
3.2: Evolución - Mecanismos y Evidencia
- Page ID
- 148261
Objetivos de aprendizaje
- Describir cómo se desarrolló la teoría actual de la evolución.
- Explicar la selección natural y su papel en la evolución.
- Explicar la evolución convergente y divergente.
- Describir estructuras homólogas y vestigiales.
- Discutir conceptos erróneos sobre la teoría de la evolución.
- Explicar las categorías de evidencia para la evolución.
- Describir especies y especiaciones.
- Explicar la selección de parientes, la aptitud inclusiva y la evolución de comportamientos altruistas como ayudar, cooperar y dar.
Visión general
La teoría de la evolución es la teoría unificadora de la biología, es decir, es el marco dentro del cual los biólogos hacen preguntas sobre el mundo viviente. Su poder es que proporciona dirección para las predicciones sobre los seres vivos que se sustentan en experimento tras experimento. Recordemos la afirmación del genetista, Teodosio Dobzhansky, citada en la Sección 3.1, de que “nada tiene sentido en biología excepto a la luz de la evolución”. Quiso decir que el principio de que toda la vida ha evolucionado y diversificado a partir de un ancestro común es la base desde la que abordamos todas las cuestiones de la biología. Es importante tener en cuenta esta afirmación cuando pensamos en la evolución cerebral y la psicología. Los vínculos evolutivos entre las especies animales ayudan a explicar las similitudes en su cerebro y comportamiento.
Resumen de los puntos clave:
- Evolución: Todas las especies de organismos vivos, desde bacterias hasta babuinos y arándanos, evolucionaron en algún momento a partir de una especie diferente. Aunque pueda parecer que los seres vivos hoy en día siguen siendo muy iguales, ese no es el caso, la evolución es un proceso continuo.
- Comprender la evolución: La evolución por selección natural describe un mecanismo de cómo las especies cambian a lo largo del tiempo. Ese cambio de especie había sido sugerido y debatido mucho antes de que Darwin comenzara a explorar esta idea. La visión de que las especies eran estáticas e inmutables se basaba en los escritos de Platón, sin embargo, también había antiguos griegos que expresaban ideas evolutivas. Darwin fue el primero en describir el mecanismo principal del cambio evolutivo: la selección natural.
- Cuando Darwin propuso su teoría de la evolución, se desconocían los mecanismos de herencia. Ahora bien, sabemos mucho de esos mecanismos. Aquí hay algunas definiciones clave. Los cromosomas son largas cadenas de genes hechos de la molécula, ácido desoxirribonucleico (ADN). Un gen es un segmento de un cromosoma que codifica para la síntesis de una proteína específica. Los alelos son formas alternativas de un gen en la misma ubicación en el cromosoma (por ejemplo, un gen azul para el color de ojos y un gen marrón para el color de ojos son alelos). El genotipo es la composición genética de un individuo. El fenotipo es las características anatómicas, fisiológicas y comportamiento/psicológicas reales de un individuo.

Comprensión de la evolución
La evolución de Darwin por selección natural describe un mecanismo de cómo las especies cambian con el tiempo. Darwin describió la evolución como “descenso con modificación”.
En el siglo XVIII, James Hutton, naturalista escocés, propuso que el cambio geológico se producía gradualmente por la acumulación de pequeños cambios de procesos que operan como lo son hoy en día durante largos periodos de tiempo. Esto contrastaba con la visión predominante de que la geología del planeta era consecuencia de eventos catastróficos ocurridos durante un pasado relativamente breve. La visión de Hutton fue popularizada en el siglo XIX por el geólogo Charles Lyell quien se hizo amigo de Darwin. Las ideas de Lyell influyeron en el pensamiento de Darwin: la noción de Lyell de la mayor edad de la Tierra dio más tiempo para el cambio gradual en las especies, y el proceso de cambio proporcionó una analogía para el cambio gradual en las especies. A principios del siglo XIX, Jean-Baptiste Lamarck publicó un libro que detallaba un mecanismo para el cambio evolutivo. Este mecanismo se conoce ahora como una herencia de características adquiridas por la cual las modificaciones en un individuo que son causadas por su entorno, o el uso o desuso de una estructura durante su vida, podrían ser heredadas por su descendencia y así provocar cambios en una especie. Un simple ejemplo o dos ilustrarán por qué esto está mal; la idea sugiere que si vas al gimnasio y te pones aficionado, entonces tendrás hijos buff, o si pierdes un dedo en un accidente, que tendrás niños faltando un dedo, y que tales cambios eventualmente se convertirán en características de la especie. Hoy sabemos que si el cambio en las características no cambia los genes, el cambio no puede transmitirse a las generaciones futuras. Si bien este mecanismo para el cambio evolutivo fue desacreditado, las ideas de Lamarck fueron una influencia importante en el pensamiento evolutivo.
Charles Darwin y la selección natural
A mediados del siglo XIX, el mecanismo real de evolución fue concebido y descrito independientemente por dos naturalistas: Charles Darwin y Alfred Russel Wallace. Es importante destacar que cada naturalista dedicó tiempo a explorar el mundo natural en expediciones a los trópicos. De 1831 a 1836, Darwin viajó por todo el mundo en H.M.S. Beagle, incluyendo paradas en Sudamérica, Australia y el extremo sur de África. Wallace viajó a Brasil para recolectar insectos en la selva amazónica de 1848 a 1852 y al archipiélago malayo de 1854 a 1862. El viaje de Darwin, como los viajes posteriores de Wallace al archipiélago malayo, incluyó paradas en varias cadenas de islas, siendo la última las Islas Galápagos, al oeste de Ecuador. En estas islas, Darwin observó especies de organismos en diferentes islas que eran claramente similares, pero que tenían diferencias distintas. Por ejemplo, los pinzones terrestres que habitaban las islas Galápagos comprendían varias especies con una forma de pico única. Las especies en las islas tenían una serie graduada de tamaños y formas de pico con diferencias muy pequeñas entre las más similares. Observó que estos pinzones se parecían mucho a otra especie de pinzón en el continente de América del Sur. Darwin imaginó que las especies de la isla podrían ser especies modificadas de una de las especies originales del continente. Al estudiar más a fondo, se dio cuenta de que los variados picos de cada pinzón ayudaban a las aves a adquirir un tipo específico de alimento. Por ejemplo, los pinzones comedores de semillas tenían picos más fuertes y gruesos para romper las semillas, y los pinzones devoradores de insectos tenían picos en forma de lanza para apuñalar a sus presas.
