
5.3: Transmisión sináptica
- Page ID
- 148306
Objetivos de aprendizaje
- Describir los canales iónicos y qué cambios experimentan cuando se producen potenciales neuronales; ¿qué hace que los canales iónicos cambien durante la transmisión sináptica?
- Definir receptores ionotrópicos y metabotrópicos y discutir en qué formas difieren entre sí en sus efectos durante la transmisión sináptica
- Explicar los pasos en la transmisión sináptica de neurona presináptica a neurona possináptica
- Describir cómo los transmisores excitatorios e inhibidores difieren en sus efectos sobre las neuronas possinápticas durante la transmisión sináptica
- Discutir cómo las capas de neuronas, simuladas por unidades de procesamiento en redes neuronales artificiales, podrían producir procesos psicológicos como el aprendizaje
Visión general
Después de que se genera un potencial de acción en la neurona presináptica, este impulso total o ninguno se realiza a lo largo del axón hasta la terminación del axón (el botón terminal). En el botón terminal presináptico, la llegada del potencial de acción desencadena la liberación de neurotransmisores (ver Figura 5.3.2). Los neurotransmisores cruzan la brecha sináptica y los subtipos de receptores abiertos en forma de llave y llave (ver Figura 5.3.2). Dependiendo del tipo de neurotransmisor, se produce un EPSP o IPSP en la dendrita de la célula postsináptica. Los neurotransmisores que abren canales de Na+ o calcio (Ca+) provocan una EPSP; un ejemplo son los receptores NMDA, los cuales son activados por el glutamato (el principal neurotransmisor excitatorio en el cerebro). En contraste, los neurotransmisores que abren canales Cl- o K+ provocan un IPSP; un ejemplo son los receptores del ácido gamma-aminobutrírico (GABA), los cuales son activados por GABA, el principal neurotransmisor inhibidor en el cerebro. Una vez que las EPSP y las IPSP ocurren en el sitio postsináptico, el proceso de comunicación dentro y entre neuronas se activa en ciclos (ver Figura 5.3.3). Un neurotransmisor que no se une a los receptores es descompuesto e inactivado por enzimas o células gliales, o bien es devuelto al botón terminal presináptico en un proceso llamado recaptación (Figura 5.3.2, Paso 6).
TRANSMISIÓN SINÁPTICA
La transmisión de mensajes neuronales a través de la brecha sináptica, a través de la liberación del transmisor de una neurona presináptica a sitios receptores possinápticos en una neurona possináptica, se llama transmisión sináptica o neurotransmisión. La transmisión sináptica es fundamental para la capacidad del cerebro para procesar información, generar estados mentales y generar comportamiento adaptativo. Algunas sinapsis son puramente eléctricas y hacen conexiones eléctricas directas entre neuronas (por ejemplo, entre células de cesta en el cerebelo). Sin embargo, la mayoría de las sinapsis son sinapsis químicas, involucrando neurotransmisores. La transmisión de señales neuronales a través de sinapsis químicas es más compleja que en las sinapsis eléctricas e implica muchos pasos.
Sinapsis Eléctricas
Las sinapsis eléctricas son mucho menos comunes que las sinapsis químicas, pero sin embargo se distribuyen por todo el cerebro. En las sinapsis químicas se necesita neurotransmisor para la comunicación entre neuronas, pero para las sinapsis eléctricas este no es el caso. En las sinapsis eléctricas, la corriente, los iones y las moléculas pueden fluir entre dos neuronas a través de conexiones físicas directas que permiten que el citoplasma fluya entre ellas. La conexión física entre neuronas con sinapsis eléctricas está en forma de estructuras de poros grandes, llamadas conexones, en uniones de brecha entre tales neuronas. La comunicación entre neuronas con sinapsis eléctricas es más rápida que en las sinapsis químicas que deben pasar por más pasos para transmitir señales a otra neurona. Por lo tanto, las sinapsis eléctricas a menudo se encuentran en sistemas neuronales que requieren una respuesta rápida como reflejos defensivos. Las sinapsis eléctricas pueden comunicar señales en ambas direcciones entre las neuronas en contraste con las sinapsis químicas que transmiten mensajes en una dirección, desde la liberación presináptica por transmisor hasta la neurona possináptica.
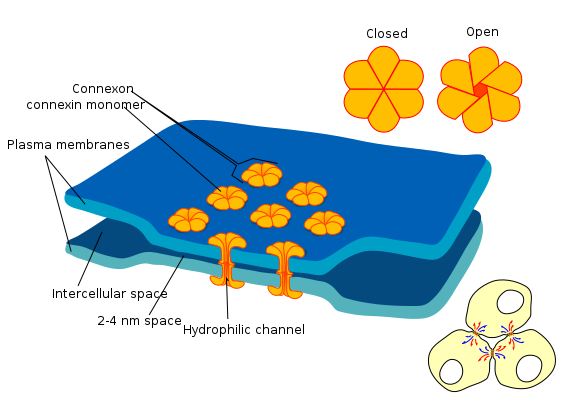
Figura\(\PageIndex{1}\): Gap junction en una sinapsis eléctrica. Dos neuronas adyacentes con sinapsis eléctrica entre ellas pueden comunicarse a través de canales hidrofílicos Tenga en cuenta que la brecha entre las membranas celulares de las neuronas pre y possinápticas en las sinapsis eléctricas es mucho menor que la brecha sináptica en las sinapsis químicas que es aproximadamente 10 veces mayor. (Imagen de Wikimedia Commons; Archivo:Gap cell junction-en.svg; por Mariana Ruiz LadyOfHats. https://commons.wikimedia.org/wiki/F...unction-en.svg; por Mariana Ruiz LadyOfHats; dominio público por su autor, LadyOfHats).
