
5.1: Las neuronas y sus funciones básicas
- Page ID
- 148303
Objetivos de aprendizaje
- Describir la anatomía de las neuronas y la función de cada una de las tres partes principales de una neurona
- Describir neurotransmisores, sinapsis, vesículas sinápticas, potencial de reposo, EPSP, IPSP y potencial de acción
- Explicar la conducción salatoria y por qué es importante la comunicación dentro del sistema nervioso; incluir descripción de la vaina de mielina y los ganglios de Ranvier
- Describir el papel de los sitios receptores possinápticos en la comunicación entre neuronas
- Describir neuronas clasificadas por forma y por sus funciones
Visión general
En esta sección, continuamos nuestra exploración de las neuronas, los bloques de construcción del sistema nervioso. Examinamos cómo generan señales electroquímicas y cómo los miles de millones de neuronas en el sistema nervioso se comunican entre sí, un proceso conocido como transmisión sináptica. Antes de abordar estos temas, revisamos y ampliamos la anatomía básica y el funcionamiento de las neuronas cubiertas en parte en el Capítulo 4. Una comprensión sólida de estos hechos proporciona la base para comprender cómo se generan los potenciales neuronales dentro de las neuronas y cómo se combinan para desencadenar la transmisión sináptica. Al leer, recuerde que los voltajes y eventos químicos que discutimos en esta sección, operando en grandes poblaciones de células cerebrales, de alguna manera generan nuestras percepciones, pensamientos, emociones, y la totalidad de nuestra experiencia mental. Hasta la fecha, cómo sucede esto, cómo los patrones de potenciales neuronales en los circuitos cerebrales se convierten en mentes conscientes, sigue siendo el mayor misterio de todos los que enfrenta la ciencia moderna.
Introducción a la Anatomía Neurónica, Potenciales Neuronales y Transmisión Sináptica
El sistema nervioso de vertebrados como nosotros está compuesto por el sistema nervioso central, formado por el cerebro y la médula espinal, y el sistema nervioso periférico, que consiste en todos los nervios fuera del cerebro y la médula espinal.
Figura\(\PageIndex{1}\): El sistema nervioso humano y sus principales divisiones. Todos los componentes consisten en neuronas: células nerviosas. (NSDiagram NGB colors fixed.png adaptado por Naomi Bahm (colores en caja y fijos) de Archivo:NSDiagram.svg; https://commons.wikimedia.org/wiki/File:NSdiagram.svg; por Fuzzform vía Wikimedia Commons; Se concede permiso para copiar, distribuir y/o modificar este documento bajo los términos de la Licencia GNU Free Documentation, versión 1.2 o cualquier versión posterior publicada por la Free Software Foundation; sin Secciones Invariantes, sin Textos de Portada Frontal y sin Textos de Tapa Trasera. Una copia de la licencia se incluye en la sección titulada Licencia de Documentación Libre de GNU. Sujeto a descargos de responsabilidad).
El sistema nervioso periférico está compuesto por los nervios sensoriales somáticos (nervios de entrada como el nervio auditivo, el nervio óptico, los nervios sensoriales espinales que median las sensaciones cutáneas, etc.) y los nervios motores somáticos (nervios de salida, sirviendo a los músculos esqueléticos, activándolos o inhibiéndolos ) así como los nervios autonómicos sensoriales y motores (que sirven a los órganos internos del cuerpo como el corazón, los pulmones, los vasos sanguíneos, el sistema digestivo, las glándulas suprarrenales, etc.). Estos nervios motores autónomos, que activan o inhiben los órganos internos del cuerpo, son de dos tipos: el simpático (activar el corazón, los pulmones, constriñir los vasos sanguíneos, etc. para preparar al cuerpo para la “lucha o huida”) y parasimpático (conservar los recursos del cuerpo durante la no- situaciones de emergencia, no amenazantes) nervios autonómicos motores. Todas estas partes del sistema nervioso, incluyendo el cerebro, la médula espinal y el sistema nervioso periférico, están formadas por células nerviosas, técnicamente conocidas como neuronas. Se estima que el cerebro humano tiene entre 80 y 100 mil millones de neuronas. Recordemos del Capítulo 4 que las neuronas tienen tres partes principales: el soma o cuerpo celular, las dendritas que reciben entradas de otras neuronas, y el axón, que lleva la salida de las neuronas, los impulsos nerviosos (potenciales de acción), a otras neuronas, a los músculos o a las glándulas.
Probablemente hayas visto fotografías o dibujos de neuronas previamente en tu curso introductorio de psicología. Las siguientes imágenes muestran pequeñas secciones de la corteza cerebral en un adulto humano y un lactante humano. Observe la densidad de neuronas y conexiones en cada corte cerebral. La complejidad de las interconexiones entre neuronas es evidente en la imagen del extremo derecho que se basa en el uso de la tinción de Golgi.
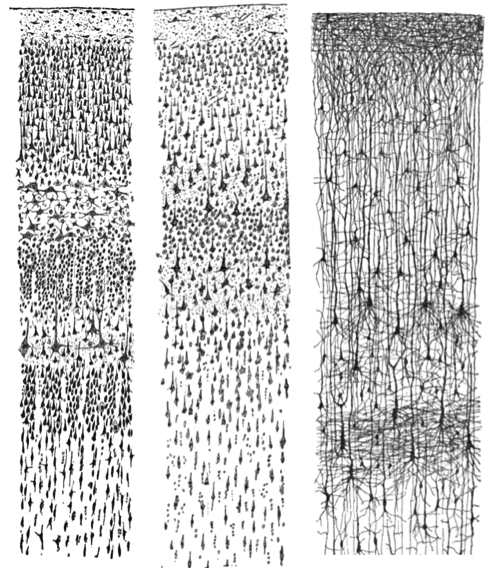
Figura\(\PageIndex{2}\): Pequeñas secciones de tejido del cerebro de un lactante humano y un adulto humano. La imagen en el extremo derecho con tinción de Golgi resalta cuerpos celulares y ramificaciones extensas de dendritas. Izquierda: Corteza visual teñida con NisSL de un adulto humano. Medio: corteza motora teñida de NisSL de un adulto humano. Derecha: Corteza teñida con Golgi de un lactante de 1 1/2 mes de edad. [Imagen: Santiago Ramon y Cajal, https://goo.gl/zOb2l1, CC0 Dominio Público, https://goo.gl/m25gce]
Manchas Nissl y Golgi
Obsérvese que se utilizaron diferentes químicos para teñir las neuronas en la figura anterior para que pudieran verse bajo el microscopio. La tinción Nissl marca solo la parte principal de la célula (es decir, el cuerpo celular; ver los paneles izquierdo y medio de la Figura 5.1.2). Sin embargo, por el contrario, una tinción de Golgi llena el cuerpo celular y todos los procesos que se extienden hacia afuera desde el cuerpo celular (ver panel derecho de la Figura 5.1.2). Una característica más notable de una tinción de Golgi es que solo tiñe aproximadamente el 1-2% de las neuronas (Pasternak & Woolsey, 1975; Smit & Colon, 1969), lo que permite al observador distinguir una célula de otra. Estas cualidades de la tinción de Golgi permitieron al neuroanatomista pionero Santiago Ramón y Cajal examinar por primera vez la estructura anatómica completa de las neuronas individuales. Esto mejoró significativamente nuestra apreciación de las intrincadas redes que forman sus procesos. Basándose en su observación del tejido teñido con Golgi, Cajal sugirió que las neuronas eran unidades de procesamiento distinguibles en lugar de formar parte de una red continua de nervios, como se había creído anteriormente. Cajal y Golgi compartieron el Premio Nobel de Medicina en 1906 por su contribución combinada al avance de la ciencia y nuestra comprensión de la estructura del sistema nervioso.