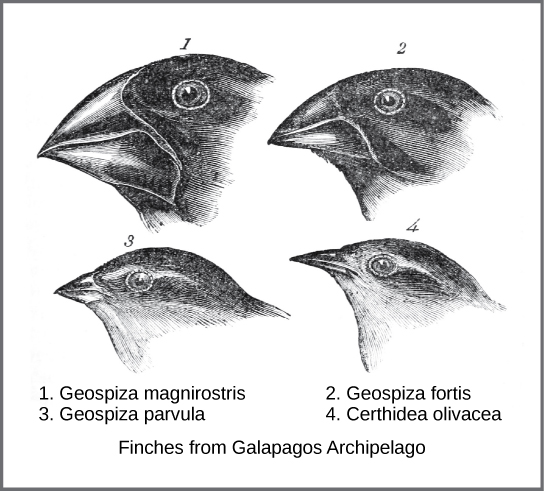
Wallace y Darwin observaron patrones similares en otros organismos y desarrollaron independientemente la misma explicación de cómo y por qué podrían ocurrir tales cambios. Darwin llamó a este mecanismo de selección natural. La selección natural, también conocida como “supervivencia del más apto”, se describe con mayor precisión como supervivencia y reproducción del más apto. La supervivencia por sí sola es insuficiente. La evolución implica tasas diferenciales de reproducción. La supervivencia sola sin reproducción no tiene ningún efecto sobre la evolución genética de una especie. La selección natural da como resultado la reproducción más prolífica de individuos con rasgos que contribuyen a la supervivencia y reproducción en un ambiente cambiante; esta mayor tasa de reproducción en individuos con rasgos que mejor se ajustan al ambiente en comparación con individuos con rasgos menos aptos conduce al cambio evolutivo.
Por ejemplo, Darwin observó que una población de tortugas gigantes encontradas en el archipiélago de Galápagos tenía cuellos más largos que los que vivían en otras islas con tierras bajas secas. Estas tortugas fueron “seleccionadas” porque podían llegar a más hojas y acceder a más alimentos que las de cuellos cortos. En tiempos de sequía en los que habría menos hojas disponibles, las que podían llegar a más hojas tenían más posibilidades de comer y sobrevivir que las que no podían llegar a la fuente de alimento. En consecuencia, las tortugas de cuello largo tendrían más probabilidades de tener éxito reproductivo y pasar el rasgo de cuello largo a sus crías. Con el tiempo, solo las tortugas de cuello largo estarían presentes en la población, ya que los animales de cuello corto no lograron sobrevivir y reproducirse.
La selección natural, argumentó Darwin, fue un resultado inevitable de tres principios que operaban en la naturaleza.
Primero, la mayoría de las características de los organismos se heredan, o se transmiten de padres a hijos. Aunque nadie, entre ellos Darwin y Wallace, sabía cómo sucedía esto en ese momento, era un entendimiento común.
Segundo, se producen más crías de las que son capaces de sobrevivir, por lo que los recursos para la supervivencia y reproducción son limitados. La capacidad de reproducción en todos los organismos supera la disponibilidad de recursos para apoyar su número. Así, hay competencia por esos recursos en cada generación. Tanto la comprensión de Darwin como Wallace de este principio provino de la lectura de un ensayo del economista Thomas Malthus quien discutió este principio en relación con las poblaciones humanas.
Tercero, las crías varían entre sí en cuanto a sus características y esas variaciones se heredan. Darwin y Wallace razonaron que las crías con características heredadas que les permitan competir mejor por recursos limitados sobrevivirán y tendrán más descendencia que aquellos individuos con variaciones que son menos capaces de competir. Debido a que las características son heredadas, estos rasgos estarán mejor representados en la próxima generación. Esto conducirá a un cambio en las poblaciones a lo largo de generaciones en un proceso que Darwin llamó descendencia con modificación. En última instancia, la selección natural conduce a una mayor adaptación de la población a su entorno local; es el único mecanismo conocido para la evolución adaptativa.
Los trabajos de Darwin y Wallace que presentaban la idea de la selección natural se leyeron juntos en 1858 antes de la Linnean Society de Londres. Al año siguiente se publicó el libro de Darwin, Sobre el origen de las especies. Su libro esbozó con considerable detalle sus argumentos a favor de la evolución por selección natural.

