturaleza [Internet] [citado el 21 de ago de 2015]; 523 (7562): 512-513. Disponible en: http://www.nature.com.offcampus.lib....iology-1.18086. doi: 10.1038/523512a
Cooper A, Stringer CB. (2013). ¿Los denisovanos cruzaron la Línea Wallace? Ciencia [Internet] [citado 2015 Ago 21}; 342 (6156): 321-323. Disponible en: http://www.sciencemag.org.offcampus....d-963a99b700ea. doi: 10.1126/ciencia.1244869.
Cosmides, L., & Tooby, J. (2002). Desentrañar el enigma de la inteligencia humana: La psicología evolutiva y la mente multimodular. La evolución de la inteligencia, 145-198.
Dembo M, Matzke NJ, Mooers AØ, Collard M. (2015). El análisis bayesiano de una supermatriz morfológica arroja luz sobre las controvertidas relaciones homínidos. Proc R Soc B [Internet] [citado el 20 de ago de 2015]; 282 (1812). Disponible en: http://rspb.royalsocietypublishing.o...0943.e-letters. doi: 10.1098/rspb.2015.0943.
Domínguez-Rodrigo M, Pickering TR, Baquedano E, Mabulla A, Mark DF, Musiba C, et al. (2013). Primer esqueleto parcial de un Paranthropus boisei de 1.34 millones de años de edad procedente del lecho II, Garganta de Olduvai, Tanzania. PLoS ONE 8 (12): e80347. Disponible en: http://journals.plos.org/plosone/art...l.pone.0080347. doi:10.1371/journal.pone.0080347
Dunbar, R. I. (1998). La hipótesis del cerebro social. Antropología evolutiva: temas, noticias y reseñas: números, noticias y reseñas, 6 (5), 178-190.
eFósiles [Internet] [citado 2015 ago 10]. Departamento de Antropología, La Universidad de Texas en Austin. Disponible en: http://efossils.org/
eLucy: Paso a paso: la evolución del bipedalismo [Internet]. c2007. Austin (TX): Departamento de Antropología, Universidad de Texas-Austin. [citado 2015 ago 3]. Disponible en: elucy.org/main/lessonoverview.html
Geertz, A. W. (2013). El cerebro significativo: Clifford Geertz y la ciencia cognitiva de la cultura. En: La cultura mental: la teoría social clásica y la ciencia cognitiva de la religión. Editado por Dimitris Xygalatas y William W. McCorkle Jr., 176-196.
Gibbons, A. (2012). Las brechas generacionales sugieren una división antigua humano-simio. Ciencia, 13 de agosto de 2012. Asociación Americana para el Avance de la Ciencia. https://www.science.org/content/arti...%20years%20ago. Recuperado 4 de julio de 2022.
Harmand S, Lewis JE, Feibel CS, Lepre CJ, Prat S, Lenoble A, Boës X, Quinn RL, Brenet M, Arroyo A, Taylor N, Clément S, Daver G, Brugal JP, Leakey L, Mortlock RA, Wright JD, Lokorodi S, Kirwa C, Kent DV, Roche H. 2015. Herramientas de 3.3 millones de años de Lomekwi 3, Turkana Occidental, Kenia. Naturaleza 521 (21 de mayo): 310-315. Disponible en: http://www.nature.com.offcampus.lib....ture14464.html. doi:10.1038/naturaleza14464.
Caza KD. Australopitecinos. En: Enciclopedia de antropología, Vol. 1. Thousand Oaks (CA): Referencia SAGE, 2006. p.311-317.
Higham T, Douka K, Wood R et al. (2014). El momento y el patrón espaciotemporal de la desaparición neandertal. Naturaleza [Internet] [citado el 21 de ago de 2015]; 512 (7514): 306-309. Disponible en: http://www.nature.com.offcampus.lib....ture13621.html. doi:10.1038/naturaleza13621
Jurmain R, Kilgore L, Trevathan W. (2013). Esenciales de la antropología física, 9ª edición. Belmont (CA): Aprendizaje de Wadsworth Cengage.
Koenigshofer, K. A. (2017). Inteligencia general: adaptación a invariantes relacionales abstractas evolutivamente familiares, no a la novedad ambiental o evolutiva. El diario de la mente y el comportamiento, 38 (2), 119-153.
Larsen, CS. (2014). Nuestros orígenes: descubrir la antropología física. Nueva York (NY): W. W. Norton & Company, Inc.