Anatomía de las neuronas y funciones básicas
Para entender la función de las neuronas, es importante conocer la anatomía involucrada. Como se señaló anteriormente, las neuronas tienen tres partes estructurales principales: el soma o cuerpo celular, el axón (lleva la salida de la neurona) y las dendritas (los “receptores” de la neurona). Toda la neurona está delimitada por una membrana celular, la membrana neural. La membrana celular de una neurona tiene canales o “puertas” para iones (átomos cargados eléctricamente) que pueden pasar a través de la membrana cuando se abren canales específicos para iones específicos. La Figura 5.1.4 muestra esta anatomía básica de la neurona. A continuación se discuten diversos tipos de neuronas.
El soma o cuerpo celular contiene orgánulos, comunes a todo tipo de células del cuerpo. Estos están involucrados en el metabolismo básico de la célula. El soma también contiene el núcleo, donde se localizan los genes y cromosomas (que contienen ADN).
La segunda parte principal de la neurona son las dendritas, los receptores de la neurona. Las dendritas en algunas neuronas pueden ramificarse profusamente (un gran número de ramas dendríticas del eje principal de una dendrita con sus propias ramas a menudo se denominan colectivamente árboles dendríticos), expandiendo la región de la neurona que puede recibir entradas de otras neuronas. Los sitios receptores (o más técnicamente, los sitios receptores postsinápticos debido a su ubicación en la neurona receptora o postsináptica) que reciben moléculas de neurotransmisor se localizan en las dendritas (y, en menor grado, en el soma) de la neurona receptora. En las dendritas se encuentran pequeñas espinas dendríticas que se asocian con las conexiones entre neuronas (las sinapsis) y pueden cambiar de forma rápidamente cuando se produce el aprendizaje (ver Capítulo 10 para más detalles). Tenga en cuenta que las espinas no son lo mismo que las ramas dendríticas. En la Figura 5.1.3, una rama dendrítica con espinas dendríticas se muestra a la izquierda en detalle microscópico, y a la derecha, son árboles dendríticos de dos tipos de neuronas que se encuentran en la retina. Las espinas no son visibles en las imágenes de árboles dendríticos de la derecha porque las espinas dendríticas son demasiado pequeñas, mientras que las ramas dendríticas que comprenden los árboles dendríticos se pueden ver fácilmente (ver subtítulo para la Figura 5.1.3).

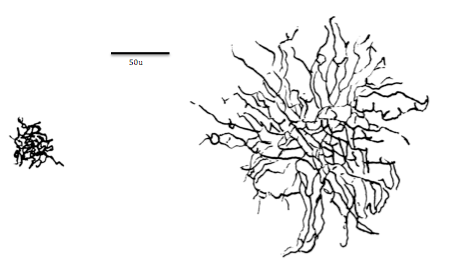
Figura\(\PageIndex{3}\): (Izquierda) Un segmento de dendrita de células piramidales del estrato radiatum (CA1) del hipocampo con espinas delgadas, regordetas y en forma de hongo. Las sinapsis de la columna vertebral están coloreadas en rojo, las sinapsis del tallo (o tallo) están coloreadas en azul. La dendrita se hizo transparente en la imagen inferior para permitir la visualización de todas las sinapsis. (Derecha) Comparación de tamaños entre árboles dendríticos de células enanas y sombrillas. Las espinas dendríticas son demasiado pequeñas para ser visibles en las dos imágenes de la derecha. Las neuronas sombrillas y enanas se encuentran en la capa celular ganglionar de la retina. (A la izquierda, dendrita y espinas de células piramidales: Imagen y subtítulo de Wikimedia Commons; Archivo:Un segmento de dendrita de células piramidales del estrato radiatum (CA1) .jpg; https://commons.wikimedia.org/wiki/F...atum_(CA1).jpg; por Synapse Web, Kristen M. Harris, PI, http://synapses.clm.utexas.edu/ ; licenciado bajo licencia Creative Commons Attribution 2.0 Genérica. A la derecha, árboles dendríticos celulares enanos y sombrillas: Imagen y leyenda de una línea de Wikimedia Commons; Archivo:Midget vs Parasol cell.png; https://commons.wikimedia.org/wiki/F...rasol_cell.png; por Stromdabomb; licenciado bajo la Creative Commons Atribución-Compartir Igual 4.0 Licencia internacional; explicación de dos oraciones por Kenneth A. Koenigshofer, PhD).
La tercera mayor parte de la neurona es el axón, saliendo del soma como una manguera. El axón lleva los mensajes de salida de una neurona (impulsos nerviosos) a lo largo de su longitud hasta los botones terminales del axón (terminaciones axónicas). Solo hay un axón por neurona, aunque puede ramificarse en múltiples botones terminales axónicos. En una neurona típica, el extremo de la raíz del axón emerge del soma en una pequeña hinchazón llamada montículo del axón. Entre el montículo del axón y el primer segmento del axón es donde primero se genera el impulso nervioso (ver discusión del potencial de acción, el impulso nervioso, que sigue a continuación).

Figura\(\PageIndex{4}\): Estructura básica de una neurona (Imagen de Wikimedia Commons; Archivo:Componentes de neuron.jpg; https://commons.wikimedia.org/wiki/F... _of_neuron.jpg; por Jennifer Walinga; licenciado bajo la licencia Creative Commons Reconocimiento-Compartir Igual 4.0 Internacional).