Las demostraciones de evolución por selección natural consumen mucho tiempo y son difíciles de obtener. Como se discutió brevemente en el Módulo 3.1, uno de los mejores ejemplos se ha demostrado en las mismas aves que ayudaron a inspirar la teoría de Darwin: los pinzones de Galápagos. Peter y Rosemary Grant y sus colegas han estudiado poblaciones de pinzones de Galápagos todos los años desde 1976 y han brindado importantes demostraciones de selección natural. Los Grants encontraron cambios de una generación a otra en la distribución de formas de pico en el pinzón de tierra mediana en la isla Galápagos de Daphne Major. Las aves han heredado variación en la forma del pico con algunas aves que tienen pico ancho profundo y otros tienen pico más delgado. Durante un periodo en el que las precipitaciones fueron superiores a lo normal debido a un El Niño (un inusual calentamiento del Océano Pacífico cerca de la costa oeste de las Américas), las grandes semillas duras que comieron las aves de pico grande se redujeron en número; sin embargo, hubo abundancia de las pequeñas semillas blandas que las pequeñas los pájaros comieron. Por lo tanto, la supervivencia y reproducción fueron mucho mejores en los años siguientes para las aves de pico pequeño. En los años siguientes a este El Niño, los Subvenciones midieron tamaños de pico en la población y encontraron que el tamaño promedio de los billetes era menor. Dado que el tamaño del pico es un rasgo heredado, los padres con billetes más pequeños tenían más descendencia y el tamaño de los billetes había evolucionado para ser más pequeño. A medida que cambiaron las condiciones y se produjo un largo período de sequía, las semillas más grandes se volvieron más disponibles. La tendencia hacia un tamaño promedio de pico más pequeño cesó y la selección de picos más grandes y tamaño corporal resultó en un aumento duradero en el tamaño del pico.
Como se discute en la Sección 3.1, otro ejemplo bien conocido de evolución observable es la polilla salpimentada. Por su importancia como otro ejemplo de selección natural en acción, revisamos los detalles aquí. Antes de la industrialización en Inglaterra, la polilla salpicada tenía alas de color claro que coincidían estrechamente con el color de la corteza de los árboles. Este rasgo heredado proporcionó camuflaje a las polillas ayudando a protegerlas de las aves depredadoras. Aunque la mayoría de las polillas tenían alas de color claro, algunas de cada generación tenían alas más oscuras, pero sin suficiente camuflaje estas polillas de color más oscuro habían sido presa fácil y sobrevivieron y reproducidas con baja frecuencia. A medida que la industrialización llegó a Inglaterra, el hollín de las fábricas comenzó a tornar lentamente la corteza de los árboles cada vez más oscura. Al ocurrir esto, las alas más oscuras eran ahora una ventaja, ayudando a ocultar las polillas más oscuras de los depredadores cuando aterrizaban en árboles, aumentando su supervivencia y tasas reproductivas sobre las polillas de color más claro. A medida que la selección natural continuó a lo largo de décadas, las polillas de color claro, alguna vez la forma predominante, fueron reemplazadas cada vez más a lo largo de generaciones por variantes aladas más oscuras, hasta hoy, después de no más de cien años de evolución, la polilla salpimentada tiene alas oscuras (aunque como se menciona en el Módulo 3.1 con limpiador aire, los números de la variante más ligera van aumentando a medida que la corteza del árbol continúa aclarándose). Esta transformación de la especie como consecuencia de la selección natural, a medida que cambió el ambiente, es un buen ejemplo de evolución lo suficientemente rápida como para que pueda observarse.
Proceso y Patrón de Evolución
La selección natural sólo puede realizarse si hay variación, o diferencias, entre los individuos de una población. Es importante destacar que estas diferencias deben tener alguna base genética; de lo contrario, la selección no conducirá a cambios en la próxima generación. Esto es crítico porque la variación entre los individuos puede ser causada por razones no genéticas, como que un individuo sea más alto debido a una mejor nutrición que a diferentes genes.
La diversidad genética en una población proviene de dos mecanismos principales: la mutación y la reproducción sexual. La mutación, un cambio en el ADN, es la fuente última de nuevos alelos (formas alternativas de un gen ubicado en el mismo lugar de un cromosoma), o nueva variación genética en cualquier población. Los cambios genéticos causados por la mutación pueden tener uno de tres resultados en el fenotipo. Una mutación afecta el fenotipo del organismo de una manera que le da forma reducida, menor probabilidad de supervivencia o menor descendencia. Una mutación puede producir un fenotipo con un efecto beneficioso sobre la condición física. Y, muchas mutaciones tampoco tendrán ningún efecto sobre la aptitud del fenotipo; estas se denominan mutaciones neutras. Las mutaciones también pueden tener toda una gama de tamaños de efecto sobre la aptitud del organismo que las expresa en su fenotipo, desde un pequeño efecto hasta un gran efecto. La reproducción sexual también conduce a la diversidad genética: cuando dos padres se reproducen, se ensamblan combinaciones únicas de alelos para producir los genotipos únicos y así los fenotipos en cada una de las crías.
Un rasgo heredable que ayuda a la supervivencia y reproducción de un organismo en su entorno actual se llama adaptación. Los científicos describen grupos de organismos que se adaptan a su entorno cuando se produce un cambio en el rango de variación genética con el tiempo que aumenta o mantiene el “ajuste” de la población a su entorno. Los pies palmeados de ornitorrinco son una adaptación para nadar. El grueso pelaje de los leopardos de las nieves es una adaptación para vivir en el frío. La rápida velocidad de los guepardos es una adaptación para atrapar presas.
El hecho de que un rasgo sea favorable o no depende de las condiciones ambientales en ese momento. No siempre se seleccionan los mismos rasgos porque las condiciones ambientales pueden cambiar. Por ejemplo, considere una especie de planta que creció en un clima húmedo y no necesitó conservar el agua. Se seleccionaron hojas grandes porque permitieron que la planta obtuviera más energía del sol. Las hojas grandes requieren más agua para mantener que las hojas pequeñas, y el ambiente húmedo proporcionó condiciones favorables para soportar hojas grandes. Después de miles de años, el clima cambió, y la zona ya no tenía exceso de agua. La dirección de la selección natural cambió de manera que se seleccionaron plantas con hojas pequeñas debido a que esas poblaciones pudieron conservar el agua para sobrevivir a las nuevas condiciones ambientales.
