
5.2: Las neuronas generan cambios de voltaje en la información del código
- Page ID
- 148304
Visión general
Según los materialistas (aquellos que creen que todo en el universo es físico), todas las actividades mentales de nuestra mente y todos nuestros comportamientos son productos de las actividades físicas del cerebro y del sistema nervioso. Estas operaciones de procesamiento de información que crean nuestras mentes y controlan nuestro comportamiento dependen de las neuronas y sus interacciones eléctricas y químicas. Las neuronas producen potenciales eléctricos (voltajes) que actúan como señales en las actividades de procesamiento de información del cerebro. Como se mencionó anteriormente, las neuronas se comunican entre sí a través de espacios sinápticos utilizando sustancias químicas conocidas como neurotransmisores. En este módulo, examinamos cómo las neuronas crean potenciales eléctricos, incluidos los potenciales graduados (potenciales possinápticos) y el impulso nervioso (el potencial de acción), y los procesos de transmisión sináptica. Los potenciales graduados pueden variar en voltaje, mientras que el potencial de acción se fija en voltaje para cualquier neurona en particular y se dice que es todo o ninguno, si ocurre, ocurre a su plena fuerza o no en absoluto. Esto es analágico al disparo de un arma de fuego. Tiene un umbral de disparo igual a una cantidad particular de presión aplicada a su gatillo y luego se alcanza esa presión umbral, el arma dispara y dispara con toda su “fuerza”. De igual manera, el potencial de acción solo se generará si se alcanza su umbral de activación, un voltaje particular dentro de la neurona. Cuando esto sucede, el potencial de acción se genera en la neurona en su “fuerza” completa, medida en voltaje, y este impulso nervioso se conduce a lo largo del axón hasta las terminaciones del axón que luego liberan neurotransmisor en sitios receptores ubicados en la neurona receptora. Ahora examinamos estos procesos con mayor detalle en este módulo y en el siguiente.
¿Cómo producen potenciales eléctricos las neuronas?
Como se describe en el Módulo 5.1, las neuronas, independientemente del tipo, utilizan cambios de voltaje, conocidos como potenciales eléctricos, para codificar y procesar información. Pero, ¿cómo lo hacen? ¿Cómo producen las neuronas el voltaje que luego usan como señales eléctricas en el código electroquímico del cerebro? Más específicamente, la pregunta es, ¿cómo producen las neuronas potenciales eléctricos como el potencial de reposo, el potencial de acción y los potenciales possinápticos?
Las neuronas producen potenciales eléctricos o voltajes por la distribución desigual y el movimiento de átomos cargados eléctricamente llamados iones a través de la membrana celular de la neurona. Estos iones provienen principalmente de sales disueltas en los fluidos corporales dentro y fuera de las neuronas. Los principales iones utilizados por las neuronas para producir sus voltajes son sodio (Na+), potasio (K+) y cloruro (Cl-). Observe que los dos primeros son iones positivos y los últimos, iones cloruro, están cargados negativamente. Un cuarto ion, los aniones orgánicos (A-), que son proteínas grandes (a escala molecular) cargadas negativamente, se fabrican dentro de la neurona. Son demasiado grandes para cruzar la membrana celular y, por lo tanto, dan un sesgo negativo al voltaje de reposo de la neurona. La distribución o concentraciones de estos iones dentro y fuera de una neurona determinan su voltaje (el voltaje es solo la separación física de partículas cargadas, como en una batería de automóvil con sus polos positivo y negativo alrededor de los cuales se concentran partículas cargadas positiva y negativamente flotando en la batería ácido).
Por estos medios, cuatro tipos principales de voltajes neuronales o potenciales neuronales son producidos por las neuronas: 1) el potencial de reposo (a menudo igual a aproximadamente 70 milésimas de voltio negativas, -70 milivoltios) 2) dos tipos de potenciales possinápticos (PSP) —potenciales possinápticos excitatorios (EPSP) e inhibidores post- potenciales sinápticos (IPSP) y 3) el potencial de acción (AP, el impulso nervioso). Examinemos estos potenciales con más detalle y veamos cómo se generan dentro de las neuronas. Estos potenciales eléctricos en grandes poblaciones de neuronas interactuantes codifican la información que guía nuestro comportamiento y produce nuestra experiencia psicológica del mundo.
La Membrana Celular Neurona
La membrana celular, que está compuesta por una bicapa lipídica de moléculas de grasa, separa el fluido dentro de la célula del fluido extracelular circundante. Existen proteínas que atraviesan la membrana, formando canales iónicos que permiten, cuando están abiertos, que iones particulares pasen entre el fluido intracelular y extracelular (ver Figura 8). Estos iones se encuentran en diferentes concentraciones dentro de la célula en relación con el exterior de la célula, y los iones tienen diferentes cargas eléctricas. Debido a esta diferencia en la concentración y carga de iones, en parte forzada por la barrera física de la membrana celular cuando se cierran los canales iónicos a iones específicos, se produce un voltaje, el potencial de reposo.
El potencial de la membrana en reposo
El potencial de reposo se puede considerar como un voltaje basal a partir del cual se generan los otros potenciales neuronales. Es el voltaje dentro de una célula nerviosa cuando está en reposo, es decir, no está recibiendo entradas (PSP) en este momento ni generando ninguna salida (potenciales de acción) en este momento. En este estado de “reposo”, el voltaje dentro de la célula nerviosa es aproximadamente de -70 mv. Es negativo dentro de la neurona en “reposo” porque hay más iones negativos dentro de la neurona (el fluido intracelular) y más iones positivos en el exterior (el fluido extracelular).