Lebatard AE, Bouriès DL, Duriner P, Jolivet M, Braucjer R, Carcaillet J, Schuster M, Arnaud N, Monié P, Lihoreau F, Likius A, Macaye HT, Vignaud P, Brunet M. 2008. Nuclido cosmogénico que data de Sahelanthropus tchadensis y Australopithecus bahrelghazali: Homínidos Mio-Pliecenos de Chad. Proc Natl Acad Sci U S A [Internet] [citado 2015 ago 3]; 105 (9): 3226-3231. Disponible en: http://www.pnas.org/content/105/9/3226.full. doi: 10.1073/pnas.0708015105
Piojos y evolución humana [Internet]. 2011 Feb 6. NovaCencienow. [citado 2015 Ago 20]. Disponible en: www.pbs.org/wgbh/nova/evolution/lice.html
Meyer M, Kircher M, Gansauge MT, et al. (2012). Secuencia genómica de alta cobertura de un individuo denisovano arcaico. Ciencia [Internet] [citado el 21 de ago de 2015]; 338 (6104): 222-226. Disponible en: http://www.sciencemag.org.offcampus..../6104/222.full. doi: 10.1126/ciencia.1224344.
O'Neil D. c1999-2012. Evolución humana temprana: un estudio de la evolución biológica y cultural de Homo habilis y Homo erectus. Departamento de Ciencias del Comportamiento, Colegio Palomar [Internet] [citado 2015 Ago 16]. Disponible en: Anthro.palomar.edu/Homo/Default.htm
O'Neil D. c1999-2014. Evolución de los humanos modernos: un estudio de la evolución biológica y cultural del Homo sapiens arcaico y moderno. Departamento de Ciencias del Comportamiento, Colegio Palomar [Internet] [citado 2015 Ago 16]. Disponible en: anthro.palomar.edu/homo2/default.htm
Osiurak, F., & Massen, C. (2014). Las bases cognitivas y neuronales del uso de herramientas humanas. Fronteras en psicología, 5, 1107.
Oxnard, C., Oberendorf, P.J., Kefford, B.J. (2010). Los esqueletos postcraneales de cretinas hipertiroideas muestran un mosaico anatómico similar al Homo floesiensis. PLoS One [Internet] [citado el 21 de ago de 2015]; 5 (9). Disponible en: http://www.sciencedaily.com/releases...0928025514.htm. doi: 10.1371/journal.pone.0013018.
Pääbo S. 2015. Los diversos orígenes del acervo genético humano. Nat Rev Genet [Internet] [citado 2015 Ago 21]; 16:313-314. Disponible en: http://www.nature.com.offcampus.lib....l/nrg3954.html. doi:10.1038/nrg3954
Reich D, Green RE, Kircher M, et al. (2010). Historia genética de un grupo arcaico de homínidos de la cueva Denisova en Siberia. Naturaleza [Internet] [citado el 21 de ago de 2015]; 468 (7327). Disponible en: http://www.nature.com/nature/journal...ture09710.html
Richmond BG, Jungers WL. (2008). Morfología femoral de Orrorin tugenensis y evolución del bipedalismo homínico. Ciencia [Internet] [citado 2015 ago 3]; 319 (5870): 1662-1665. Disponible en: www.jstor.org/stable/20053635
Scarre C. (2005). El pasado humano: la prehistoria mundial&El desarrollo de las sociedades humanas. Londres (Reino Unido): Thames & Hudson Ltd.
Scarre C. (2013). El pasado humano. Londres (Reino Unido): Thames & Hudson.
Su DF. (2013). Los primeros homínidos: Sahelanthropus, Orrorin y Ardipithecus. The Nature Education Knowledge Project [Internet] [citado 2015 Ago 3]. Disponible en: http://www.nature.com/scitable/knowl...hecus-67648286
Vaesen, K. (2012). Las bases cognitivas del uso de la herramienta humana. Ciencias del comportamiento y del cerebro, 35 (4), 203-218.
Ward CV, Manthi FK, Plavcan JM. (2013). Nuevos fósiles de Australopithecus anamensis de Kanapoi, Turkana Occidental, Kenia (2003-2008). J Hum Evol 65 (5): 501-524. Disponible desde doi:10.1016/j.jhevol.2013.05.006
Wong K. 2006. Reportaje especial: El bebé de Lucy: sale a la luz un extraordinario nuevo fósil humano. Sci Am [Internet] [citado el 10 de ago de 2015]; 20 de sep. Disponible en: http://www.scientificamerican.com/ar...rt-lucys-baby/
Madera, B. (2010). Reconstruir la evolución humana: logros, desafíos y oportunidades. Actas de la Academia Nacional de Ciencias, 107 (suplemento2), 8902-8909.