Formación de nuevas especies: Aunque toda la vida en la tierra comparte diversas similitudes genéticas, solo ciertos organismos combinan información genética por reproducción sexual y tienen descendencia que luego puede reproducirse con éxito. Los científicos llaman a tales organismos miembros de la misma especie biológica. Por ejemplo, una mula es un cruce entre un burro y un caballo. Sin embargo, dado que la mula no puede reproducirse (el burro y el caballo no tienen el mismo número de cromosomas), los burros y los caballos se consideran especies separadas. Sin embargo, un cruce entre un Dachshund y un Elkhound noruego puede resultar en crías que pueden reproducirse con otro perro doméstico, y así todos los perros domésticos son considerados de la misma especie (todos tienen el mismo número de cromosomas). La especiación (la formación de nuevas especies) ocurre en un lapso de tiempo evolutivo, por lo que cuando surge una nueva especie, hay un período de transición durante el cual las especies estrechamente relacionadas continúan interactuando.
La evolución de las especies ha dado lugar a una enorme variación en forma y función. En ocasiones, la evolución da lugar a grupos de organismos que se vuelven tremendamente diferentes entre sí. Cuando dos especies evolucionan en diversas direcciones desde un punto común, se llama evolución divergente. Tal evolución divergente se puede observar en las formas de los órganos reproductivos de las plantas con flores que comparten las mismas anatomías básicas; sin embargo, pueden verse muy diferentes como resultado de la selección en diferentes ambientes físicos y la adaptación a diferentes tipos de polinizadores. Como verá en la sección sobre la evolución humana, las líneas humanas y simios divergieron entre sí hace millones de años, sin embargo, la evidencia de una ascendencia común proviene de muchas fuentes, incluyendo similitudes en el ADN.

En otros casos, fenotipos similares evolucionan independientemente en especies distantes relacionadas. Por ejemplo, el vuelo ha evolucionado tanto en murciélagos como en insectos, y ambos tienen estructuras a las que nos referimos como alas, que son adaptaciones al vuelo. Sin embargo, las alas de murciélagos e insectos han evolucionado a partir de estructuras originales muy diferentes. Este fenómeno se denomina evolución convergente, donde rasgos similares evolucionan independientemente en especies que no comparten una ascendencia común reciente. Las dos especies llegaron a la misma función, volando, pero lo hicieron por separado una de la otra.
Selección Natural y Evolución Adaptativa
La selección natural solo actúa sobre los rasgos heredables de la población: seleccionar alelos beneficiosos y así aumentar su frecuencia en la población, al tiempo que selecciona contra alelos deletéreos y, por lo tanto, disminuye su frecuencia, un proceso conocido como evolución adaptativa. La selección natural no actúa sobre alelos individuales, sin embargo, sino sobre organismos enteros. Un individuo puede portar un genotipo muy beneficioso con un fenotipo resultante que, por ejemplo, aumenta la capacidad de reproducción (fecundidad), pero si ese mismo individuo también porta un alelo que da como resultado una enfermedad infantil fatal, ese fenotipo de fecundidad no se transmitirá a la siguiente generación porque el individuo no vivirá para llegar a la edad reproductiva. La selección natural actúa a nivel del individuo; selecciona para individuos con mayores contribuciones al acervo genético de la próxima generación, conocido como la aptitud evolutiva de un organismo (darwiniano).
La aptitud física suele ser cuantificable y es medida por científicos en el campo. Sin embargo, no es la aptitud absoluta de un individuo lo que cuenta, sino cómo se compara con los otros organismos de la población. Este concepto, llamado aptitud relativa, permite a los investigadores determinar qué individuos están aportando descendencia adicional a la siguiente generación, y así, cómo podría evolucionar la población.
Evidencia de Evolución
La evidencia de evolución es convincente y extensa. Al observar todos los niveles de organización en los sistemas vivos, los biólogos ven la firma de la evolución pasada y presente. Darwin dedicó gran parte de su libro, Sobre el origen de las especies, a identificar patrones en la naturaleza que fueran consistentes con la evolución, y desde Darwin, nuestra comprensión se ha vuelto cada vez más clara.
Fósiles
Los fósiles proporcionan evidencia sólida de que los organismos del pasado no son los mismos que los encontrados hoy en día, y los fósiles muestran una progresión de la evolución. Los científicos determinan la edad de los fósiles y los categorizan de todo el mundo para determinar cuándo vivían los organismos entre sí. El registro fósil resultante cuenta la historia del pasado y muestra la evolución de la forma a lo largo de millones de años. Por ejemplo, los científicos han recuperado registros altamente detallados que muestran la evolución de humanos y caballos.

Anatomía y Embriología
Otro tipo de evidencia para la evolución es la presencia de estructuras en organismos que comparten la misma forma básica. Por ejemplo, los huesos en los apéndices de un ser humano, perro, ave y ballena comparten la misma construcción general (Figura 18.1.618.1.6) resultante de su origen en los apéndices de un ancestro común. Con el tiempo, la evolución condujo a cambios en las formas y tamaños de estos huesos en diferentes especies, pero han mantenido el mismo diseño general. Los científicos llaman a estas partes sinónimos estructuras homólogas.