Sinapsis Químicas
Se discutió previamente qué iones están involucrados en el mantenimiento del potencial de la membrana en reposo. No es sorprendente que algunos de estos mismos iones estén involucrados en el potencial de acción. Cuando la celda se despolariza (carga más positiva) y alcanza el umbral de excitación, esto provoca que se abra un canal de Na+ dependiente del voltaje. Un canal iónico dependiente del voltaje es un canal que se abre, permitiendo que algunos iones entren o salgan de la celda, dependiendo de cuándo la celda alcance un potencial de membrana particular (es decir, un voltaje particular).
Cuando la celda está en el potencial de membrana en reposo, estos canales de Na+ dependientes del voltaje están cerrados. Como aprendimos anteriormente, tanto la difusión de los gradientes de concentración como la presión electrostática de los gradientes de carga están empujando iones Na+ hacia el interior de la célula. Sin embargo, Na+ no puede permear la membrana cuando la célula está en reposo.
Una vez que estos canales se abren cuando se ha alcanzado el umbral de activación, Na+ se precipita dentro de la celda, provocando que la celda se cargue muy positivamente en relación con el exterior de la celda. Esto es responsable de la fase ascendente o despolarizante del potencial de acción (ver Módulo 5.2). El interior de la célula se carga muy positivamente, desde aproximadamente +30mV hasta aproximadamente +60mv, dependiendo de la neurona particular. En este punto, los canales de Na+ se cierran y se vuelven refractarios. Esto significa que los canales de Na+ no pueden volver a abrirse hasta después de que la célula regrese al potencial de la membrana en reposo. Así, un nuevo potencial de acción no puede ocurrir durante el periodo refractario. El periodo refractario también asegura que el potencial de acción solo pueda moverse en una dirección hacia abajo del axón, alejándose del soma.
A medida que la celda se vuelve más despolarizada, se abre un segundo tipo de canal dependiente del voltaje; este canal es permeable a K+. Con el interior de la celda muy positivo en relación con el exterior de la celda (despolarizado) y la alta concentración de K+ dentro de la celda, tanto la fuerza de difusión, a lo largo de gradientes de concentración, como la fuerza de presión electrostática, a lo largo de gradientes de carga, impulsan K+ fuera de la celda. El movimiento de K+ fuera de la célula hace que el potencial celular regrese al potencial de la membrana en reposo; esta es la fase descendente o hiperpolarizante del potencial de acción (ver Módulo 5.2).
Una hiperpolarización corta se produce parcialmente debido al cierre gradual de los canales de K+. Con los canales de Na+ cerrados, la presión electrostática de un gradiente de carga de K+ continúa empujando a K+ fuera de la celda. Además, la bomba de sodio-potasio está empujando a Na+ fuera de la celda. La célula regresa al potencial de la membrana en reposo y el exceso de K+ extracelular se difunde. Este intercambio de iones Na+ y K+ ocurre muy rápidamente, en menos de un milisegundo. El potencial de acción se produce en un movimiento ondulatorio hacia abajo del axón hasta que alcanza el botón terminal. Solo se ven afectados los canales iónicos muy próximos al potencial de acción, lo que provoca que el potencial de acción se regenere a lo largo del axón, lo que crea el movimiento del potencial de acción hacia abajo del axón.
La unión de un neurotransmisor a su receptor es reversible, y por una buena razón. Mientras esté unido a un receptor post sináptico, un neurotransmisor continúa afectando el potencial de membrana, por lo que debe ser eliminado de la sinapsis. Los efectos del neurotransmisor generalmente duran unos milisegundos antes de ser terminados. Si el transmisor utilizado no es removido o inactivado puede provocar sobreactivación de neuronas, potencialmente conduciendo a estados mentales patológicos y comportamiento si se ven afectadas suficientes sinapsis. La terminación del neurotransmisor puede ocurrir de tres maneras: Primero, la recaptación por astrocitos o por el terminal presináptico donde el transmisor usado puede ser destruido por enzimas (Figura 5.3.2, paso 6). En segundo lugar, la degradación por enzimas en la hendidura sináptica como la enzima, la acetilcolinesterasa, que destruye el transmisor de acetilcolina usado. Tercero, difusión del neurotransmisor a medida que se aleja de la sinapsis. Nuevamente, la destrucción o eliminación del transmisor usado de la sinapsis después de que haya realizado su trabajo es esencial para el funcionamiento normal del sistema nervioso.
Sinapsis ionotrópicas y metabotrópicas
Hay dos categorías principales de receptores postsinápticos: receptores ionotrópicos y receptores metabotrópicos.
Los receptores ionotrópicos, cuando son activados por un transmisor de una neurona presináptica, hacen que los canales iónicos se abran, permitiendo que los iones, con sus cargas eléctricas, se muevan a través de la membrana celular de la neurona receptora (postsináptica), provocando una EPSP o una IPSP (ver arriba). Estos son receptores de acción rápida y están involucrados en el tipo de transmisión neural que hemos estado describiendo anteriormente. Los receptores ionotrópicos son receptores en canales iónicos que se abren, permitiendo que algunos iones entren o salgan de la célula, dependiendo de la presencia de un neurotransmisor particular. El tipo de neurotransmisor y la permeabilidad del canal iónico que activa determinará si ocurre un EPSP o IPSP en la dendrita de la célula possináptica. Estos EPSP e IPSP suman y determinan si se generará un potencial de acción. Para un resumen de video, copie y pegue esta dirección web en su navegador: https://nobaproject.com/modules/neurons.