Específicamente, hay un gran número de iones Na+ (iones de sodio) fuera de la neurona y muy pocos de estos en el interior, cuando la neurona está “en reposo” (cuando está en potencial de reposo). Y hay más iones cargados negativamente dentro de la célula que en el exterior durante el potencial de reposo. Esta distribución desigual de iones a través de la membrana celular establece el “potencial de reposo” eléctrico, lo que lo hace igual a aproximadamente -70 mv en la neurona típica. Durante el potencial de reposo (potencial de membrana en reposo) se cierran los canales iónicos a Na+.
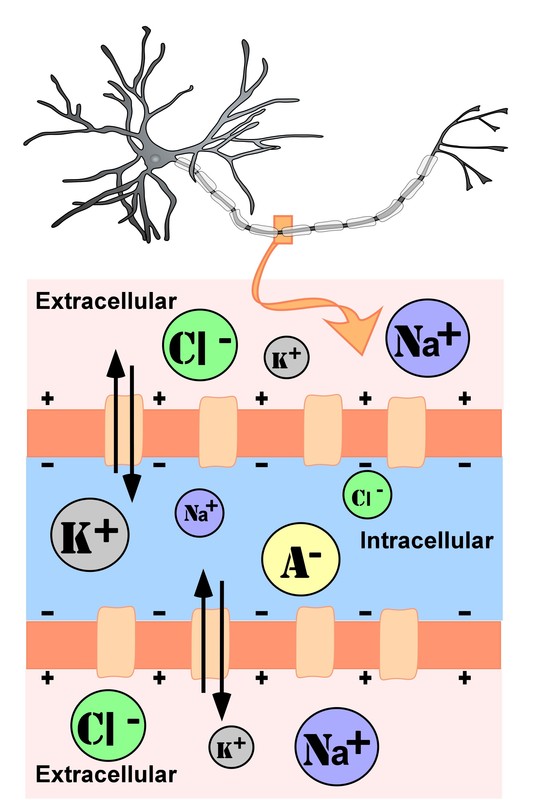
Figura\(\PageIndex{1}\): Representación de concentraciones de iones dentro (intracelulares) y externos (extracelulares) de una neurona en el segmento no mielinizado del axón. El tamaño de los círculos representa las concentraciones relativas de cada ion dentro y fuera de la neurona; tenga en cuenta que cuando la neurona está “en reposo” tiene una carga neta negativa dentro, el potencial de reposo, igual a aproximadamente -70 milivoltios en la mayoría de las neuronas. También tenga en cuenta las mayores concentraciones de iones sodio (Na+) y cloruro (Cl-) fuera de la célula. El sodio en el exterior de la neurona “quisiera” moverse hacia la célula a lo largo de sus gradientes de carga y concentración, pero no puede cuando la célula está en reposo, porque los canales iónicos para los iones de sodio están cerrados, creando una barrera física que impide que la mayoría de los iones de sodio extracelulares crucen a través de la célula membrana. Pero, ¿qué pasa cuando se abren los canales de sodio? Y, ponte a prueba, ¿qué abre los canales iónicos?
La Bomba de Sodio-Potasio
Durante el potencial de reposo, la bomba de sodio-potasio mantiene una diferencia de carga a través de la membrana celular de la neurona. La bomba de sodio-potasio es un mecanismo de transporte activo que mueve los iones de sodio fuera de las células y los iones de potasio hacia las células. La bomba de sodio-potasio mueve ambos iones de áreas de menor a mayor concentración, utilizando energía en ATP y proteínas portadoras en la membrana celular (ver Figura 5.2.2).
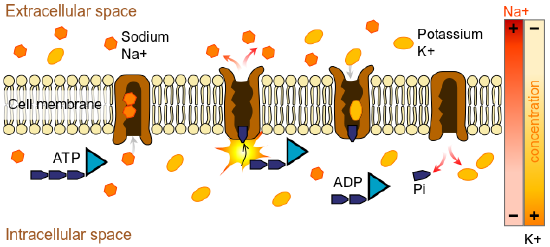
Figura\(\PageIndex{2}\): La bomba de sodio-potasio ayuda a mantener el potencial de reposo de una neurona. Durante el potencial de reposo, hay más carga negativa dentro que fuera de la célula creando un potencial de reposo de -70mv. Durante el potencial de reposo, algunos Na+ se filtran en la neurona y algunos K+ se filtran. El ATP (trifosfato de adenosina) proporciona energía para bombear sodio y potasio hacia la célula. Hay más concentración de sodio fuera de la membrana y más concentración de potasio dentro de la célula debido en parte al movimiento desigual de estos iones por parte de la bomba. La presencia de aniones orgánicos cargados negativamente (A-) contribuye a la carga negativa neta (-70mv) dentro de la neurona en reposo. (Imagen de Wikimedia Commons; Archivo:Esquema sodio-potasio pump-en.svg; https://commons.wikimedia.org/wiki/F...um_pump-en.svg; por LadyOfHats Mariana Ruiz Villarreal; liberado al dominio público por su autor, LadyOfHats . Esto se aplica en todo el mundo).
Producir otros potenciales de membrana: potenciales possinápticos y el potencial de acción
¿Cómo se producen los otros potenciales neuronales? Para generar los otros potenciales neuronales, que son solo cambios de voltaje lejos del potencial de reposo, debe ocurrir una redistribución de iones (y sus cargas eléctricas) a través de la membrana celular. En definitiva, los iones deben moverse.
Hay dos fuerzas principales (llamadas gradientes) que pueden hacer que estos iones se muevan.