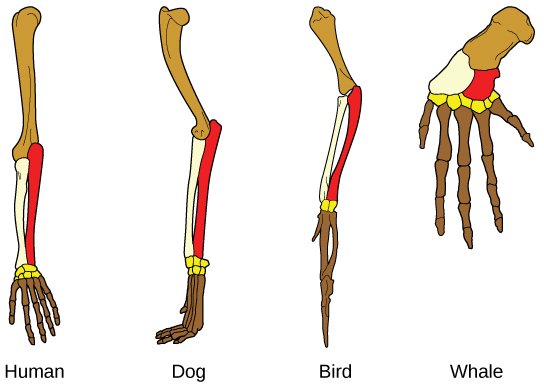
Algunas estructuras existen en organismos que no tienen ninguna función aparente en absoluto, y parecen ser partes residuales de un ancestro común pasado. Estas estructuras no utilizadas sin función se denominan estructuras vestigiales. Ejemplos de estructuras vestigiales son alas en aves no voladoras, hojas en algunos cactus y huesos de patas traseras en ballenas.
Visite este sitio interactivo para adivinar qué estructuras óseas son homólogas y cuáles son análogas, y ver ejemplos de adaptaciones evolutivas para ilustrar estos conceptos.
Otra evidencia de evolución es la convergencia de la forma en organismos que comparten ambientes similares. Por ejemplo, especies de animales no emparentados, como el zorro ártico y el ptarmigan, que viven en la región ártica, han sido seleccionadas para fenotipos blancos estacionales durante el invierno para mezclarse con la nieve y el hielo. Estas similitudes ocurren no por la ascendencia común, sino por presiones de selección similares, los beneficios de no ser vistos, por presas o depredadores, respectivamente.

La embriología, el estudio del desarrollo de la anatomía de un organismo a su forma adulta, también proporciona evidencia de relación entre grupos de organismos ahora ampliamente divergentes. El ajuste mutacional en el embrión puede tener consecuencias tan magnificadas en el adulto que la formación del embrión tiende a conservarse. En consecuencia, las estructuras que están ausentes en algunos grupos suelen aparecer en sus formas embrionarias y desaparecen cuando se alcanza la forma adulta o juvenil. Por ejemplo, todos los embriones vertebrados, incluidos los humanos, exhiben hendiduras branquiales y colas en algún momento de su desarrollo temprano. Estos desaparecen en los adultos de los grupos terrestres pero se mantienen en formas adultas de grupos acuáticos como peces y algunos anfibios. Los embriones de grandes simios, incluidos los humanos, tienen una estructura de cola durante su desarrollo que se pierde en el momento de (su) nacimiento.
Biogeografía
La distribución geográfica de los organismos en el planeta sigue patrones que mejor se explican por la evolución en conjunto con el movimiento de las placas tectónicas a lo largo del tiempo geológico. Amplios grupos que evolucionaron antes de la ruptura del supercontinente Pangea (hace unos 200 millones de años) se distribuyen a nivel mundial. Grupos que evolucionaron desde la ruptura aparecen de manera única en regiones del planeta, como la flora y fauna únicas de los continentes del norte que se formaron a partir del supercontinente Laurasia y de los continentes meridionales que se formaron a partir del supercontinente Gondwana. La presencia de miembros de la familia de plantas Proteaceae en Australia, el sur de África y Sudamérica se explica mejor por su presencia previa a la ruptura del supercontinente sur Gondwana.
La gran diversificación de marsupiales en Australia y la ausencia de otros mamíferos reflejan el largo aislamiento de Australia. Australia tiene una abundancia de especies endémicas, especies que no se encuentran en ninguna otra parte, lo que es típico de las islas cuyo aislamiento por extensiones de agua impide que las especies migren. Con el tiempo, estas especies divergen evolutivamente hacia nuevas especies que se ven muy diferentes a sus ancestros que pueden existir en el continente. Los marsupiales de Australia, los pinzones en las Galápagos y muchas especies en las islas hawaianas son todos únicos en su único punto de origen, pero muestran relaciones distantes con especies ancestrales en las tierras continentales.
Biología Molecular
Al igual que las estructuras anatómicas, las estructuras de las moléculas de la vida reflejan el “descenso con modificación” de Darwin (es decir, la evolución). La evidencia de un ancestro común para toda la vida se refleja en la universalidad del ADN como material genético y en la casi universalidad del código genético y la maquinaria de replicación y expresión del ADN. Las divisiones fundamentales en la vida entre los tres dominios (arquea-organismos unicelulares sin núcleo celular; bacterias; y eucariotas, incluyendo plantas, animales y hongos) se reflejan en grandes diferencias estructurales en estructuras conservadoras, como los componentes de los ribosomas y las estructuras de membranas. En general, la relación de grupos de organismos se refleja en la similitud de sus secuencias de ADN, exactamente el patrón que se esperaría de la descendencia y diversificación de un ancestro común.
Las secuencias de ADN también han arrojado luz sobre algunos de los mecanismos de la evolución. Por ejemplo, es claro que la evolución de nuevas funciones para las proteínas ocurre comúnmente después de eventos de duplicación génica que permiten la modificación libre de una copia por mutación, selección o deriva (cambios en el acervo genético de una población resultantes del azar), mientras que la segunda copia continúa produciendo un proteína.
Conceptos erróneos sobre la evolución
Aunque la teoría de la evolución generó cierta controversia cuando se propuso por primera vez, fue aceptada casi universalmente por los biólogos, particularmente los biólogos más jóvenes, dentro de los 20 años posteriores a la publicación de Sobre el origen de las especies. Sin embargo, la teoría de la evolución es un concepto difícil y abundan los conceptos erróneos sobre su funcionamiento.
Este sitio aborda algunos de los principales conceptos erróneos asociados a la teoría de la evolución.