Por el contrario, los receptores metabotrópicos (generalmente acoplados con proteínas G; es decir, proteínas de unión a nucleótidos de guanina), cuando se activan por un transmisor de una neurona presináptica, actúan indirectamente y más lentamente, utilizando segundos mensajeros para producir una variedad de efectos metabólicos para modular la actividad celular. Estos efectos incluyen cambios en la transcripción génica, regulación de proteínas en la célula, liberación de Ca+ (iones calcio) dentro de la célula y efectos sobre los canales iónicos en la membrana celular de la neurona (Sterling & Laughlin, 2015). Dicha modulación de neuronas y sinapsis puede ser más duradera que los efectos de la activación de receptores ionotrópicos y puede desempeñar un papel importante en los mecanismos de aprendizaje y memoria a nivel celular (Nadim y Bucher, 2014).
Figura\(\PageIndex{3}\): Comparación de Receptores Postsinápticos Ionotrópicos y Metabotrópicos. La imagen en la parte superior (a) muestra receptores ionotrópicos que cuando se activan por el transmisor abren canales iónicos inmediatamente dando como resultado movimiento iónico y una respuesta inmediata, un potencial postsináptico. La imagen en la parte inferior (b) muestra receptores metabotrópicos que cuando son activados por el transmisor inician un segundo sistema mensajero. Los segundos mensajeros pueden tener una variedad de efectos que incluyen la apertura indirecta de canales iónicos (Imagen de Wikimedia Commons; Archivo:1226 Receptor Types.jpg; https://commons.wikimedia.org/wiki/F...ptor_Types.jpg; por OpenStax; licenciado bajo la Creative Commons Atribución 4.0 Licencia internacional).
Resumen de 8 pasos de la transmisión sináptica
Aquí hay un resumen de 8 pasos algo simplificado pero útil de los pasos en la transmisión sináptica en sinapsis con receptores ionotrópicos:
2) el aumento de Ca+2 intracelular en la terminación del axón se une a las vesículas sinápticas a la membrana presináptica desencadenando la liberación del transmisor de vesículas sinápticas en la terminación axónica de esta neurona presináptica
3) moléculas de neurotransmisor cruzan el espacio sináptico lleno de líquido entre las neuronas pre y possinápticas
4) moléculas de transmisor se unen a sitios receptores ionotrópicos possinápticos en la dendrita o soma de la neurona possináptica en el otro lado del espacio sináptico
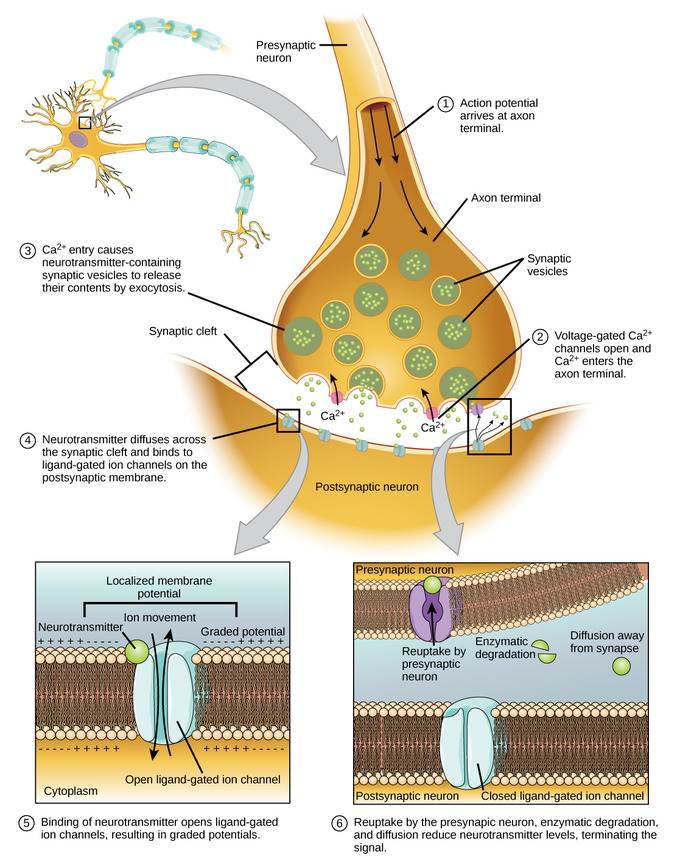
Figura\(\PageIndex{4}\): La imagen de la izquierda muestra dos neuronas comunicándose entre sí (transmisión sináptica). La neurona en la parte superior es la neurona emisora o presináptica. La neurona en la parte inferior es la neurona receptora o possináptica. En la imagen de la izquierda de la figura, observe la pequeña caja en la sinapsis. Esta caja se agranda en la imagen del lado derecho de la figura para revelar detalles de la sinapsis y eventos allí durante la transmisión sináptica. El extremo axón o boton/botón de la neurona presináptica está en la parte superior. Vemos moléculas transmisoras (pequeños cuadrados rojos) transportadas por el axón (flecha) desde el cuerpo celular hasta el axón terminando donde se almacenan en vesículas sinápticas (círculos amarillos que contienen pequeños cuadrados rojos, es decir, moléculas transmisoras). Cuando un impulso nervioso (potencial de acción) alcanza la terminación del axón, los iones de calcio ingresan al extremo axón/botono/botón haciendo que las vesículas sinápticas se muevan hacia la membrana celular del extremo axón (pasos 1 y 2 mencionados anteriormente) causando la liberación de moléculas transmisoras dentro y a través del espacio sináptico (pasos 2 y 3). Las moléculas transmisoras se unen a receptores possinápticos (verdes) provocando que canales iónicos específicos (“puertas”) se abran a iones específicos (pasos 4 y 5). Los iones (no mostrados en esta figura) se mueven a través de los canales iónicos en la membrana celular de la neurona possináptica (etapa 6) con sus cargas eléctricas cambiando el voltaje dentro de la neurona possináptica (ya sea una EPSP o una IPSP). Si se alcanza el umbral de activación (aproximadamente 55 milivoltios negativos), se genera un potencial de acción en la neurona receptora, que ahora se convierte en una neurona emisora para la siguiente célula en línea (no se muestra en esta figura). “La inactivación del transmisor utilizado es la etapa final, etapa 8, y se representa en esta figura por “" degradación enzimática "” (destrucción enzimática).” (Imagen de Wikimedia Commons; Archivo:Neurotransmisor Genérico System.jpg; https://commons.wikimedia.org/wiki/F...ter_System.jpg; por NIDA (NIH); esta obra es de dominio público en Estados Unidos. Leyenda de Kenneth A. Koenigshofer, Ph.D.).