Primero, las cargas opuestas se atraen entre sí (“los opuestos se atraen”), y las cargas similares se repelen entre sí. Cuando los iones de carga opuesta se distribuyen de manera desigual a través de la membrana celular (como es el caso durante el potencial de reposo), esto establece lo que se llama un gradiente de carga (una distribución desigual de partículas cargadas) que crea una fuerza electrostática. Si se permite que estos iones se muevan libremente, se moverán a lo largo del gradiente de carga. Las cargas positivas se moverán hacia las negativas y viceversa.
En segundo lugar, cuando los iones de cualquier tipo particular (por ejemplo, iones de sodio) se distribuyen de manera desigual a través de la membrana celular (como más Na+ fuera de la neurona que dentro durante el potencial de reposo), esto establece lo que se llama un gradiente de concentración que puede causar difusión, la red movimiento de iones de una región de mayor concentración a una región de menor concentración. La difusión es impulsada por un gradiente de concentración. Dichos gradientes de concentración para varios iones (iones sodio, potasio y cloruro) existen cuando la neurona está en reposo potencial (ver Figura 10). Los iones, si se les permite moverse libremente, se moverán a lo largo de sus gradientes de concentración, de manera que los iones de un tipo particular (como los iones Na+) se moverán de una región de alta concentración (de Na+) a una región de menor concentración (de Na+).
En resumen, los iones, si se les permite moverse libremente, se moverán a lo largo del gradiente de carga y a lo largo de sus gradientes de concentración. Los iones “quieren” moverse para igualar sus concentraciones a través de la membrana celular y también “quieren” moverse para igualar las cargas a través de la membrana celular (moviéndose hacia cargas opuestas y alejándose de cargas similares).
Sin embargo, es importante señalar que estas dos fuerzas creadas por el gradiente de carga y el gradiente de concentración para cada ion pueden oponerse entre sí en el caso de algunos iones, o pueden actuar en concierto (como es el caso de los iones Na+ durante el potencial de reposo).
Veamos cómo estas dos fuerzas, la difusión (debido a los gradientes de concentración) y la presión electrostática (debido a un gradiente de carga), actúan sobre los cuatro grupos de iones mencionados anteriormente.
- Aniones (A-): Los aniones están altamente concentrados dentro de la célula y contribuyen a la carga negativa del potencial de membrana en reposo. La difusión y la presión electrostática no son fuerzas que determinen A - concentración porque A - es impermeable a la membrana celular. No hay canales iónicos que permitan que A - se mueva entre el fluido intracelular y extracelular.
- Potasio (K+): La membrana celular es muy permeable al potasio en reposo, pero el potasio permanece en altas concentraciones dentro de la célula. La difusión creada por el gradiente de concentración empuja K + hacia el exterior de la célula porque está en alta concentración dentro de la célula. Sin embargo, la presión electrostática creada por el gradiente de carga empuja K + a la celda porque la carga positiva de K + es atraída hacia la carga negativa dentro de la celda. En combinación, estas fuerzas se oponen entre sí con respecto a K + con el gradiente de carga (las cargas opuestas atraen) dominando el gradiente de concentración para que el efecto neto sea empujar y mantener K+ dentro de la neurona en concentraciones más altas.
- Cloruro (Cl-): La membrana celular también es muy permeable al cloruro en reposo, pero el cloruro permanece en alta concentración fuera de la célula. La difusión creada por un gradiente de concentración para Cl- empuja a Cl - hacia el interior de la célula porque se encuentra en alta concentración fuera de la célula. Sin embargo, la presión electrostática creada por el gradiente de carga para Cl- empuja a Cl - hacia el exterior de la celda porque la carga negativa de Cl - es atraída a la carga positiva fuera de la celda creada principalmente por la alta concentración de Na+ allí. Similar a K +, estas fuerzas se oponen entre sí con respecto a Cl -y nuevamente, en este caso, como Na+, el gradiente de carga más potente (como las cargas repelen) supera el gradiente de concentración más débil para Cl-.
- Sodio (Na+): La membrana celular no es muy permeable al sodio cuando la neurona está en reposo. La difusión creada por un gradiente de concentración empuja Na + hacia el interior de la célula porque está en alta concentración fuera de la célula. La presión electrostática creada por el gradiente de carga para Na+ también empuja a Na+ hacia el interior de la celda porque la carga positiva de Na + es atraída a la carga negativa dentro de la celda. Ambas fuerzas empujan a Na + dentro de la célula; sin embargo, como se discutió anteriormente, Na + no puede permear la membrana celular porque los canales para Na+ están cerrados y por lo tanto Na+ permanece en alta concentración fuera de la célula. Las pequeñas cantidades de Na + dentro de la célula son removidas por la bomba de sodio-potasio (Figura 5.2.2), que utiliza la energía de la neurona (trifosfato de adenosina, ATP) para bombear tres iones Na + fuera de la célula a cambio de traer dos iones K + dentro de la célula.
Aunque los iones “quieren” moverse de esta manera dictada por sus gradientes de carga y gradientes de concentración, Na+ no puede moverse libremente cuando la neurona está en reposo potencial de membrana porque los canales iónicos de Na+ están cerrados y la membrana celular actúa como una barrera física a los iones Na+, preservando las distribuciones desiguales de Iones Na+ dentro y fuera de la neurona. Para obtener el movimiento iónico, se deben abrir los poros o canales iónicos en la membrana celular de la neurona “en reposo”, superando la barrera física creada por la membrana celular cuando la neurona está “en reposo” (no recibiendo ninguna entrada a través de sus dendritas y no generando ninguna salida, potenciales de acción, a través de su axón) y su los canales de sodio están cerrados.