Revistas de prensa de la Universidad de Chicago [Internet]. Volcanes aniquilaron a los neandertales, sugiere un nuevo estudio. ScienceDaily 2010 Oct 7 [citado 2015 Ago 21]. Disponible en: http://www.sciencedaily.com/releases...1006094057.htm
Bullock, T. H. (1984). La neurociencia comparada es prometedora para revoluciones silenciosas. Ciencia, 225 (4661), 473—478.
Cachel, S. (2012). El comportamiento de las herramientas humanas es específico de cada especie y sigue siendo único. Ciencias del Comportamiento y del Cerebro, 35 (4), 222.
Crick, F., & Koch, C. (1990). Hacia una teoría neurobiológica de la conciencia. Seminarios en las Neurociencias, 2, 263—275.
Darwin, C. (1871). El descenso del hombre, y la selección en relación al sexo. Londres: J. Murray.
Darwin, C. (1859). Sobre los orígenes de las especies por medio de la selección natural, o, La preservación de razas favorecidas en la lucha por la vida. Londres, Reino Unido: J. Murray.
Demonet, J. F., Chollet, F., Ramsay, S., Cardebat, D., Nespoulous, J. L., Wise, R.,. Frackowiak, R. (1992). Anatomía del procesamiento fonológico y semántico en sujetos normales. Cerebro, 115 (6), 1753—1768.
Edelman, G. (2004). Más ancho que el cielo: El don fenomenal de la conciencia. New Haven, CT: Prensa de la Universidad de Yale.
Gardner, R. A., & Gardner, B. T. (1969). Enseñar el lenguaje de señas a un chimpancé. Ciencia, 165 (3894), 664—672.
Goodall, J. (1986). Los chimpancés de Gombe: Patrones de comportamiento. Cambridge, MA: Prensa de la Universidad de Harvard.
Hubel, D. H. (1995). Ojo, cerebro y visión. Freeman & Co., NY: Scientific American Library/Scientific American Books.
Knapp, M. L., & Hall, J. A. (2009). La comunicación no verbal en la interacción humana. Boston, MA: Wadsworth Cengage Learning.
Osiurak, F., & Massen, C. (2014). Las bases cognitivas y neuronales del uso de herramientas humanas. Fronteras en psicología, 5, 1107.
Ramón y Cahal, S. (1937). Recolecciones de mi vida. Memorias de la American Philosophical Society, 8, 1901—1917.
Rolls, E. T., & Cowey, A. (1970). Topografía de la retina y corteza estriada y su relación con la agudeza visual en monos rhesus y monos ardilla. Investigación Cerebral Experimental, 10 (3), 298—310.
Rosenzweig, M. R., Breedlove, S. M., & Leiman, A. L. (2002). Psicología biológica (3ª ed.). Sunderland, MA: Asociados Sinauer.
Smith, E. E., & Jonides, J. (1999). Almacenamiento y procesos ejecutivos en los lóbulos frontales. Ciencia, 283 (5408), 1657—1661.
Smith, E. E., & Jonides, J. (1997). Memoria de trabajo: Una visión desde la neuroimagen. Psicología cognitiva, 33, 5—42.
Van Essen, D. C., Anderson, C. H., & Felleman, D. J. (1992). Procesamiento de la información en el sistema visual de primates: Una perspectiva de sistemas integrados. Ciencia, 255 (5043), 419—423.
Walker, L. A. (1987). Una pérdida de palabras: La historia de la sordera en una familia. Nueva York, NY: Harper Perennial.
Atribuciones
Texto y figuras de “Evolución Humana” no atribuidas a otras fuentes adaptadas por Kenneth A. Koenigshofer, PhD, Chaffey College, de Antropología Biológica (Saneda y Campo), capítulo: Evolución humana; LibreTextos de Tori Saneda & Michelle Field, Profesores (Antropología) en Colegio Comunitario Cascadia (vía Wikieducador)
El texto y las figuras “Defining Hominins” no atribuidas a otras fuentes fueron adaptados y modificados por Kenneth A. Koenigshofer, Ph.D., Chaffey College, de Exploraciones: Una invitación abierta a la antropología biológica;https://explorations.americananthro.....php/chapters/; Capítulo 9: “Los primeros homínidos”, Editores: Beth Shook, Katie Nelson, Kelsie Aguilera y Lara Braff, American Anthropological Association Arlington, VA 2019; CC BY-NC 4.0 International, salvo que se indique otra cosa.