5) esta interacción de “cerradura y llave”, entre las moléculas transmisoras y los sitios receptores a los que se unen, “abre puertas”, es decir, provoca que orificios o poros (canales iónicos) en la membrana celular de la neurona possináptica se abran a iones específicos (átomos cargados eléctricamente; sodio, cloruro, potasio)
6) los iones se mueven (a lo largo de gradientes de carga y concentración) a través de canales iónicos específicos a través de la membrana celular dentro o fuera de la neurona possináptica, llevando consigo sus cargas eléctricas, alterando el potencial eléctrico dentro de la neurona possináptica (como se señaló anteriormente, esto cambio de voltaje se llama potencial possináptico o PSP). Si el transmisor es excitatorio, entonces los canales de sodio se abren y los iones de sodio (Na+) siguen los gradientes eléctricos y de carga hacia la neurona possináptica haciendo que el voltaje dentro de la neurona sea más positivo (un EPSP). Si el transmisor es inhibitorio, piense en los iones cloruro (Cl -) entrando y los iones de potasio (K+) saliendo, haciendo que el interior de la neurona possináptica esté más cargado negativamente, alejando el voltaje de la neurona más lejos del umbral de activación, inhibiendo así el neurona de disparo (esto es un IPSP). Los EPSP y los IPSP pueden sumar.
7) si el cambio de voltaje en la neurona possináptica es positivo (un EPSP) y si es lo suficientemente grande, o si las entradas sumadas (suma) son lo suficientemente positivas como para alcanzar el “umbral de activación” de la neurona possináptica (-55 milivoltios), entonces los canales de Na+ se abren repentinamente (voltaje cerrado canales), Na+ se precipita dentro de la célula, y se genera un potencial de acción en la neurona possináptica (receptora), que ahora se convierte en una neurona presináptica (emisora) para la siguiente célula en línea. El potencial de acción se genera en el montículo del axón y luego se conduce a lo largo del axón, saltando de nodo a nodo si el axón está mielinizado. A medida que el Na+ se precipita dentro de la célula, esto es responsable de la fase ascendente o despolarizante del potencial de acción (ver Figura 5.3.2).
8) Inactivación del transmisor usado (por su recaptación en la neurona presináptica o por su destrucción enzimática por enzimas específicas)
Figura\(\PageIndex{5}\): Comunicación en una sinapsis química: La comunicación en las sinapsis químicas requiere liberación de neurotransmisores (consultar el resumen de 8 pasos anterior). Cuando la membrana presináptica se despolariza, los canales de Ca2+ regulados por voltaje se abren y permiten que el Ca2+ entre en la celda. La entrada de calcio hace que las vesículas sinápticas se fusionen con la membrana y liberen moléculas neurotransmisoras en la hendidura sináptica. El neurotransmisor se difunde a través de la hendidura sináptica y se une a canales iónicos regulados por ligando (canales abiertos por transmisor químico) en la membrana possináptica, dando como resultado una despolarización localizada (EPSP) o hiperpolarización (IPSP) de la neurona possináptica dependiendo del tipo de transmisor. excitatorio o inhibitorio. (Imagen y subtítulo adaptado de: Biología General (Boundless), Capítulo 35, El sistema nervioso; https://bio.libretexts.org/Bookshelv...gy_(Boundless); el contenido de LibreTexts está licenciado por CC BY-NC-SA 3.0. Legal).
Tipos de neurotransmisores
Hay al menos 60-100 neurotransmisores y probablemente muchos otros aún por descubrir. Los más conocidos pueden agruparse en tipos en función de su estructura química.
Hoy en día, la mayoría de los neurocientíficos te dirán que la mayoría de las neuronas liberan el mismo neurotransmisor de sus axones, razón por la cual es posible que veas algunas neuronas denominadas “dopaminérgicas” o “serotoninérgicas”, liberando dopamina o serotonina, respectivamente. Pero un nuevo trabajo en el campo ha descubierto que las neuronas no son fijas cuando se trata de los químicos que liberan.
Algunas células cambian el tipo de neurotransmisores que liberan dependiendo de las circunstancias, a veces liberando hasta cinco tipos diferentes. Los científicos llaman a este fenómeno “cambio de neurotransmisores”.