¿Qué es lo que hace que los canales iónicos se abran, permitiendo que los iones se muevan a lo largo de sus gradientes? Puedes ponerte a prueba aquí para ver cuánto has retenido de tu lectura hasta el momento. ¿Qué es lo que abre los canales iónicos?
La llegada de moléculas transmisoras de la terminación axónica de una neurona presináptica, y su unión a sitios receptores possinápticos, es el evento clave. Esta interacción de “cerradura y llave” entre las moléculas del transmisor y los sitios receptores possinápticos provoca que se abran “puertas”, canales iónicos específicos.
Cuando los canales iónicos específicos se abren de esta manera (canales de Na+, por ejemplo), esos iones específicos (Na+, en este caso) se mueven a través de la membrana celular a lo largo de sus gradientes de concentración y carga específicos. Esto provocará un cambio de voltaje lejos del potencial de reposo. Este cambio de voltaje es el potencial possináptico (PSP), en este caso es un potencial possináptico excitatorio (EPSP), un desplazamiento positivo en el voltaje lejos del potencial en reposo porque cuando los canales de Na+ en la membrana celular son abiertos por un neurotransmisor, entonces los iones Na+ se moverán a lo largo de su gradiente de carga y gradiente de concentración en la célula, haciendo que el interior de la neurona esté más cargado positivamente.
Potenciales Possinápticos Excitatorios e Inhibitorios
- Potenciales postsinápticos excitatorios (EPSP): una corriente despolarizante que hace que el potencial de membrana se vuelva más positivo y se acerque al umbral de excitación (el “umbral desencadenante” de menos 55 milivoltios) que conducen a la generación de un potencial de acción (ver Figura 5.2.4); causado por transmisor excitatorio.
- Potenciales postsinápticos inhibitorios (IPSP): una corriente hiperpolarizante que hace que el potencial de membrana se vuelva más negativo (ver Figura 5.2.4) y más alejado del umbral de excitación (el “umbral desencadenante” de menos 55 milivoltios), causado por inhibidores transmisor.
Los potenciales postsinápticos ocurren en una neurona possináptica (de ahí el nombre), una neurona receptora. Cuando dicha neurona recibe una entrada de una neurona presináptica, en forma de moléculas transmisoras que se unen a los sitios receptores possinápticos (que son proteínas que están incrustadas en la membrana de la célula postsináptica), los canales iónicos se abren entonces, los iones se mueven a través de la membrana celular, llevando consigo sus cargas eléctricas. Estos eventos cambian el voltaje dentro de la neurona possináptica. Es este cambio de voltaje lejos del potencial de reposo lo que constituye el potencial possináptico (PSP).
Un EPSP (Potencial Possináptico Excitatorio) es un desplazamiento positivo en el voltaje (despolarización) del potencial en reposo, digamos de -70 mv a -60 mv; como se describió anteriormente, se produce un EPSP cuando los canales de Na+ se abren y los iones Na+ fluyen hacia la neurona a lo largo de sus gradientes de concentración y carga, transportando con ellos sus cargas positivas, haciendo que el interior de la neurona possináptica sea más cargado positivamente. Este cambio positivo en el voltaje también se conoce como despolarización de la neurona y es causado por moléculas transmisoras excitadoras que se unen a sitios receptores possinápticos de la neurona receptora.
Un IPSP (Potencial Post-sináptico Inhibitorio) es un cambio negativo en el voltaje (hiperpolarización) desde el potencial de reposo, digamos de -70 mv a -80 mv; un IPSP ocurre cuando los canales Cl- y K+ se abren. Estos iones luego fluyen a lo largo de gradientes de carga y concentración, lo que hace que los iones Cl- cargados negativamente entren y los iones K+ cargados positivamente se muevan fuera de la neurona possináptica, haciéndola más cargada negativamente. Este incremento en la carga neta negativa dentro de la célula es el IPSP (potencial inhibitorio post-sináptico). Los iones Cl- se mueven contra su gradiente de carga (el voltaje negativo del potencial en reposo repele los iones Cl-) porque el gradiente de concentración que los empuja hacia adentro en la neurona es más fuerte que el gradiente de carga opuesto que intenta empujarlos fuera de la neurona (como las cargas repelen). De manera similar, los iones K+ se mueven hacia afuera en oposición al gradiente de carga que los tira hacia adentro, nuevamente, porque el gradiente de concentración que los tira hacia afuera es más fuerte que el gradiente de carga que intenta atraerlos hacia adentro. Estos movimientos son prevenidos durante el potencial de reposo por la barrera física añadida, la membrana celular, pero cuando estos canales iónicos se abren más de lo que están durante el potencial de reposo, los gradientes mueven los iones causando el IPSP. Como se señaló anteriormente, en el caso de Na+ tanto los gradientes de carga como de concentración tiran de Na+ hacia adentro cuando los canales iónicos de Na+ en la membrana celular de la neurona se abren.
Nótese, nuevamente, que tanto en la EPSP como en la IPSP, es la unión de moléculas de transmisor a sitios receptores possinápticos (como llaves que entran en cerraduras de una forma específica) lo que abre las “puertas”, los canales iónicos, permitiendo que los iones se muevan a través de la membrana celular.
Pero aquí hay un tema clave más. ¿Qué es lo que determina qué canales iónicos se abren, y por lo tanto, si se produce un EPSP o un IPSP? La respuesta estaba implícita arriba. La respuesta es: es el tipo de neurotransmisor y el tipo de sitio receptor que recibe las moléculas transmisoras. Existen dos tipos básicos de neurotransmisores, excitadores e inhibidores.