La evolución es solo una teoría
Los críticos de la teoría de la evolución descartan su importancia al confundir a propósito el uso cotidiano de la palabra “teoría” con la forma en que los científicos usan la palabra. En la ciencia, una “teoría” se entiende como un cuerpo de explicaciones minuciosamente probadas y verificadas para un conjunto de observaciones del mundo natural. Los científicos tienen una teoría del átomo, una teoría de la gravedad y la teoría de la relatividad, cada una de las cuales describe hechos entendidos sobre el mundo. De la misma manera, la teoría de la evolución describe hechos sobre el mundo vivo. Como tal, una teoría en la ciencia ha sobrevivido a importantes esfuerzos por desacreditarla por parte de los científicos. En contraste, una “teoría” en lengua vernácula común es una palabra que significa una suposición o explicación sugerida; este significado es más parecido al concepto científico de “hipótesis”. Cuando los críticos de la evolución dicen que la evolución es “solo una teoría”, están dando a entender que hay poca evidencia que la apoye y que todavía está en proceso de ser rigurosamente probada. Esto es una caracterización errónea.
Los individuos evolucionan
La evolución es el cambio en la composición genética de una población a lo largo del tiempo, específicamente a lo largo de generaciones, resultante de la reproducción diferencial de individuos con ciertos alelos. Los individuos sí cambian a lo largo de su vida, obviamente, pero esto se llama desarrollo e implica cambios programados por el conjunto de genes que el individuo adquirió al nacer en coordinación con el entorno del individuo. Al pensar en la evolución de una característica, probablemente sea mejor pensar en el cambio del valor promedio de la característica en la población a lo largo del tiempo. Por ejemplo, cuando la selección natural conduce a un cambio del tamaño de la factura en los pinzones de tierra media en las Galápagos, esto no significa que las facturas individuales de los pinzones estén cambiando. Si uno mide el tamaño promedio de la factura entre todos los individuos de la población a la vez y luego mide el tamaño promedio de la factura en la población varios años después, este valor promedio será diferente como resultado de la evolución. Aunque algunos individuos pueden sobrevivir de la primera vez a la segunda, seguirán teniendo el mismo tamaño de factura; sin embargo, habrá muchos individuos nuevos que contribuyan al cambio en el tamaño promedio de la factura.
La evolución explica el origen de la vida
Es un malentendido común que la evolución incluye una explicación de los orígenes de la vida. Por el contrario, algunos críticos de la teoría creen que no puede explicar el origen de la vida. La teoría no trata de explicar el origen de la vida. La teoría de la evolución explica cómo las poblaciones cambian con el tiempo y cómo la vida diversifica el origen de las especies. No arroja luz sobre los inicios de la vida incluyendo los orígenes de las primeras células, que es como se define la vida. Los mecanismos del origen de la vida en la Tierra son un problema particularmente difícil porque ocurrió hace mucho tiempo, y presumiblemente solo ocurrió una vez. Es importante destacar que los biólogos creen que la presencia de vida en la Tierra excluye la posibilidad de que los eventos que llevaron a la vida en la Tierra puedan repetirse porque las etapas intermedias se convertirían inmediatamente en alimento para los seres vivos existentes.
Sin embargo, una vez que se implementara un mecanismo de herencia en forma de molécula como ADN, ya sea dentro de una célula o precélula, estas entidades estarían sujetas al principio de selección natural. Los reproductores más efectivos aumentarían en frecuencia a expensas de los reproductores ineficientes. Entonces, si bien la evolución no explica el origen de la vida, puede tener algo que decir sobre algunos de los procesos que operan una vez que las entidades previvientes adquirieron ciertas propiedades.
Los organismos evolucionan a propósito
Declaraciones como “los organismos evolucionan en respuesta a un cambio en un ambiente” son bastante comunes, pero tales afirmaciones pueden llevar a dos tipos de malentendidos. En primer lugar, la afirmación no debe entenderse en el sentido de que los organismos individuales evolucionan. El enunciado es la taquigrafía de “una población evoluciona en respuesta a un entorno cambiante”. Sin embargo, un segundo malentendido puede surgir al interpretar la afirmación en el sentido de que la evolución es de alguna manera intencional. Un ambiente cambiado da como resultado que algunos individuos en la población, aquellos con fenotipos particulares, beneficien y por lo tanto produzcan proporcionalmente más descendencia que otros fenotipos. Esto da como resultado un cambio en la población, si las características están determinadas genéticamente.
También es importante entender que la variación en la que trabaja la selección natural ya se encuentra en una población y no surge en respuesta a un cambio ambiental. Por ejemplo, aplicar antibióticos a una población de bacterias, con el tiempo, seleccionará una población de bacterias que sean resistentes a los antibióticos. La resistencia, que es causada por un gen, no surgió por mutación por la aplicación del antibiótico. El gen de resistencia ya estaba presente en el acervo genético de la bacteria, probablemente a baja frecuencia. El antibiótico, que mata las células bacterianas sin el gen de resistencia, selecciona fuertemente a los individuos que son resistentes, ya que estos serían los únicos que sobrevivieron y se dividieron. Los experimentos han demostrado que las mutaciones para la resistencia a los antibióticos no surgen como resultado del antibiótico.
En un sentido más amplio, la evolución no está dirigida a objetivos. Las especies no se vuelven “mejores” con el tiempo; simplemente rastrean su entorno cambiante con adaptaciones que maximizan su reproducción en un ambiente particular en un momento determinado. La evolución no tiene como objetivo hacer especies más rápidas, más grandes, más complejas o incluso más inteligentes, a pesar de lo común de este tipo de lenguaje en el discurso popular. Las características que evolucionan en una especie son una función de la variación presente y del entorno, los cuales cambian constantemente de manera no direccional. Qué rasgo encaja en un ambiente a la vez puede ser fatal en algún momento en el futuro. Esto se mantiene igual de bien para una especie de insecto que para la especie humana.