Los neurotransmisores, al más alto nivel, se pueden clasificar en dos tipos: transmisores de molécula pequeña y neuropéptidos. Los transmisores de moléculas pequeñas, como la dopamina y el glutamato, suelen actuar directamente sobre las células vecinas. Los neuropéptidos, moléculas pequeñas como la insulina y la oxitocina, funcionan más sutilmente, modulando o ajustando la forma en que las células se comunican en la sinapsis. Estos poderosos neuroquímicos están en el centro de la neurotransmisión y, como tales, son críticos para la cognición y el comportamiento humanos.
A menudo, se habla de los neurotransmisores como si tuvieran un solo papel o función. La dopamina es un “químico del placer” y GABA es un neurotransmisor de “aprendizaje”. Pero los neurocientíficos están descubriendo que son multifacéticos y complejos, trabajando entre sí y en contra para facilitar la señalización neuronal a través de la corteza. Aquí hay una lista de algunos de los neurotransmisores más comunes discutidos en neurociencia.
Aminoácidos neurotransmisores
- Glutamato (GLU). Este es el neurotransmisor excitatorio más común y abundante. El glutamato tiene un papel importante en las funciones cognitivas como el pensamiento, el aprendizaje y la memoria. Demasiado glutamato resulta en excitotoxicidad, o la muerte de neuronas por accidente cerebrovascular, lesión cerebral traumática o esclerosis lateral amiotrófica (ELA), el trastorno neurodegenerativo debilitante más conocido como la enfermedad de Lou Gehrig. GLU también es importante para el aprendizaje y la memoria: la potenciación a largo plazo (LTP), ocurre en las neuronas glutamatérgicas en el hipocampo y la corteza.
- Ácido gamma-aminobutíco (GABA). GABA trabaja para inhibir la señalización neuronal. El GABA es el neurotransmisor inhibidor más común en el sistema nervioso, particularmente en el cerebro. Una nueva investigación sugiere que GABA ayuda a establecer circuitos cerebrales importantes en el desarrollo temprano. GABA también tiene un apodo: la “química de aprendizaje”. Los estudios han encontrado un vínculo entre los niveles de GABA en el cerebro y si el aprendizaje es exitoso o no.
- Glicina. La glicina es el neurotransmisor inhibidor más común en la médula espinal y participa en el procesamiento auditivo, el dolor y el metabolismo.
Neurotransmisores monoaminas
Los neurotransmisores monoaminas están involucrados en la conciencia, la cognición, la atención y la emoción.
- Serotonina (5HT). La serotonina es un neurotransmisor inhibidor. La serotonina ayuda a regular el estado de ánimo, patrones de sueño, sexualidad, ansiedad, apetito y dolor. Las enfermedades asociadas con el desequilibrio de serotonina incluyen trastorno afectivo estacional, ansiedad, depresión, fibromialgia y dolor crónico. Los medicamentos que regulan la serotonina y tratan estos trastornos incluyen inhibidores selectivos de la recaptación de serotonina (ISRS) e inhibidores de la recaptación de serotonina-norepinefrina (IRSN), que aumentan los niveles de transmisor al inhibir su recaptación después de haber realizado su trabajo en los sitios receptores postsinápticos. La serotonina (5HT), a veces llamada el “químico calmante”, es mejor conocida por sus efectos moduladores del estado de ánimo. La falta de 5HT se ha relacionado con depresión y trastornos neuropsiquiátricos relacionados. La 5HT también se ha implicado en facilitar la memoria y, más recientemente, en la toma de decisiones
- Histamina. La histamina regula las funciones corporales, incluyendo la vigilia, el comportamiento alimentario y la motivación.
- Dopamina (DA). La dopamina está involucrada en el sistema de recompensa del cerebro, los sentimientos de placer, la excitación cognitiva, el aprendizaje, el foco de atención, la concentración, la memoria, el sueño, el estado de ánimo y la motivación. La dopamina a menudo se conoce como el “químico del placer” porque se libera cuando los mamíferos reciben una recompensa en respuesta a su comportamiento; esa recompensa podría ser comida, drogas o sexo. Las enfermedades asociadas con disfunciones del sistema de dopamina incluyen la enfermedad de Parkinson, la esquizofrenia, la enfermedad bipolar, el síndrome de piernas inquietas y el trastorno por déficit de atención con hiperactividad (TDAH). Muchas drogas altamente adictivas (cocaína, metanfetaminas, anfetaminas) actúan directamente sobre los circuitos de dopamina del cerebro.
- Epinefrina. La epinefrina (también llamada adrenalina) y la norepinefrina (ver abajo) son responsables de la “respuesta de lucha o huida” al miedo y al estrés, activando el sistema nervioso simpático (ver Capítulo 4).
- Norepinefrina (NE). La norepinefrina es tanto una hormona como un neurotransmisor. Algunos se refieren a ella como noradrenalina. Se ha relacionado con el estado de ánimo, la excitación, la vigilancia, la memoria y el estrés. Investigaciones más recientes se han centrado en su papel tanto en el trastorno de estrés postraumático (TEPT) como en la enfermedad de Parkinson. La norepinefrina (noradrenalina) aumenta la presión arterial, la frecuencia cardíaca, el estado de alerta, la excitación, la atención y el enfoque. Muchos medicamentos (estimulantes y medicamentos para la depresión) aumentan la norepinefrina para mejorar el enfoque o la concentración para tratar el TDAH o para reducir los síntomas de la depresión.