Los transmisores excitadores (como glutamato, acetilcolina—ACH, norepinefrina—NE, dopamina—DA) son aquellos que abren canales de sodio (Na+) en la membrana possináptica, permitiendo que los iones de sodio, transportando su carga positiva, entren en la célula, haciendo una EPSP.
Los transmisores inhibidores (como ácido gamma-amino-butírico-GABA, serotonina-5-HT o dopamina-DA) son aquellos que afectan los canales de cloruro y potasio en la membrana possináptica, permitiendo que el cloruro siga su gradiente de concentración (superando el gradiente de carga opuesto) en la neurona possináptica y permitiendo que el potasio siga su gradiente de concentración (superando su gradiente de carga opuesto) y saliendo. Estos movimientos iónicos hacen que el interior de la neurona sea más negativo, haciendo un IPSP.
Observe que DA está catalogado como excitatorio e inhibitorio. Eso se debe a que algunos receptores DA post-sinápticos son inhibidores, lo que lleva a IPSP cuando se activan por DA, y otros receptores DA son excitadores, produciendo EPSP cuando se unen con DA.
Suma espacial y temporal
Hay un factor adicional en este proceso. Cada neurona se conecta con muchas otras neuronas, a menudo recibiendo múltiples impulsos de ellas. A veces, un solo potencial postsináptico excitatorio (EPSP) es lo suficientemente fuerte como para inducir un potencial de acción en la neurona postsináptica, pero a menudo múltiples entradas presinápticas deben crear EPSP aproximadamente al mismo tiempo para que la neurona postsináptica sea suficientemente despolarizada para disparar un potencial de acción. La suma, ya sea espacial o temporal, es la adición de estos impulsos en el montículo axónico. En conjunto, la suma sináptica y el umbral de excitación actúan como un filtro para que el “ruido” aleatorio en el sistema no se transmita como información importante.
En cualquier momento, cada neurona puede estar recibiendo mensajes mixtos, tanto EPSP como IPSP. Así, las neuronas receptoras (neuronas postsinápticas) pueden recibir múltiples entradas, simultáneamente o en el tiempo o el espacio. Las múltiples entradas pueden sumar, esto se llama suma y hay dos tipos. La suma espacial se refiere a dos o más PSP que llegan a diferentes ubicaciones (es decir, diferentes espacios en la neurona receptora, por lo tanto, suma espacial) en la neurona possináptica (receptora) simultáneamente o lo suficientemente cerca en el tiempo para que sus voltajes se sumen juntos. Por ejemplo, un IPSP de 25 milivoltios negativos (milésimas de voltio) podría sumarse a un EPSP de 50 milivoltios que ocurre al mismo tiempo = 50 milivoltios positivos - 25 milivoltios = 25 milivoltios netos positivos. Eso sería una suma espacial. El otro tipo de suma es la suma temporal cuando los PSP de una sola fuente presináptica, que llegan a la neurona possináptica en “fuego rápido”, se suman. Por ejemplo, tres EPSP de 10 milivoltios cada uno ocurriendo en secuencia rápida se sumarían juntos. La suma espacial y temporal son importantes para determinar si se alcanzará el “umbral de activación” en la neurona receptora, desencadenando un potencial de acción en esa neurona. La suma permite que la actividad de muchas neuronas de entrada se integre en la neurona que recibe las entradas. Si la despolarización de la membrana no alcanza el nivel umbral, no ocurrirá un potencial de acción.
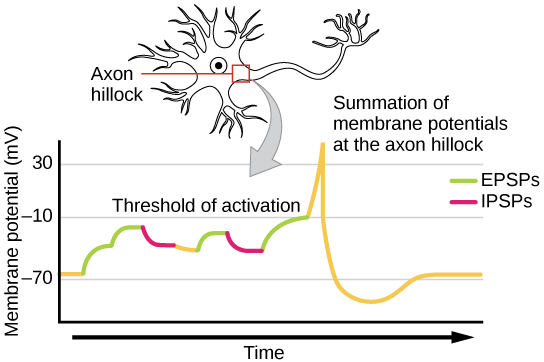
Una neurona a menudo tiene entrada de muchas neuronas presinápticas, ya sean excitadoras o inhibidoras; por lo tanto, los potenciales postsinápticos inhibitorios (IPSP) pueden cancelar los EPSP y viceversa. El cambio neto en el voltaje de la membrana postsináptica determina si la célula postsináptica ha alcanzado su umbral de excitación (“umbral de activación”) necesario para disparar un potencial de acción. Si la neurona solo recibe impulsos excitatorios, también generará un potencial de acción. Sin embargo, si la neurona recibe tanto entradas inhibitorias como excitadoras, la inhibición puede cancelar la excitación y el impulso nervioso se detendrá ahí. Para revisar, la suma espacial significa que los efectos de los impulsos recibidos en diferentes lugares sobre la neurona se suman para que la neurona pueda disparar cuando dichos impulsos se reciben simultáneamente, aunque cada entrada por sí sola no sea suficiente para provocar el disparo. La suma temporal significa que los efectos de los impulsos recibidos en el mismo lugar pueden sumar si los impulsos se reciben en estrecha sucesión temporal. Así, la neurona puede disparar cuando se reciben múltiples entradas, incluso si cada entrada por sí sola no sería suficiente para provocar el disparo.
Puntos Clave
- Los impulsos simultáneos pueden sumarse desde diferentes lugares de la neurona para alcanzar el umbral de excitación durante la suma espacial.