Evolución poblacional y síntesis moderna
Los mecanismos de herencia, o genética, no se entendieron en el momento en que Charles Darwin y Alfred Russel Wallace desarrollaban su idea de selección natural. Esta falta de comprensión fue un obstáculo para comprender muchos aspectos de la evolución. De hecho, la teoría genética predominante (e incorrecta) de la época, mezclando la herencia, dificultaba la comprensión de cómo podría operar la selección natural. Darwin y Wallace desconocían el trabajo genético del monje austriaco Gregor Mendel, que se publicó en 1866, poco después de la publicación del libro de Darwin, Sobre el origen de las especies (1859). La obra de Mendel fue redescubierta a principios del siglo XX, momento en el que los genetistas llegaban rápidamente a comprender los fundamentos de la herencia. Inicialmente, la naturaleza particulada recién descubierta de los genes dificultaba que los biólogos entendieran cómo podría ocurrir la evolución gradual. Pero en las próximas décadas, la genética y la evolución se integraron en lo que se conoció como la síntesis moderna, la comprensión coherente de la relación entre la selección natural y la genética que tomó forma en la década de 1940 y que generalmente se acepta hoy en día. En suma, la síntesis moderna describe cómo los procesos evolutivos, como la selección natural, pueden afectar la composición genética de una población y, a su vez, cómo esto puede resultar en la evolución gradual de poblaciones y especies. La teoría también conecta este cambio de una población a lo largo del tiempo, llamada microevolución, con los procesos de macroevolución que dieron lugar a nuevas especies y grupos taxonómicos superiores con caracteres ampliamente divergentes.
Selección Kin
En la discusión de la selección natural, se hizo énfasis en cómo funciona la selección natural en individuos para favorecer a los más aptos y desfavorecer a los menos aptos en una población. El énfasis estuvo en la supervivencia (selección de mortalidad), el éxito de apareamiento (selección sexual) o el tamaño de la familia (selección de fecundidad) de los individuos. Pero, ¿qué pasa con la abeja obrera que da la vida cuando el peligro amenaza su colmena? ¿O el pájaro madre que, fingiendo lesión, revolotea lejos de su nido de crías, arriesgando así la muerte a manos de un depredador? ¿Cómo puede la evolución producir genes para tales patrones instintivos de comportamiento cuando el dueño de estos genes corre el riesgo de fallar en la primera prueba de aptitud: supervivencia?
Una posible solución a este dilema radica en el efecto de tal comportamiento aparentemente altruista sobre la aptitud general (“inclusiva”) de la familia del individuo altruista. Vinculados entre sí por una dotación genética similar, el miembro altruista de la familia aumenta la posibilidad de que muchos de sus propios genes sean transmitidos a las generaciones futuras sacrificándose por el bienestar de sus familiares. Es interesante señalar que la mayoría de los comportamientos altruistas se observan donde los individuos están vinculados por lazos familiares bastante estrechos. La selección natural que trabaja a nivel de la familia y no del individuo se llama selección de parentesco.
¿Qué tan buena es la evidencia para la selección de familiares? ¿El comportamiento de la ave madre realmente aumenta sus posibilidades de ser asesinada? Después de todo, puede ser ventajoso tomar la iniciativa en un encuentro con un depredador que deambula cerca. Pero aunque sí aumente su riesgo, ¿esto es algo más que otro ejemplo de comportamiento materno? Sus hijos son, en efecto, sus familiares. Pero, ¿no es la selección natural simplemente operando de una de las maneras que Darwin describió: para producir familias maduras más grandes?
Quizás se encuentren ejemplos más claros de altruismo y selección de parientes en aquellas especies de aves que emplean “ayudantes”. Un ejemplo: arrendajos de matorral de Florida (Aphelocoma coerulescens coerulescens). Estas aves ocupan territorios bien definidos. Cuando alcanzan la madurez, muchas de las aves jóvenes permanecen por un tiempo (uno a cuatro años) en el territorio y ayudan a sus padres con la crianza de crías adicionales. ¡Qué abnegación! ¿No debería haber producido la selección natural un genotipo que lleve a sus dueños a buscar pareja y comenzar a criar a sus propias familias (para recibir esos genes)?
Pero la idea de selección de parientes sugiere que los genes que guían su comportamiento aparentemente altruista han sido seleccionados porque es más probable que se transmitan a las generaciones posteriores en los cuerpos de un mayor número de hermanos y hermanas menores que en los cuerpos de sus propios hijos. Para demostrar que esto es así, es necesario demostrar que:
- el comportamiento de “ayuda” de estas aves no apareadas es realmente una ayuda y que
- realmente han sacrificado oportunidades para ser padres exitosos ellos mismos.
Gracias a las observaciones de los pacientes de Glen Woolfenden, se establece el primer punto. Ha demostrado que los padres con ayudantes crían crías más grandes que las que no las tenían. Pero el segundo punto sigue sin resolverse. Quizás al esperar hasta que hayan adquirido experiencia guardando nidos y alimentando a los jóvenes y hasta que esté disponible un territorio adecuado, estos ayudantes aparentemente altruistas en realidad están mejorando sus posibilidades de criar familias más grandes de lo que tendrían si ellos comenzó justo en ello. Si es así, entonces una vez más simplemente estamos viendo la selección natural trabajando a través de uno de los criterios de aptitud individual de Darwin: la capacidad de producir familias maduras más grandes.
La ventaja evolutiva de ayudar cesa si los jóvenes no son en realidad hermanos del ayudante. Está bien establecido (por ejemplo, por análisis de ADN) que las hembras de muchas especies de aves tienen asuntos “extramatrimoniales”; es decir, producen crías donde las crías han sido engendradas por más de un macho. Curiosamente, resulta que cuanto más promiscuas sean las hembras de una especie dada, menos probable es que sean asistidas por ayudantes. Por el contrario, aquellas especies que emplean ayudantes tienden a ser monógamas (sin embargo, hay algunas excepciones).