Neurotransmisores peptídicos
Los péptidos son cadenas cortas de aminoácidos.
- Endorfinas. Las endorfinas son analgésicos naturales. Son sustancias naturales similares a los opiáceos, similares en propiedades moleculares a la morfina. La liberación de endorfinas reduce el dolor y eleva el estado de ánimo. Son opiáceos endógenos liberados por el hipotálamo y la glándula pituitaria durante el estrés o el dolor.
Acetilcolina (ACh)
Este neurotransmisor excitatorio se encuentra tanto en el sistema nervioso central como periférico, específicamente en el sistema nervioso autónomo y en las neuronas motoras espinales liberando acetilcolina sobre los músculos esqueléticos produciendo movimientos. También se involucra en la memoria, la motivación, el deseo sexual, el sueño y el aprendizaje. Las anomalías en los niveles de acetilcolina están asociadas con la enfermedad de Alzheimer. Pero también tiene otros papeles en el cerebro, incluyendo ayudar a dirigir la atención y desempeñar un papel clave en facilitar la neuroplasticidad a través de la corteza.
Otros Neurotransmisores
Los neuroquímicos como la oxitocina y la vasopresina también se clasifican como neurotransmisores. Elaborados y liberados del hipotálamo, actúan directamente sobre las neuronas y se han relacionado con la formación de vínculos de pareja, comportamientos monógamos y adicción a las drogas. Hormonas como el estrógeno y la testosterona también pueden funcionar como neurotransmisores e influir en la actividad sináptica.
Otros tipos de neurotransmisores incluyen el factor liberador de corticotropina (CRF), galanina, encefalina, dinorfina y neuropéptido Y. CRH, dinorfina y neuropéptido Y han sido implicados en la respuesta del cerebro al estrés. La galanina, la encefalina y el neuropéptido Y a menudo se denominan “cotransmisores”, porque se liberan y luego trabajan en asociación con otros neurotransmisores. La encefalina, por ejemplo, se libera con glutamato para señalar el deseo de comer y procesar recompensas.
A medida que los neurocientíficos están aprendiendo más sobre la complejidad de la neurotransmisión, es claro que el cerebro necesita estas diferentes moléculas para que pueda tener un mayor rango de flexibilidad y función.
Neurotransmisores de liberación de glía, también
Alguna vez se creía que solo las neuronas liberaban neurotransmisores. Nuevas investigaciones, sin embargo, han demostrado que la glía, las células que conforman el “pegamento” que llena el espacio entre las neuronas para ayudar a apoyar y mantener esas células, también tienen el poder de liberar neurotransmisores en las sinapsis. En 2004, los investigadores encontraron que las células gliales liberan glutamato en sinapsis en el hipocampo, ayudando a sincronizar la actividad de señalización.
Se sabe que los astrocitos, una célula glial en forma de estrella, liberan una variedad de neurotransmisores diferentes en la sinapsis para ayudar a fomentar la plasticidad sináptica, cuando es necesario. Los investigadores están trabajando diligentemente para comprender las contribuciones de estos diferentes tipos de células, y las moléculas neurotransmisoras que liberan, sobre cómo piensan, sienten y se comportan los humanos.
Diferentes tipos de receptores activados por el mismo transmisor
Existen diferentes tipos de receptores para transmisores de un tipo específico. Por ejemplo, para la dopamina existen al menos 5 tipos, receptores D1 a D5, todos para transmisores de dopamina. Diferentes tipos de receptores para un transmisor específico pueden localizarse en diferentes partes del cerebro y, por lo tanto, pueden producir diferentes efectos y tener diferentes funciones.
La función de cada tipo de receptor de dopamina (Mishra, et al., 2018):
- D1: memoria, atención, control de impulsos, regulación de la función renal (renal), locomoción (movimiento)
- D2: locomoción, atención, sueño, memoria, aprendizaje
- D3: cognición, control de impulsos, atención, sueño
- D4: cognición, control de impulsos, atención, sueño
- D5: toma de decisiones, cognición, atención, secreción de renina (por el riñón)
De particular interés, el trastorno mental esquizofrenia, caracterizado por pensamientos desordenados, alucinaciones y delirios, se asocia con un exceso de actividad de la neurona dopamina en el cerebro. Algunos tratamientos farmacológicos para la esquizofrenia disminuyen la actividad principalmente en los receptores D2.
Otro ejemplo es el transmisor de acetilcolina. Existen dos tipos distintos de receptores de acetilcolina afectados por dos sustancias diferentes, ya sea muscarina o nicotina. Aquellos receptores postsinápticos de acetilcolina que responden a la muscarina se denominan receptores muscarínicos. Aquellos que responden a la nicotina (en los productos de tabaco, por ejemplo) se llaman nicotínicos. Los receptores nicotínicos hacen que las neuronas simpáticas postganglionares y las neuronas posganglionares parasimpáticas se disparen y liberen sus sustancias químicas y hacen que el músculo esquelético se contraiga. Los receptores muscarínicos se asocian principalmente con funciones parasimpáticas y estimulan receptores localizados en tejidos periféricos (por ejemplo, glándulas, músculo liso). El transmisor de acetilcolina activa todos estos sitios.