Axones Mielinizados y Conducción Saltatoria
Algunos axones tienen una cubierta de células gliales (las gliales son células no neuronales del sistema nervioso, ver Capítulo 4.1) conocida como vaina de mielina. Esta vaina de mielina grasa, aislante, tiene huecos en ella, revelando el axón desnudo, a intervalos regulares a lo largo del axón. Estas manchas desnudas a lo largo de la longitud de un axón mielinado se llaman nódulos, o nodos de Ranvier, después de su descubridor.
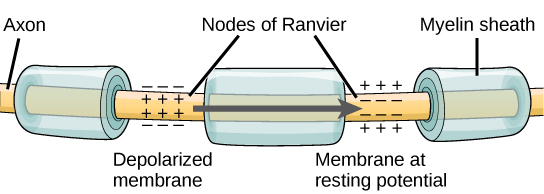
Figura\(\PageIndex{5}\): Nodos de Ranvier. Los nódulos de Ranvier son huecos en la vaina de mielina que cubre los axones mielinizados. Los nodos contienen canales de potasio (K+) y sodio (Na+) regulados por voltaje. Los potenciales de acción viajan por el axón “saltando” de un nodo al siguiente, acelerando la conducción del potencial de acción a lo largo del axón hacia el final del axón, también conocido como el botón axón, botón axón o terminal del axón.
La función de la vaina de mielina y los nódulos es acelerar la velocidad a la que los impulsos nerviosos viajan a lo largo del axón hacia su destino, la terminación del axón (axón bouton). En los axones mielinizados, los impulsos “saltan” de nodo a nodo permitiendo que el potencial de acción se mueva más rápidamente hacia abajo del axón. Este salto del impulso nervioso (potencial de acción) de nodo a nodo se llama conducción saltatoria, del latín “saltatore” que significa bailar. Imagina la imagen romántica del impulso bailando de nodo a nodo.
Un nodo de Ranvier es un hueco natural en la vaina de mielina a lo largo del axón. Estos espacios no mielinizados son de aproximadamente un micrómetro de largo y contienen canales iónicos de sodio (Na +) y potasio (K +) regulados por voltaje (es decir, abiertos por voltaje) (los iones son átomos cargados eléctricamente). El flujo de iones a través de estos canales, particularmente los canales de Na +, regenera el potencial de acción una y otra vez a lo largo del axón en cada nodo sucesivo de Ranvier. Como se señaló anteriormente, el potencial de acción “salta” de un nodo al siguiente en la conducción saltatoria. Si los nodos de Ranvier no están presentes a lo largo de un axón (como es el caso en los axones no mielinizados; ver más abajo), el potencial de acción se propaga mucho más lentamente; el movimiento iónico a través de los canales Na + y K + regenera continuamente nuevos potenciales de acción en cada punto sucesivo a lo largo del axón, utilizando tiempo extra para hacerlo. En efecto, la conducción del potencial de acción a lo largo de un axón implica canales regulados por voltaje (canales que responden al voltaje, más que al neurotransmisor), en el axón, respondiendo a la tensión que surge de un campo eléctrico que se extiende desde el potencial de acción en el segmento anterior del axón. En un axón mielinado, debido a que el potencial de acción salta de nodo a nodo, no tiene que regenerarse en cada punto sucesivo a lo largo del axón, sino solo en los nodos, saltando a través de segmentos mielinizados del axón entre nodos. Debido a que el potencial de acción en un axón mielinado debe regenerarse menos veces para moverse cualquier distancia particular a lo largo de la longitud del axón, alcanza su destino más rápido, en comparación con la velocidad de conducción en un axón no mielinizado.
Los nodos de Ranvier también ahorran energía para la neurona ya que los canales iónicos solo necesitan estar presentes y abiertos y cerrados en los nodos y no a lo largo de todo el axón. Es extraordinario que los nodos se coloquen a lo largo del axón en los intervalos espaciales correctos para hacer que la conducción de impulsos hacia abajo del axón sea lo más eficiente y rápida posible. Uno solo puede preguntarse la increíble precisión con la que la selección natural operó sobre esta característica de los axones mielinizados a lo largo del largo curso de la evolución animal para crear este espaciamiento óptimo de los nodos.
Conducción Neural en Axones No Mielinizados
No todos los axones están mielinizados. Los axones no mielinizados tienden a ser más antiguos en evolución y a ser los axones de menor diámetro (clasificados como fibras C en función de sus diámetros pequeños; los axones mielinizados de gran diámetro se denominan fibras A). En los axones no mielinizados, para poder moverse, el impulso nervioso debe regenerarse en cada punto sucesivo a lo largo del axón. Esto lleva tiempo y ralentiza la conducción del impulso nervioso (el potencial de acción) a lo largo del axón. Por lo tanto, la conducción del potencial de acción a lo largo de un axón no mielinizado es relativamente lenta. Un ejemplo de fibras C no mielinizadas son los axones que forman parte de las vías lentas del dolor. Estas vías median el dolor dolorido más lento que sigue al daño tisular. El dolor rápido y agudo de una lesión está mediado por fibras A de mayor diámetro (axones).