- Cuando los impulsos individuales no pueden alcanzar el umbral de excitación por sí mismos, pueden sumarse en la misma ubicación en la neurona en poco tiempo; esto se conoce como suma temporal.
- El potencial de acción de una neurona se dispara solo cuando el cambio neto de los impulsos excitatorios e inhibidores es distinto de cero.
Términos Clave
- suma temporal: el efecto aditivo cuando se suman impulsos sucesivos (y los PSP resultantes) recibidos en el mismo lugar en la neurona
- suma espacial: el efecto aditivo cuando se suman impulsos simultáneos (y los PSP resultantes) recibidos en diferentes lugares de la neurona
- montículo axón: la parte especializada del soma de una neurona en la raíz del axón donde se suman los impulsos
El Potencial de Acción (el “impulso nervioso”)
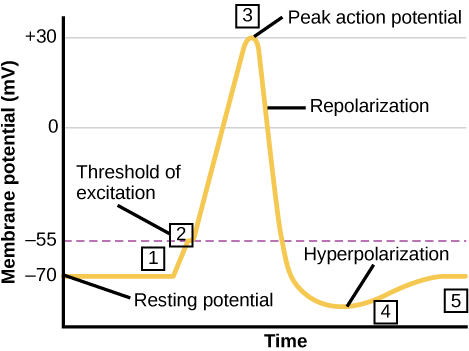
El impulso nervioso, o potencial de acción, se genera en la neurona possináptica solo si se alcanza un “umbral de activación” (“umbral de excitación”) de -55 milivoltios (menos 55 mv) (el umbral de activación varía de neurona a neurona y puede estar en cualquier lugar de menos 65 a menos 55, pero siempre será lo mismo para cualquier neurona en particular. Para fines de nuestra discusión, continuaremos refiriéndonos al umbral de activación de las neuronas como menos 55 milivoltios, no menos 65 como a veces se indica en algunos libros de texto). Cuando se alcanza ese voltaje de activación (por ejemplo, como resultado de un EPSP de al menos 15 milivoltios), entonces todos los canales de iones de sodio regulados por voltaje se abren repentinamente, permitiendo una entrada masiva de iones de sodio, Na+, a la celda a lo largo de gradientes de concentración y eléctricos para el sodio. Esto produce un cambio positivo rápido y grande o “pico” en el voltaje de la neurona possináptica. Este es el impulso nervioso o potencial de acción. En la mayoría de las neuronas, es un desplazamiento positivo de aproximadamente 100 a 130 milivoltios, si medimos desde los -70 milivoltios del potencial de reposo, hasta aproximadamente un positivo de 30 a 60 milivoltios, dependiendo de la neurona. Este valor para cualquier neurona en particular es siempre el mismo para esa neurona. Típicamente, las neuronas con diámetros mayores (tales como fibras A que son típicamente mielinizadas así como de gran diámetro) producen los mayores potenciales de acción (hasta aproximadamente 60 mv positivos, es decir, 130 mv por encima del potencial de reposo), mientras que las neuronas con axones de menor diámetro (clasificadas como fibras C, típicamente no mielinizadas) producir potenciales de acción en el rango inferior.
Tan rápido como aumenta el voltaje del potencial de acción, comienza a caer después de alcanzar su pico (correspondiente a la concentración máxima de Na+ dentro de la neurona). Rápidamente vuelve al potencial de reposo e incluso un poco por debajo del potencial de reposo (el llamado período refractario), ya que los iones Na+ y K+ se mueven, antes del retorno del potencial de la neurona de nuevo a -70 mv, el potencial de reposo. Este rápido ascenso al pico del potencial de acción y luego su rápida caída le da al potencial de acción, cuando se grafica, una apariencia de pico. Por esta razón, los potenciales de acción a menudo son llamados “picos” por los neurocientíficos. Consulte el diagrama del potencial de acción (Figuras 11, 12) para obtener una imagen más clara de estos eventos. Tenga en cuenta que un EPSP, un potencial excitatorio postsináptico, mueve el voltaje de la neurona más cerca del “umbral de activación”, aumentando las posibilidades de que la neurona esté lo suficientemente “excitada” para generar un potencial de acción. Por el contrario, el IPSP, el potencial post-sináptico inhibitorio, aleja el voltaje de la neurona más lejos del “umbral de activación” inhibiendo a la neurona de disparar un potencial de acción.
Una vez que se alcanza el umbral de activación (“umbral de excitación”) (línea punteada en la Figura 9) y se genera un potencial de acción, se conduce a lo largo de la longitud del axón de esta neurona (conducción saltatoria en un axón mielinado; ver arriba). Una vez que la AP alcanza el botón final o terminal del axón de esta neurona, su llegada provoca la liberación de moléculas neurotransmisoras de las vesículas sinápticas localizadas allí. Ahora bien, esta neurona ya no se llama neurona possináptica, sino que se convierte en una neurona presináptica (una neurona emisora) con respecto a la siguiente célula en línea. Es la liberación del transmisor (paso 3 en la lista de 8 pasos en transmisión sináptica que se muestra en Transmisión sináptica a continuación) conduce a los pasos (4 y 5 en la lista de 8 pasos) y se genera un EPSP o IPSP en la siguiente celda en línea. Recuerde que si se produce una EPSP o una IPSP en la siguiente célula en línea se determina por si se ha liberado un transmisor excitatorio o inhibidor de la neurona presináptica (ver los 8 pasos a continuación).