Transmisión sináptica en revisión
Resumimos la secuencia de eventos en la transmisión sináptica en sinapsis con receptores ionotrópicos:
Potencial de acción en la neurona presináptica — liberación del transmisor — el transmisor cruza la brecha sináptica — el transmisor se conecta a sitios receptores postsinápticos — canales iónicos abiertos — los iones se mueven — resultados de PSP — si el PSP es un EPSP y si el EPSP es lo suficientemente grande (o si la suma da como resultado un cambio de voltaje grande suficiente), luego se alcanza un umbral desencadenante de -55mv en la neurona postsináptica —la neurona postsináptica produce su propio potencial de acción. Estos mismos pasos se resumen en los primeros 7 de los 8 pasos en transmisión sináptica enumerados anteriormente. El último paso es la inactivación del transmisor.
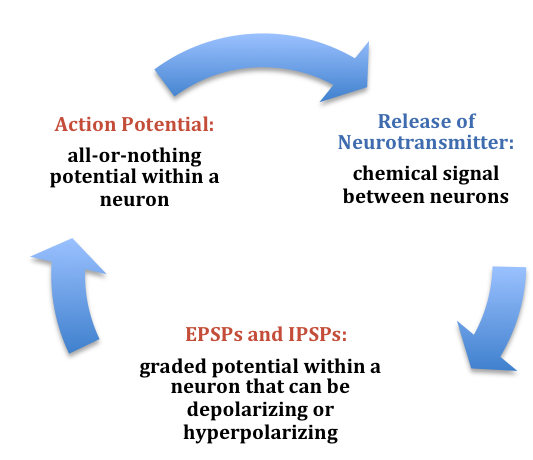
Figura\(\PageIndex{6}\): Resumen de la comunicación electroquímica dentro y entre neuronas. (Imagen de Furtak, S. (2021). Neuronas. En R. Biswas-Diener & E. Diener (Eds), serie de libros de texto Noba: Psicología. Champaign, IL: Editores DEF. Recuperado de http://noba.to/s678why4).
El paso 8 en la lista de 8 pasos anteriores simplemente se refiere al hecho de que después de que las moléculas transmisoras liberadas hayan hecho su trabajo, las moléculas transmisoras usadas deben limpiarse de la sinapsis. De no ser así, la neurona receptora se verá afectada durante demasiado tiempo y esto puede resultar en disfunción en la sinapsis, dando como resultado efectos patológicos, y si se producen en un gran número de sinapsis puede provocar anomalías como convulsiones o alucinaciones u otros efectos desadaptativos.
Como se señaló anteriormente, la eliminación del transmisor usado se logra mediante dos mecanismos, la recaptación y la destrucción enzimática. La recaptación significa que el transmisor usado se reabsorbe de nuevo en la terminación presináptica de la que se liberó. La destrucción enzimática significa que el transmisor usado es destruido químicamente por una enzima, y con ello inactivado.
Si el paso 8, inactivación del transmisor, no ocurriera, en el caso de los transmisores excitatorios, entonces la neurona possináptica estaría sobreactivada. Si esto ocurre a gran escala, en muchas sinapsis, como se señaló anteriormente, resultarán anomalías conductuales y mentales. Por ejemplo, existen algunos fármacos (Soman, Sarin, Malatión) que bloquean la destrucción enzimática del neurotransmisor Acetilcolina, ACh. Como se señaló anteriormente, la ACh es un transmisor involucrado en diversas funciones en el cerebro y el sistema nervioso periférico, incluyendo la estimulación de los músculos esqueléticos, los cuales son responsables del movimiento. Soman y Sarin son agentes nerviosos. Bloquean la enzima acetilcolinesterasa, AChE, y con ello evitan la destrucción enzimática de ACh en las sinapsis motoras que estimulan los músculos. Como resultado, el exceso de ACh en los músculos provoca su sobreactivación. ¿El resultado? Convulsiones epilépticas tan intensas que puede ocurrir la muerte. El tratamiento para el envenenamiento por estos agentes es un fármaco que bloquea el neurotransmisor de acetilcolina en los sitios receptores de acetilcolina, contrarrestando los efectos del exceso de neurotransmisor de acetilcolina, previniendo o revirtiendo las convulsiones que de otra manera ocasionan estos agentes nerviosos.
Neuronas, la mente y redes neuronales artificiales
¿Cómo se relacionan estos procesos neuronales con el mundo real de nuestra experiencia y comportamiento consciente cotidiano? Como se menciona en el módulo 5.2, en especies de vertebrados complejos, las células nerviosas individuales y su actividad no controlan un comportamiento ni crean un pensamiento o un sentimiento. En cambio, se procesa la información, se crean estados mentales y los comportamientos se organizan por eventos neuronales en circuitos complejos que involucran un gran número de neuronas. Es la interacción entre un gran número de neuronas, interconectadas por enormes cantidades de sinapsis, las que crean nuestras percepciones, pensamientos, emociones, o un comportamiento complejo como el habla humana o la creación de una obra de arte.
Se ha estudiado cómo grupos de neuronas en circuitos complejos pueden producir cosas complejas como una percepción o una memoria mediante modelado por computadora (ver Módulo 10.9). Las redes neuronales artificiales son modelos computacionales de circuitos neuronales y su funcionamiento. En un programa de computadora, las unidades artificiales de procesamiento de información, a veces llamadas “neurodas”, están programadas para codificar y procesar información similar a cómo las neuronas procesan la información. Estas unidades de procesamiento similares a neuronas están interconectadas entre sí y organizadas en capas, y luego estas capas de unidades de procesamiento se conectan.