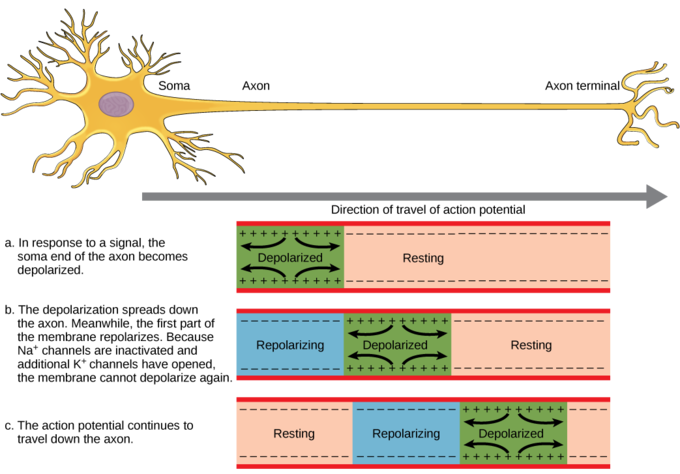
Figura\(\PageIndex{6}\): Potencial de acción viajando a lo largo de un axón neuronal no mielinizado. El potencial de acción se conduce hacia abajo del axón a medida que la membrana del axón se despolariza y luego se vuelve a polarizar. Debido a estas dinámicas, el potencial de acción solo se puede conducir en una dirección, lejos del cuerpo celular. Imagen: figure-35-02-04_NGB_added_resting.png adaptada por Naomi Bahm (descanso agregado a la parte a) de https://s3-us-west-2.amazonaws.com/c...e-35-02-04.png
Los canales iónicos a lo largo de la longitud del axón no mielinizado se denominan canales regulados por voltaje porque el voltaje generado por el potencial de acción en el segmento anterior del axón desencadena la apertura de estos canales en el siguiente segmento. Esto lleva a la regeneración del potencial de acción. A medida que este proceso se repite una y otra vez a lo largo de la longitud del axón no mielinado, el potencial de acción se lleva a cabo a lo largo de la longitud del axón lejos del cuerpo celular hacia las terminaciones axónicas de la neurona. La figura\(\PageIndex{7}\) muestra la apertura y cierre de los canales y los iones que se mueven a través de la membrana celular generando un potencial de acción en segmentos sucesivos del axón. Esto provoca el movimiento del potencial de acción, su conducción, a lo largo del axón alejándose del cuerpo celular (de derecha a izquierda en la figura, como muestra la flecha).
Figura\(\PageIndex{7}\): Otra representación de la propagación del impulso nervioso (el potencial axónico) a lo largo de un axón. Esta animación ilustra la propagación del potencial de acción en un axón. Se muestran tres tipos de canales iónicos: canales de “fuga” de potasio (azul), canales de sodio regulados por voltaje (rojo) y canales de potasio regulados por voltaje (verde). El movimiento de iones de sodio y potasio cargados de manera positiva a través de estos canales iónicos controla el potencial de membrana del axón. Los potenciales de acción se inician en el segmento inicial del axón después de que el neurotransmisor activa los receptores excitadores en las dendritas de la neurona y el cuerpo celular. Esto despolariza el segmento inicial del axón al voltaje umbral para la apertura de canales de sodio regulados por voltaje. Los iones de sodio que ingresan a través de los canales de sodio desplazan el potencial de la membrana al interior positivo El voltaje interno positivo durante el potencial de acción en el segmento inicial hace que la parte adyacente de la membrana del axón alcance el voltaje umbral. Cuando se alcanzan los potenciales positivos dentro de la membrana, los canales de potasio activados por voltaje se abren y los canales de sodio de puerta de voltaje se cierran. Los iones de potasio que salen del axón a través de canales de potasio regulados por voltaje devuelven el potencial de la membrana a valores negativos internos. Cuando se cierra la puerta de los canales de potasio regulados por voltaje, el potencial de la membrana vuelve al potencial de reposo. (Imagen y subtítulo de Wkimedia Commons; Archivo:Acción potencial propagación animation.gif; https://commons.wikimedia.org/wiki/F... _animation.gif; por John Schmidt; licenciado bajo la licencia Creative Commons Reconocimiento-Compartir Igual 4.0 Internacional, 3.0 Unported, 2.5 Genérica, 2.0 Genérica y 1.0 Genérica).
Por el contrario, como se describió anteriormente, en los axones mielinizados, el impulso se regenera solo en las manchas desnudas del axón, los nódulos de Ranvier. Debido a que el potencial de acción se regenera menos veces para recorrer una distancia determinada, que en el caso de los axones no mielinizados, la conducción neural es más rápida con la vaina aislante de mielina. Los axones mielinizados tienden a ser de gran diámetro (fibras A y B) y se encuentran en vías neurales mediando una respuesta conductual rápida, como el tracto piramidal que va desde la corteza motora a las neuronas motoras de la médula espinal que generan acción voluntaria.
Sinapsis
El axón se divide muchas veces, de manera que puede comunicarse, o sinapsis, con varias otras neuronas (ver Figura 5.1.4). Al final del axón, el botón terminal del axón (también llamado la terminación del axón, el ojal terminal, o el axón terminal) forma sinapsis con las espinas dendríticas, pequeñas protuberancias mencionadas anteriormente, en las dendritas de las neuronas receptoras (neuronas postsinápticas ). Se forman sinapsis entre el botón axón terminal de la neurona presináptica (neurona que envía la señal) y la membrana postsináptica (membrana de la neurona que recibe la señal; ver Figura 5.1.4). Aquí nos centraremos específicamente en las sinapsis entre el botón terminal axón de un axón y una columna dendrítica; sin embargo, las sinapsis también se pueden formar entre el botón terminal axón de la neurona presináptica y el soma de la neurona postsináptica, el eje dendrítico directamente, o el axón de otra neurona.
Existe un espacio muy pequeño llamado brecha sináptica o hendidura sináptica entre el botón terminal del axón presináptico y la columna dendrítica de la neurona possináptica. Para darte una idea del tamaño de la brecha sináptica, una moneda de diez centavos tiene un grosor de 1.35 mm (milímetro). Hay 1,350,000 nm (nanómetros) en el grosor de una moneda de diez centavos. La brecha sináptica es de solo unos 5 nm de ancho. En el botón terminal presináptico, hay vesículas sinápticas que agrupan grupos de químicos, neurotransmisores (ver Figura 5.1.4). Los neurotransmisores se liberan del botón terminal del axón presináptico o axón que termina en la brecha sináptica; las moléculas del neurotransmisor viajan a través de la brecha sináptica y abren canales iónicos en la columna postsináptica uniéndose a los sitios receptores allí. Discutiremos el papel de estos receptores con más detalle más adelante en la sección 5.2.
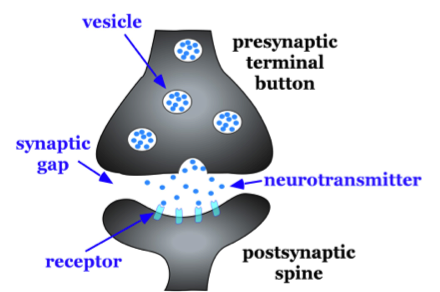
Figura\(\PageIndex{8}\): Características básicas de una sinapsis típica. Agrandamiento de la sinapsis entre uno de los botones terminales del axón (marcado botón terminal presináptico) y una de las dendritas de la segunda neurona que se muestran (a la derecha) en la Figura 5.1.4 anterior.