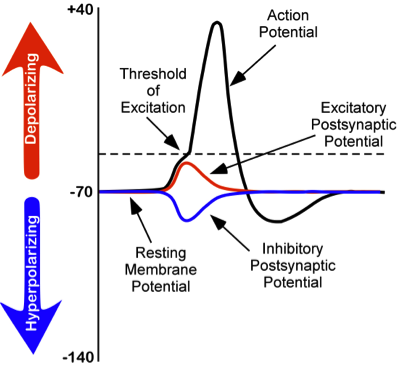
Figura\(\PageIndex{4}\): Cambios en los potenciales de membrana de las neuronas. (izquierda) La línea punteada representa el umbral de activación (“umbral de excitación”), aproximadamente -55 milivoltios, se genera un potencial de acción una vez que se alcanza el voltaje umbral de activación.
Tenga en cuenta que una EPSP (despolarización) mueve el voltaje de la neurona más positivo y, por lo tanto, más cerca del umbral de disparo, haciendo que sea más probable que el voltaje alcance el umbral de disparo, “disparando” un potencial de acción por el axón de la neurona; por lo tanto, es excitador. Un IPSP hace lo contrario. Mueve el voltaje en la dirección negativa, más lejos del umbral de disparo, e inhibe así que la neurona produzca una salida (un potencial de acción) en su axón. También tenga en cuenta que el pico del potencial de acción (parte superior de la línea negra con forma de pico) es el pico de su voltaje positivo y corresponde a la concentración máxima de iones Na+ dentro de la célula como resultado de la apertura de los canales iónicos Na+ después de que se haya alcanzado el umbral de activación. Después de esta concentración pico de Na+ haciendo el pico del voltaje del potencial de acción, los iones cargados positivamente (incluidos los iones K+) comienzan a salir del interior de la celda y a medida que lo hacen, el voltaje positivo dentro de la neurona disminuye progresivamente, correspondiente a la pendiente descendente del pico. Obsérvese además que el pico va más negativo que el potencial de reposo. En este estado la neurona se ve inhibida por este periodo refractario y no puede disparar otro potencial de acción por un breve tiempo. Esto mantiene el potencial de acción como un evento digital discreto. Esto es importante para la codificación fuente, una característica importante de la codificación neuronal, que se discute en el capítulo sobre aprendizaje y memoria en este texto. Todo el proceso desde el desencadenamiento del potencial de acción (que comienza en la raíz del axón, llamado el montículo del axón, más cercano al cuerpo celular) hasta el final del período refractario toma aproximadamente 1 milisegundo, haciendo que la velocidad máxima a la que una neurona pueda generar y “disparar” potenciales de acción ( “impulsos” nerviosos) alrededor de 1,000 por segundo, aunque la mayoría de las neuronas cuando están activas disparan a una frecuencia mucho menor. La frecuencia de los potenciales de acción es un código que el sistema nervioso utiliza para representar la información. Por ejemplo, cuanto más brillante es una fuente de luz, mayor es la frecuencia de potenciales de acción en el nervio óptico en respuesta a ella.
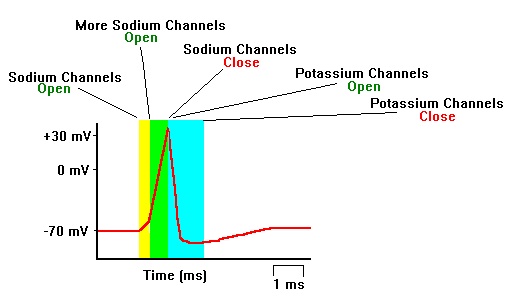
Figura\(\PageIndex{5}\): Formación de un potencial de acción. (izquierda) La formación de un potencial de acción se puede dividir en cinco pasos. (1) Un estímulo de una célula sensorial u otra neurona hace que la célula diana se despolarice hacia el potencial umbral. (2) Si se alcanza el umbral de excitación, todos los canales de Na+ se abren y la membrana se despolariza. (3) En el pico potencial de acción, canales K+ abiertos y K+ comienza a salir de la celda. Al mismo tiempo, los canales de Na+ se cierran. (4) La membrana se hiperpolariza a medida que los iones K+ continúan saliendo de la célula. La membrana hiperpolarizada se encuentra en un periodo refractario y no puede disparar. (5) Los canales de K+ se cierran y el transportador Na+/K+ (que requiere gasto de energía) restaura el potencial de reposo. (derecha) Secuencia de apertura y cierre de los canales iónicos Sodio (Na+) y Potasio (K+) produciendo las fases ascendente y descendente del potencial de acción. (Imagen a la izquierda y pie de foto de Lumen Boundless Biology; Cómo se comunican las neuronas; https://courses.lumenlearning.com/bo...s-communicate/. A menos que se indique lo contrario, el contenido está bajo la licencia Creative Commons Attribution 4.0. Imagen a la derecha de Wikimedia Commons; Archivo.; https://commons.wikimedia.org/wiki/F...8%A8%DB%8C.jpg; por Vidakarimnia; licenciado bajo la licencia Creative Commons Attribution-Share Alike 3.0 Unported).
Una sola neurona por sí misma no puede generar estados psicológicos como sentimientos, percepciones o pensamientos. Las neuronas deben interactuar con otras neuronas. La generación de un pensamiento, u otra experiencia psicológico/mental compleja, requiere que un enorme número de neuronas interactúen entre sí. Para interactuar, deben comunicarse con otras neuronas.