Selección de Kin en Insectos Sociales
La abeja y otros insectos sociales proporcionan el ejemplo más claro de selección de familiares. También son ejemplos particularmente interesantes por las peculiares relaciones genéticas entre los miembros de la familia.
Las abejas melíferas macho (drones) se desarrollan a partir de los huevos no fertilizados de la reina y son haploides. Así, todos sus espermatozoides contendrán exactamente el mismo conjunto de genes. Esto significa que todas sus hijas compartirán exactamente el mismo conjunto de genes paternos, aunque compartirán —en promedio— sólo la mitad de los genes de su madre. (Las hermanas humanas, en contraste, comparten la mitad de los genes de su padre y la mitad de los genes de su madre). Por lo que cualquier comportamiento que favorezca a las hermanas abejas (75% de los genes compartidos) será más favorable a su genotipo que el comportamiento que favorezca a sus hijos (50% de los genes compartidos).
Ya que ese es el caso, ¿por qué molestarse con los niños en absoluto? ¿Por qué no hacer que la mayoría de las hermanas sean trabajadoras estériles, cuidando a su madre ya que cada vez produce más hermanas menores, algunas de las cuales algún día serán reinas? En cuanto a sus hermanos, las abejas obreras comparten solo el 25% de sus genes con ellos. ¿Es sorprendente, entonces, que a medida que se acerca el otoño, los trabajadores pierdan la paciencia con los perezosos y exigentes caminos de sus hermanos y finalmente los expulsen de la colmena?
Ningún organismo perfecto
La selección natural es una fuerza impulsora en la evolución y puede generar poblaciones que están mejor adaptadas para sobrevivir y reproducirse con éxito en sus entornos. Pero la selección natural no puede producir el organismo perfecto. La selección natural solo puede seleccionar sobre la variación existente en la población; no crea nada desde cero. Por lo tanto, está limitado por la varianza genética existente de una población y cualesquiera que sean los nuevos alelos (variantes genéticas) que surjan a través de la mutación y el flujo génico (cuando algunos organismos de una población migran a otra población).
La selección natural también es limitada porque funciona a nivel de individuos, no alelos, y algunos alelos están unidos entre sí debido a su proximidad física en el genoma, haciéndolos más propensos a transmitirse juntos (desequilibrio de ligamiento). Cualquier individuo dado puede portar algunos alelos beneficiosos y algunos alelos desfavorables. Es el efecto neto de estos alelos, o la aptitud del organismo, sobre el que puede actuar la selección natural. Como resultado, los alelos buenos se pueden perder si son portados por individuos que también tienen varios alelos abrumadoramente malos; de igual manera, los alelos malos se pueden mantener si son portados por individuos que tienen suficientes alelos buenos para resultar en un beneficio general de aptitud física.
Por último, es importante entender que no toda la evolución es adaptativa. Si bien la selección natural selecciona a los individuos más aptos y a menudo da como resultado una población más adecuada en general, otras fuerzas de evolución, incluyendo la deriva genética y el flujo génico, a menudo hacen lo contrario: introducir alelos perjudiciales al acervo genético de la población. La evolución no tiene propósito, no es transformar una población en un ideal preconcebido. Es simplemente la suma de las diversas fuerzas descritas en este capítulo y cómo influyen en la varianza genética y fenotípica de una población.
Resumen
La evolución es el proceso de adaptación a través de la mutación y la selección natural, lo que permite que las características mejor ajustadas biológicamente (es decir, mejor adaptadas) se pasen a las generaciones siguientes, mientras que las características menos adaptadas tienden a ser eliminadas. Con el tiempo, los organismos evolucionan características que son beneficiosas para su supervivencia y reproducción. Para que los organismos vivos se adapten y cambien a las presiones ambientales, la variación genética debe estar presente. Con la variación genética, los individuos tienen diferencias en forma y función que permiten que algunos sobrevivan mejor a las condiciones ambientales que otros. Estos organismos pasan sus rasgos favorables a sus crías. Eventualmente, los ambientes cambian, y lo que alguna vez fue un rasgo deseable y ventajoso puede convertirse en un rasgo indeseable y los organismos pueden evolucionar aún más. La evolución puede ser convergente con rasgos similares que evolucionan en múltiples especies o divergente con diversos rasgos que evolucionan en múltiples especies que provienen de un ancestro común. La evidencia de evolución se puede observar por medio del código de ADN y el registro fósil, y también por la existencia de estructuras homólogas y vestigiales.
La síntesis moderna de la teoría evolutiva surgió de la combinación de las formulaciones de evolución de Darwin y Wallace con el análisis de la herencia de Mendel, junto con el estudio más moderno de la genética de poblaciones. La síntesis moderna describe la evolución de poblaciones y especies, desde cambios a pequeña escala entre individuos hasta cambios a gran escala durante períodos paleontológicos. Para entender cómo evolucionan los organismos, los científicos pueden rastrear las frecuencias alélicas de las poblaciones a lo largo del tiempo.
La selección de kin y la aptitud inclusiva implican la selección de comportamientos altruistas que benefician a parientes genéticos cercanos.
Referencias
- Teodosio Dobzhansky. “Biología, Molecular y Orgánico”. Zoólogo Americano 4, núm. 4 (1964): 449.
Atribuciones
- Evolución: Mecanismos y Evidencias adaptados de Evolución y Origen de las Especies OpenStax, licenciado CC BY 4.0
- Selección Kin y Selección Kin en Insectos adaptados de Libretextos, Biología escrita por John W. Kimball. Este contenido se distribuye bajo una licencia Creative Commons Attribution 3.0 Unported (CC BY 3.0) y es posible gracias al financiamiento de The Saylor Foundation.