Botones terminales de axón, vesículas sinápticas, PSP y transmisión sináptica
Recordemos que al final de una rama de axón o axón se le llama botón terminal axón (o terminal, o simplemente el final del axón). Dentro del botón terminal del axón se encuentran estructuras llamadas vesículas sinápticas, que contienen sustancias químicas neurotransmisoras (las cuales han sido fabricadas en el soma y transportadas a la terminación del axón y almacenadas allí en las vesículas sinápticas, listas para su liberación). Cuando las neuronas se comunican entre sí a través de la brecha sináptica que las separa, es el neurotransmisor, liberado de las vesículas sinápticas en el botón terminal axón de la neurona “emisora” (la neurona presináptica) el que transmite el mensaje neural a través de la brecha sináptica (el espacio entre la membrana del botón axón terminal de la neurona presináptica y la membrana de la dendrita o soma de la neurona postsináptica). El evento que desencadena la liberación del neurotransmisor de las vesículas sinápticas en el botón terminal del axón es la llegada de un potencial de acción al botón terminal axón de la neurona emisora. La célula nerviosa que libera el neurotransmisor, la neurona emisora, se conoce técnicamente como la neurona presináptica. La neurona que recibe el neurotransmisor, la célula receptora, se llama neurona postsináptica.
Figura\(\PageIndex{9}\): Una sinapsis. Las vesículas sinápticas liberan neurotransmisores (pequeñas bolas amarillas) que se unen a los receptores (estructuras tipo peg azul) en la membrana postsináptica. Vesículas sinápticas dentro de un botón terminal axónico presináptico (terminación axónica) liberando moléculas neurotransmisoras sobre receptores en una dendrita de una neurona receptora (possináptica). (Imagen de Wikimedia Commons; Archivo:Neurotransmitters.jpg; https://commons.wikimedia.org/wiki/F...ansmitters.jpg; por https://www.scientificanimations.com/; por https://www.scientificanimations.com/; con licencia Creative Commons Atribución-Compartir Igual 4.0 Licencia internacional).
Las moléculas neurotransmisoras se adhieren a sitios especiales en la membrana de la neurona postsináptica. Estos sitios especiales se denominan sitios receptores o sitios receptores postsinápticos y normalmente se localizan en las dendritas y espinas dendríticas de la neurona receptora, pero también pueden estar en su soma. Sus formas moleculares coinciden con las formas de las moléculas neurotransmisoras que reciben, una especie de ajuste de “cerradura y llave”, que abre canales iónicos regulados químicamente permitiendo que los iones con sus cargas eléctricas se muevan a través de la membrana celular creando cambios de voltaje llamados potenciales postsinápticos (PSP; los detalles adicionales siguen en la Sección 5.2).
Estos eventos desde la liberación del transmisor hasta la generación de potenciales possinápticos comprenden la transmisión sináptica. Muchas drogas psicoactivas (drogas que alteran la mente y/o el comportamiento) como anfetaminas, LSD, “hongos mágicos”, etc. producen sus efectos bloqueando o activando sitios receptores específicos (otros psicoactivos producen sus efectos por otros mecanismos que afectan la transmisión sináptica, algunas de las cuales se discuten a continuación y en el capítulo 6 de este libro de texto).
Como se mencionó anteriormente, la membrana celular de una neurona tiene canales o “puertas” para iones (átomos cargados eléctricamente) que pueden pasar a través de la membrana cuando se abren canales específicos para iones específicos. Normalmente los canales se cierran hasta que actúan sobre ellos mediante la unión de moléculas neurotransmisoras a sitios receptores en la membrana de la neurona receptora (la neurona possináptica). La unión de las moléculas de transmisor a los sitios receptores, como una llave en una cerradura, abre las “puertas” (canales iónicos) a iones específicos que pasan a través de la membrana celular llevando consigo su carga eléctrica. Esto da como resultado un cambio en el estado eléctrico de la neurona, un potencial postsináptico (es decir, un cambio de voltaje en la neurona possináptica) —si ocurre un cambio positivo en el voltaje, se llama EPSP, un potencial possináptico excitatorio; si ocurre un cambio negativo en el voltaje en el postsináptico neurona, entonces se le llama IPSP, un potencial inhibitorio postsináptico. Los EPSP son causados por neurotransmisores excitadores (por ejemplo, glutamato; acetilcolina; histamina; epinefrina), IPSP por transmisores inhibidores (por ejemplo GABA, ácido gamma aminobutírico; serotonina, también conocida por su nombre químico, 5-hidroxitriptamina, abreviada 5-HT).
Estos PSP, ya sean excitatorios o inhibidores, se denominan potenciales graduados porque no son de un voltaje fijo, sino que varían en voltaje dependiendo de la cantidad de neurotransmisor (y otros factores) que se ha liberado en los sitios receptores en el Dendríta de neurona postsináptica. Esto contrasta con el potencial de acción (el impulso nervioso) que es de un voltaje fijo y es “todo o ninguno” lo que significa que si ocurre, se produce a toda su fuerza o nada en absoluto. Piensa en un arma de fuego disparando. Si aprietas el gatillo de la pistola con suficiente fuerza (una especie de “umbral de gatillo” de fuerza), entonces el arma dispara una bala con toda su fuerza. Pero si no se alcanza el umbral del gatillo, el arma no dispara en absoluto. No obtienes ni medio disparo ni un octavo de disparo si tiras con menos fuerza, solo puedes obtener un resultado de “todo o ninguno”: los disparos de arma, o no lo hace. O la fuerza sobre el gatillo alcanza el umbral y el arma dispara con toda su “fuerza”, o si la fuerza es insuficiente, el arma no dispara una bala. Lo mismo ocurre con el potencial de acción; si ocurre un potencial de acción, ocurre “todo o ninguno”, a toda su fuerza (voltaje), o nada, y, como el arma, la neurona “dispara” solo si su umbral de disparo o umbral de disparo (en voltaje, alrededor de -55 milivoltios para la mayoría de las neuronas) se alcanza. Los potenciales graduados (EPSP e IPSP) son similares a las señales analógicas (de diferentes voltajes), mientras que los potenciales de acción son similares a las señales digitales (voltaje fijo).
Tipos de neuronas
¡No todas las neuronas son creadas iguales! Hay neuronas que reciben información sobre el mundo que nos rodea, neuronas sensoriales. Hay neuronas motoras que nos permiten iniciar el movimiento y el comportamiento, permitiéndonos finalmente interactuar con el mundo que nos rodea. Finalmente, hay interneuronas, que procesan la entrada sensorial de nuestro entorno en representaciones significativas, planifican la respuesta conductual adecuada y se conectan con las neuronas motoras para ejecutar estos planes de comportamiento.