“Homo Genus”, “Definiendo el género Homo”, “Clima”, “Homo habilis: Los primeros miembros de nuestro género”, “Homo erectus”, “Homo erectus Culture”, “Uso de herramientas y habilidades cognitivas del Homo erectus” y “El panorama general del Homo temprano”, texto y figuras no atribuidas a otras fuentes adaptadas por Kenneth A. Koenigshofer , PhD, Chaffey College, de Exploraciones: una invitación abierta a la antropología biológica;https://explorations.americananthro.....php/chapters/; Capítulo 10, “Primeros miembros del género Homo” de Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, y Lara Braff, Eds.; licenciada bajo una Licencia Creative Commons Reconocimiento-NoComercial 4.0 Internacional, salvo que se indique otra cosa.
Cuadro 3.3: “Características resumidas del Homo habilis” adaptado por Kenneth A. Koenigshofer, PhD, Chaffey College, de Exploraciones: Una invitación abierta a la antropología biológica;https://explorations.americananthro.....php/chapters/; Capítulo 10, “Primeros miembros del género Homo” de Bonnie Yoshida-Levine Ph.D.; Beth Shook, Katie Nelson, Kelsie Aguilera, y Lara Braff, Eds.; licenciados bajo una Licencia Creative Commons Reconocimiento-NoComercial 4.0 Internacional, salvo que se indique otra cosa.


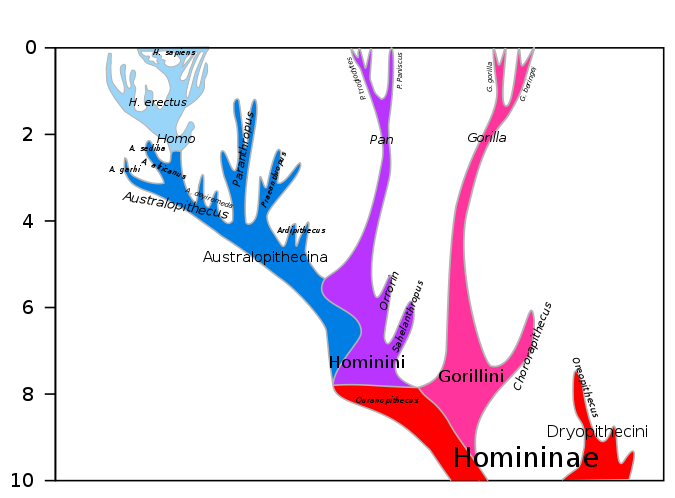
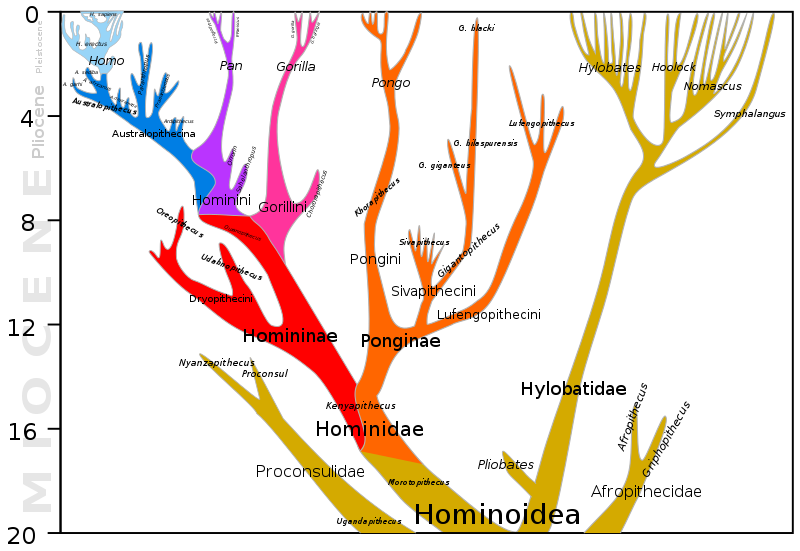


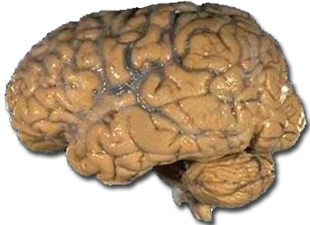
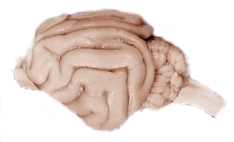

.jpg)
.jpg)

.jpg)
.jpg)