Como se describe en el módulo 5.1, las neuronas se comunican entre sí, a través de espacios entre ellas conocidas como “brechas sinápticas” por la liberación de sustancias químicas (llamadas neurotransmisores o simplemente transmisores) desde el axón de una neurona que termina en sitios receptores en las dendritas o soma de la neurona diana (un neurona). Los neurotransmisores involucrados en esta comunicación entre neuronas se fabrican en el soma de la neurona y luego se transportan por un axón largo donde se almacenan en las vesículas sinápticas hasta que se liberan del axón terminando en el espacio sináptico; la liberación se desencadena por la llegada de una acción potencial (el impulso nervioso) en la terminación del axón.
Como se discute en el módulo 5.1, la sinapsis incluye la neurona presináptica (la célula emisora) y su axón terminando con sus vesículas sinápticas, junto con el hueco sináptico, y la neurona possináptica (la célula receptora) con sus sitios receptores. Hay una enorme cantidad de neuronas en el cerebro humano, pero el número de posibles combinaciones diferentes de conexiones sinápticas entre esos 80-100 mil millones de neuronas es inimaginable; un neurocientífico estimó que el número de posibles patrones de interconexión entre neuronas en un cerebro humano supera el número de átomos en todo el universo! Esta complejidad neuronal parece ser suficiente para codificar toda la información contenida en un cerebro humano incluyendo todos los pensamientos, sentimientos, percepciones y recuerdos de una vida humana.
Puntos Clave
- El potencial de reposo, típicamente igual a -70 milivoltios, es el voltaje dentro de una neurona cuando no recibe entradas de otras neuronas y no produce salidas (sin potenciales de acción)
- Los potenciales possinápticos (PSP), EPSP (excitatorios; despolarizaciones) e IPSP (inhibitorios; hiperpolarizaciones) son cambios de voltaje positivos y negativos, respectivamente, lejos del potencial de reposo de la neurona. Estos cambios de voltaje, PSP, en la neurona possináptica son el resultado de la liberación del transmisor de una o más neuronas presinápticas.
- Los potenciales de acción se forman cuando las entradas (EPSP e IPSP sumadas) hacen que la membrana celular se despolarice más allá del umbral de excitación (“umbral de activación”), provocando que se abran todos los canales iónicos de sodio, lo que lleva a un gran cambio positivo en el voltaje de la neurona (Figuras 5.2.4 y 5.2.5).
- Cuando se abren los canales de iones potasio y se cierran los canales iónicos de sodio, la membrana celular se hiperpolariza a medida que los iones potasio salen de la célula; la célula no puede disparar durante este periodo refractario (Figuras 5.2.4 y 5.2.5).
- El potencial de acción se desplaza por el axón a medida que la membrana del axón se despolariza y repolariza (ver 5.1, Figuras 5.1.4 y 5.1.5).
- La mielina aísla muchos axones para evitar fugas de la corriente a medida que “salta” de nodo a nodo por el axón.
- Los nodos de Ranvier son huecos en la mielina a lo largo de los axones; contienen canales iónicos de sodio y potasio, lo que permite que el potencial de acción viaje rápidamente por el axón saltando de un nodo al siguiente (conducción saltatoria; ver módulo 5.1).
Términos Clave
- potencial de acción: un cambio a corto plazo en el potencial eléctrico que viaja a lo largo de una celda
- despolarización: una disminución en la diferencia de voltaje entre el interior y el exterior de la neurona
- hiperpolarizar: aumentar la polaridad de algo, especialmente la polaridad a través de una membrana biológica
- ganglio de Ranvier: una pequeña constricción en la vaina de mielina de los axones
- conducción saltatoria: el proceso de regeneración del potencial de acción en cada nodo de Ranvier
Atribuciones
1. Capítulo 5, Comunicación dentro del Sistema Nervioso, 5.2. “Las neuronas generan cambios de voltaje a la información del código” por Kenneth A. Koenigshofer, PhD, Chaffey College, está licenciado bajo CC BY 4.0
2. Figuras 5.2.1, 5.2.4, Vocabulario, Preguntas de Discusión, Recursos Externos, y algún texto adaptado de: Furtak, S. (2021). Neuronas. En R. Biswas-Diener & E. Diener (Eds), serie de libros de texto Noba: Psicología. Champaign, IL: Editores DEF. Recuperado de http://noba.to/s678why4; Neuronas por Sharon Furtak en NOBA está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Figura 5.2.4 pie de foto de Kenneth A. Koenigshofer, PhD, Chaffey College.
3. “Puntos Clave” y “Términos Clave” adaptados de: Biología General (Sin Límites), Capítulo 35, El Sistema Nervioso;
https://bio.libretexts.org/Bookshelv...gy_(Boundless); el contenido de LibreTexts está licenciado por CC BY-NC-SA 3.0. Jurídico.
4. Figuras 5.2.3, 5.2.5, y algún texto adaptado de: Biología general (sin límites), Capítulo 35, El sistema nervioso;
https://bio.libretexts.org/Bookshelv...gy_(Boundless); el contenido de LibreTexts está licenciado por CC BY-NC-SA 3.0. Jurídico.
5. Figuras 5.2.2 adaptadas del Capítulo 11.3 (Neuronas y Células Gliales), 11.4 (Impulsos Nerviosos) en Libro: Biología Humana (Wakim & Grewal) - Biología LibreTexts by Suzanne Wakim & Mandeep Grewal, bajo licencia CC BY-NC
Licencia Creative Commons



 Neuronas de Sharon Furtak está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Los permisos más allá del alcance de esta licencia pueden estar disponibles en nuestro Acuerdo de licencia.
Neuronas de Sharon Furtak está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Los permisos más allá del alcance de esta licencia pueden estar disponibles en nuestro Acuerdo de licencia.