Figura\(\PageIndex{10}\): Las neuronas sensoriales transportan información hacia el SNC. Las neuronas motoras transportan información del SNC. Las interneuronas transportan información entre neuronas sensoriales y motoras.
Además de esta categorización funcional general de neuronas (sensoriales, motoras e interneuronas), las neuronas también pueden clasificarse por estructura (por ejemplo, unipolar, bipolar, multipolar), forma u otras características. Consulte el Capítulo 4.1 para una visión general de estas clasificaciones. Dado que la forma de la neurona se relaciona con su papel en la comunicación, esta categorización se explora en mayor profundidad a continuación.
Categorizar las neuronas por forma
Las neuronas también se pueden clasificar por su forma. Por ejemplo, Cajal utilizó nombres como cesta, estrellado, musgo, piramidal, etc. para describir diversos tipos de neuronas que observó. Sin embargo, esta clasificación es anatómica y no refleja si la célula es, por ejemplo, una neurona motora o una interneurona. Sin embargo, la forma de las neuronas puede reflejar aspectos de su función y su papel en el procesamiento de la información en el sistema nervioso. Por ejemplo, las neuronas piramidales (Figura\(\PageIndex{11}\)) son “una clase común de neuronas que se encuentra en la corteza cerebral de prácticamente todos los mamíferos, así como en aves, peces y reptiles. Las neuronas piramidales también son comunes en estructuras subcorticales como el hipocampo y la amígdala. Se les llama así por su forma: normalmente tienen un soma (cuerpo celular) que tiene forma de lágrima o pirámide redondeada. También tienden a tener una pulverización cónica de dendritas más largas que emergen del extremo puntiagudo del soma (dendritas apicales) y un racimo de dendritas más cortas que emergen del extremo redondeado. Comprenden alrededor de dos tercios de todas las neuronas de la corteza cerebral de los mamíferos.. son 'neuronas de proyección' — a menudo envían sus axones a largas distancias, a veces fuera del cerebro por completo. Por ejemplo, las neuronas piramidales en la capa 5 de la corteza motora envían sus axones por la médula espinal para impulsar los músculos” (Bekkers, 2011). Son las neuronas excitadoras primarias en la corteza motora y la corteza prefrontal. La tabla\(\PageIndex{1}\) resume las características de las neuronas piramidales.
\ (\ PageIndex {1}\). Características de las neuronas piramidales. (adaptado de John Bekkars; Neuronas Piramindales; Biología Actual).” >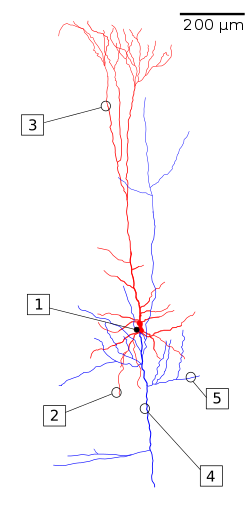 Figura\(\PageIndex{11}\): (izquierda) Reconstrucción de una célula piramidal. Soma y dendritas están marcadas en rojo, axón cenador en azul. (1) Soma, (2) dendrita basal, (3) dendrita apical, (4) axón, (5) axón colateral. (derecha) Neurona piramidal de cultivo orgánico hipocampal de rata. Los axones mostrados (axones) son de otra neurona la mayoría de las cuales no se muestra (Imagen a la izquierda, Wikipedia, La célula piramidal, recuperada 30/8/21. Imagen a la derecha de Wikimedia Commons, Archivo:Neurone pyramidal.jpg; https://commons.wikimedia.org/wiki/F... _pyramidal.jpg; por Mathias De Roo; licenciado bajo la licencia Creative Commons Reconocimiento-Compartir Igual 3.0 Unported.). |
|
| Mesa\(\PageIndex{1}\). Características de las neuronas piramidales. (adaptado de John Bekkars; Neuronas Piramindales; Biología Actual). | |
|---|---|
| \ (\ PageIndex {1}\). Características de las neuronas piramidales. (adaptado de John Bekkars; Neuronas Piramindales; Biología Actual).” scope="fila">Ubicación | Común en la corteza cerebral de mamíferos, especialmente las Capas III y V (la corteza cerebral en mamíferos tiene seis capas celulares); también se encuentra en el hipocampo, y la amígdala. También se encuentran en otros vertebrados |
| \ (\ PageIndex {1}\). Características de las neuronas piramidales. (adaptado de John Bekkars; Neuronas Piramindales; Biología Actual).” scope="fila">forma y números | Piramidal multipolar, pero algunas variaciones entre especies y ubicación en el cerebro; alrededor de 2/3 de todas las neuronas en la corteza cerebral de mamíferos |
| \ (\ PageIndex {1}\). Características de las neuronas piramidales. (adaptado de John Bekkars; Neuronas Piramindales; Biología Actual).” scope="fila">función | Las neuronas de proyección excitatoria con axones largos que transportan potenciales de acción largas distancias, a veces completamente fuera del cerebro; por ejemplo, las neuronas piramidales en la capa 5 de la corteza motora van hasta las neuronas motoras espinales; otras que permanecen en la corteza interconectando áreas distantes de la corteza son críticamente importante en muchas funciones cognitivas |
| \ (\ PageIndex {1}\). Características de las neuronas piramidales. (adaptado de John Bekkars; Neuronas Piramindales; Biología Actual).” scope="fila">Neurotransmisor | El glutamato, el transmisor excitatorio más abundante en el sistema nervioso vertebrado |
Por el contrario, las neuronas estrelladas tienen forma de estrella en virtud de las dendritas que irradian desde el cuerpo celular de la neurona estrellada (ver Figura 5.1.12 (b)). Las neuronas estrelladas espinosas, como las neuronas piramidales, tienen un gran número de espinas dendríticas, pero carecen de la larga dendrita apical característica de las neuronas piramidales. Al igual que las neuronas piramidales, las neuronas estrelladas espinosas en la corteza cerebral son excitadoras y usan glutamato como transmisor. Van Essen y Kelly (1973) reportaron diferentes funciones para las neuronas corticales visuales dependiendo de si eran neuronas piramidales o estrelladas. Después de las neuronas piramidales, las neuronas estrelladas son el segundo tipo más numeroso de neurona cortical. Otro tipo de neurona estrellada que solo tiene espinas escasas es inhibitoria y utiliza GABA (ácido gamma-amino-butírico) como su transmisor.
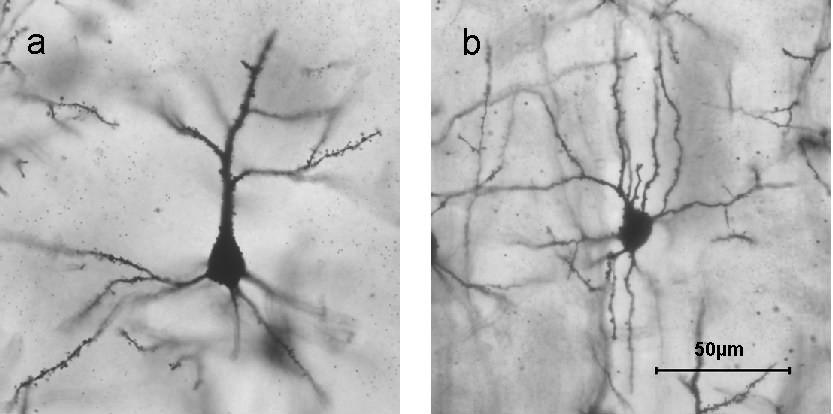
Figura\(\PageIndex{12}\): Neuronas teñidas con Golgi en diferentes capas de corteza cerebral: a) Capa II/III célula piramidal; b) célula estrellada espinosa capa IV. Las espinas dendríticas son visibles en las dendritas ramificadas como racimos de pequeñas cerdas similares a espinas. (Imagen y subtítulo de una oración de Wikipedia, Stellate Cell, recuperado 8/30/21; descripción de espinas dendríticas por Kenneth A. Koenigshofer, Ph.D.).
Brown et al. (2019) encontraron diferentes roles de procesamiento para las neuronas de canasta (ver Figura 5.1.13) y las neuronas estrelladas en el cerebelo. Las células estrelladas y las células de cesta producen sinapsis inhibitorias directamente sobre las neuronas de Purkinje en el cerebelo. Las neuronas estrelladas influyen en el patrón de disparo y las neuronas de canasta afectan la tasa de disparo de las neuronas de Purkinje en el cerebelo. Las células cesta también se encuentran en el neocórtex (corteza cerebral). Las células de cesta son interneuronas inhibidoras. Envuelven cuerpos celulares piramidales en el neocórtex en complejos densos que se asemejan a cestas (Kirkcaldie, 2012). La figura\(\PageIndex{13}\) muestra una variedad de interneuronas clasificadas por forma.
“Aproximadamente el 95% de la actividad neuronal cortical está mediada por neuronas excitadoras rápidas (glutamato, 80%) e inhibidoras rápidas (GABA, 15%). El 5% restante se asocia con la lenta acción moduladora de neuronas monoaminérgicas (dopamina, serotonina, noradrenalina) y no monoaminérgicas (acetilcolina, endorfinas, etc.) localizadas en pequeños núcleos subcorticales del mesencéfalo y proyectándose hacia la corteza cerebral” (Marco Catani, en Encyclopedia of Behavioral Neuroscience, 2a edición, 2022, Science Direct; https://www.sciencedirect.com/topics...pyramidal-cell; recuperado 4/25/22).
Aunque se han clasificado varios tipos básicos de neuronas, el cuadro en el cerebro con respecto a los tipos de neuronas presentes es bastante complejo, como se expresa en esta cita: “Mientras que en la médula espinal podríamos distinguir fácilmente las neuronas en función de su función [sensorial, interneurona, motora], ese no es el caso en el cerebro. Ciertamente, hay neuronas cerebrales involucradas en el procesamiento sensorial —como las de la corteza visual o auditiva— y otras involucradas en el procesamiento motor, como las del cerebelo o la corteza motora. Sin embargo, dentro de cualquiera de estas regiones sensoriales o motoras, hay decenas o incluso cientos de diferentes tipos de neuronas. De hecho, los investigadores todavía están tratando de idear una manera de clasificar pulcramente la enorme variedad de neuronas que existen en el cerebro. parte de lo que le da al cerebro su complejidad es la enorme cantidad de tipos de neuronas especializadas. Los investigadores todavía están tratando de ponerse de acuerdo sobre cuáles son estos” (Universidad de Queensland, n.d.).
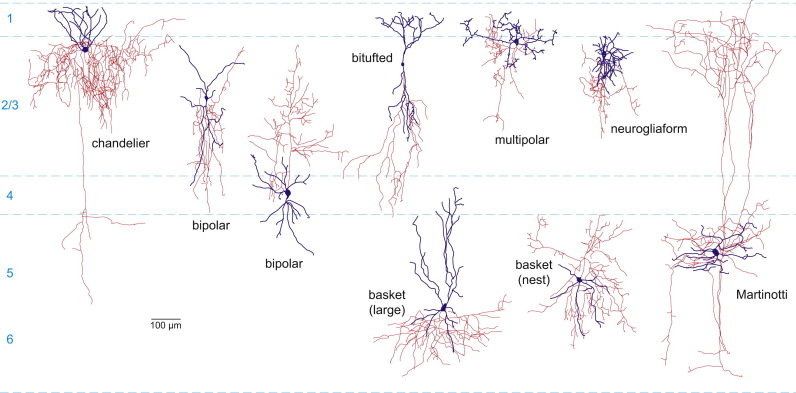
Figura\(\PageIndex{13}\): (arriba). Variedad de tipos de interneuronas clasificadas por forma. Morfologías representativas de tipos interneuronales de ratón. Tenga en cuenta que los cenadores axones (rojos) y dendritas (azules) suelen ser menos elaborados que en la corteza de primates. Se muestran dos variantes de células bipolares y células de cesta (grandes y nido) (Kirkcaldie, 2012).
Otra forma más de clasificar las neuronas es por el neurotransmisor que utilizan (ver Transmisión sináptica a continuación). Quizás igualmente importante es intentar trazar las conexiones que hacen las neuronas con otras neuronas para tratar de entender sus entradas y salidas y qué roles desempeñan en el procesamiento. Esto se ha hecho para muchas células estrelladas y cesteladas, inhibidoras interneuronas sinapsándose en células de Purkinje en el cerebelo. Sin embargo, debido a la complejidad de los circuitos cerebrales esto es muy difícil de lograr, especialmente en la corteza cerebral, aunque es posible en los casos en que las neuronas pertenecen a grandes tractos neuronales o vías con conexiones directas que son más fáciles de rastrear, como las neuronas piramidales en motor córtex a neuronas motoras de la médula espinal o del tálamo a la corteza sensorial. La comprensión de los complejos circuitos del cerebro en su totalidad puede que nunca se logre completamente. Con 80-100 mil millones de neuronas en el cerebro humano y la mayoría haciendo quizás muchos miles de conexiones, ¡el número de conexiones en el cerebro humano es astronómico!
Referencias
Bekkers, J. M. (2011). Neuronas piramidales. Biología actual, 21 (24), R975.
Brown, A. M., Arancillo, M., Lin, T., Catt, D. R., Zhou, J., Lackey, E. P.,... & Sillitoe, R. V. (2019). Las interneuronas de capa molecular dan forma a la actividad de pico de las células cerebelosas de Purkinje. Informes científicos, 9 (1), 1-19.
Churchland, P. M. (2013). Materia y conciencia. Prensa MIT.
Fields, R. D., & Stevens-Graham, B. (2002). Nuevos conocimientos sobre la comunicación neurona-glía. Ciencia, 298 (5593), 556-562.
Kirkcaldie, M. T. (2012). Neociórtex. En El sistema nervioso del ratón (pp. 52-111). Prensa Académica.
Koenigshofer, K.A. (2011). Diseño de la mente: La organización adaptativa de la naturaleza humana, las mentes y el comportamiento. Educación Pearson, Boston.
Mishra A, Singh S, Shukla S. (2018). Bases Fisiológicas y Funcionales de los Receptores de Dopamina y su Papel en la Neurogénesis: Posible Implicación para la Enfermedad de J Exp Neurosci. 12: 1179069518779829. [Artículo gratuito de PMC] [PubMed]
Nadim, F., & Bucher, D. (2014). Neuromodulación de neuronas y sinapsis. Opinión actual en neurobiología, 29, 48-56.
Pasternak, J. F., & Woolsey, T. A. (1975). Sobre la “selectividad” del método Golgi-Cox. J Comp Neurol, 160 (3), 307-312. doi: 10.1002/cne.901600304
Perea, G., Sur, M., & Araque, A. (2014). Redes neurona-glía: engranaje integral de la función cerebral. Fronteras en la neurociencia celular, 8, 378.
Smit, G. J., & Colon, E. J. (1969). Análisis cuantitativo de la corteza cerebral. I. Aelectividad de la técnica de tinción de Golgi-Cox. Brain Res, 13 (3), 485-510.
Universidad de Queensland, Queensland Brain Institute, Tipos de neuronas, n.d. https://qbi.uq.edu.au/brain/brain-an... /tipos-neuronas; Recuperado 31/8/21
Van Essen, D., & Kelly, J. (1973). Correlación de la forma y función celular en la corteza visual del gato. Naturaleza, 241 (5389), 403-405.
Atribuciones
1. El Capítulo 5, Comunicación dentro del Sistema Nervioso, sección 5.1., “Las neuronas y sus funciones básicas”, escrito por Kenneth A. Koenigshofer, Ph.D. (excepto material enumerado en las atribuciones a continuación), está licenciado bajo CC BY 4.0
2. Figura 5.1.2, Vocabulario, Preguntas de Discusión, Recursos Externos, y “Manchas Nissl y Golgi” adaptada por Kenneth A. Koenigshofer de: Furtak, S. (2021). Neuronas. En R. Biswas-Diener & E. Diener (Eds), serie de libros de texto Noba: Psicología. Champaign, IL: Editores DEF. Recuperado a partir de http://noba.to/s678why4
Licencia Creative Commons



 Neuronas de Sharon Furtak está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Los permisos más allá del alcance de esta licencia pueden estar disponibles en nuestro Acuerdo de licencia.
Neuronas de Sharon Furtak está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Los permisos más allá del alcance de esta licencia pueden estar disponibles en nuestro Acuerdo de licencia.
3. Figuras 5.1.4., 5.1.5, y algún texto adaptado de: Biología General (Sin límites), Capítulo 35, El sistema nervioso;
https://bio.libretexts.org/Bookshelv...gy_(Boundless); el contenido de LibreTexts está licenciado por CC BY-NC-SA 3.0. Jurídico.
4. Figuras 5.1.9 y 5.1.10 adaptadas del Capítulo 11.3 (Neuronas y Células Gliales), 11.4 (Impulsos Nerviosos) en Libro: Biología Humana (Wakim & Grewal) - Biología LibreTexts by Suzanne Wakim & Mandeep Grewal, bajo licencia CC BY-NC
Serie de videos: Neurobiología/Biopsicología - Animaciones tutoriales de potenciales de acción, potenciales de membrana en reposo y transmisión sináptica.
- http://www.sumanasinc.com/webcontent/animations/neurobiology.html
- Video: Una animación y una explicación de un potencial de acción
- Video: Una animación de las acciones de los neurotransmisores en la sinapsis
- Video: Una animación interactiva que permite a los estudiantes observar los resultados de manipulaciones a potenciales postsinápticos excitatorios e inhibitorios. También incluye animaciones y explicaciones de transmisión y circuitos neuronales.
- https://apps.childrenshospital.org/clinical/animation/neuron/
- Video: Otra animación de un potencial de acción
- Video: Otra animación de las acciones de los neurotransmisores en la sinapsis
- Video: Potencial de acción dominó: Esta actividad práctica ayuda a los estudiantes a comprender el complejo proceso del potencial de acción, así como a familiarizarse con las características de transmisión (por ejemplo, respuesta de todo o ninguno, período refractario).
- Video: Por perspectiva sobre técnicas en neurociencia para mirar dentro del cerebro
- Video: El cerebro comportándose es el tercer programa de la serie DISCOVERING PSICOLOGY Este programa analiza la estructura y composición del cerebro humano: cómo funcionan las neuronas, cómo se recopila y transmite la información, y cómo las reacciones químicas se relacionan con el pensamiento y el comportamiento.
- http://www.learner.org/series/discoveringpsychology/03/e03expand.html
- Video: Se pueden cultivar nuevas células cerebrales. Aquí\\'s cómo. - ¿Podemos, como adultos, crecer nuevas neuronas? La neurocientífica Sandrine Thuret dice que podemos, y ofrece investigación y consejos prácticos sobre cómo podemos ayudar a nuestros cerebros a realizar mejor la neurogénesis, mejorando el estado de ánimo, aumentando la formación de memoria y previniendo el declive asociado con el envejecimiento en el camino.
- Web: Para más información sobre el Premio Nobel compartido por Ramón y Cajal y Golgi
- http://www.nobelprize.org/nobel_prizes/medicine/laureates/1906/