Una red neuronal artificial de tres capas (ver Módulo 10.9) es capaz de realizar algunas tareas de procesamiento de información muy complejas, produciendo respuestas similares a las que podría producir un cerebro real. Además, debido a que los neurodos artificiales o unidades de procesamiento reciben (por programadores) la capacidad de alterar las fortalezas o “pesos” de sus “conexiones sinápticas”, las redes neuronales artificiales son capaces de aprender cuando se les da retroalimentación sobre su desempeño. Dadas estas propiedades, los investigadores (muchos de los cuales están en UC San Diego) han demostrado que las redes neuronales artificiales pueden aprender patrones y regularidades en insumos para hacer generalizaciones, formar categorías y resolver problemas complejos (Churchland, 2013).
Por ejemplo, una red neuronal, entrenada (electrónicamente) en los principios matemáticos utilizados para encontrar pruebas matemáticas, descubrió una prueba que había eludido a los matemáticos humanos durante muchas décadas. Otro, conocido como NetTalk, equipado con un sintetizador de voz y fotocélula “ojos” aprendió desde cero a leer y pronunciar correctamente el texto escrito en inglés. Logró esta hazaña asombrosa después de solo 10 horas de aprendizaje de prueba y error, durante las cuales se le dio retroalimentación sobre sus intentos de pronunciación. Pasó por una etapa de balbuceo inicialmente, como lo haría un niño, pero pronto fue articulando un discurso comprensible. Cuando se probó en material nuevo, generalizó lo que había aprendido al nuevo texto incluyendo muchas palabras nuevas que no había “visto” antes. Un análisis realizado por investigadores de cómo lo hizo mostró que había formado categorías de letras, aprendiendo a dividirlas en vocales y consonantes, con reglas de pronunciación para cada categoría. Esto lo logró sin programación directa por parte de los humanos. Extrae regularidades de las palabras que se le presentaron, e hizo generalizaciones, reglas que aplicó a nuevos casos (Churchland, 2013).
Las redes neuronales artificiales utilizan una forma de procesamiento de información llamada procesamiento distribuido paralelo (PDP). Aunque estas redes no son cerebros, se han acercado más que cualquier otro modelo de computadora a simular la función cerebral y las capacidades psicológicas de cerebros reales. Esto sugiere que los cerebros biológicos pueden utilizar PDP, realizado por redes PDP hechas de neuronas reales, para producir los tipos de capacidades psicológicas y conductuales que los cerebros reales como el nuestro demuestran. Se trata de un área muy prometedora de investigación que se está llevando a cabo en los departamentos de psicología, informática y neurociencia en muchas universidades del mundo.
Tenga en cuenta que los circuitos en los cerebros biológicos, como el cerebro humano, aunque modificables por la experiencia, también están cableados en gran medida por la selección natural y otras fuerzas de la evolución (ver Capítulo 3). Esto explica el hecho de que los seres humanos tienen una naturaleza humana innata o innata al igual que otros animales tienen su propia naturaleza innata. Los humanos se relacionan con el mundo de una manera distintivamente humana, mientras que los lobos, tigres, aves, ñus y otras especies animales entienden y se relacionan con el mundo en sus propias formas típicas de especie particular. La naturaleza psicológica distintiva de cada especie es el resultado de los circuitos cerebrales heredados de sus predecesores, circuitos moldeados por eones de evolución (Koenigshofer, 2011).
Referencias
Bekkers, J. M. (2011). Neuronas piramidales. Biología actual, 21 (24), R975.
Brown, A. M., Arancillo, M., Lin, T., Catt, D. R., Zhou, J., Lackey, E. P.,... & Sillitoe, R. V. (2019). Las interneuronas de capa molecular dan forma a la actividad de pico de las células cerebelosas de Purkinje. Informes científicos, 9 (1), 1-19.
Churchland, P. M. (2013). Materia y conciencia. Prensa MIT.
Fields, R. D., & Stevens-Graham, B. (2002). Nuevos conocimientos sobre la comunicación neurona-glía. Ciencia, 298 (5593), 556-562.
Kirkcaldie, M. T. (2012). Neocórtex. En El sistema nervioso del ratón (pp. 52-111). Prensa Académica.
Koenigshofer, K.A. (2011). Diseño de la mente: La organización adaptativa de la naturaleza humana, las mentes y el comportamiento. Educación Pearson, Boston.
Mishra A, Singh S, Shukla S. (2018). Bases fisiológicas y funcionales de los receptores de dopamina y su papel en la neurogénesis: posible implicación para la enfermedad de Parkinson. J Exp Neurosci. 12: 1179069518779829. [Artículo gratuito de PMC] [PubMed]
Nadim, F., & Bucher, D. (2014). Neuromodulación de neuronas y sinapsis. Opinión actual en neurobiología, 29, 48-56.
Pasternak, J. F., & Woolsey, T. A. (1975). Sobre la “selectividad” del método Golgi-Cox. J Comp Neurol, 160 (3), 307-312. doi: 10.1002/cne.901600304
Perea, G., Sur, M., & Araque, A. (2014). Redes neurona-glía: engranaje integral de la función cerebral. Fronteras en la neurociencia celular, 8, 378.
Smit, G. J., & Colon, E. J. (1969). Análisis cuantitativo de la corteza cerebral. I. Aelectividad de la técnica de tinción de Golgi-Cox. Brain Res, 13 (3), 485-510.
Sterling, P., & Laughlin, S. (2015). Principios del diseño neural. Prensa MIT.
Universidad de Queensland, Queensland Brain Institute, Tipos de neuronas, n.d. https://qbi.uq.edu.au/brain/brain-an... /tipos-neuronas; Recuperado 31/8/21
Van Essen, D., & Kelly, J. (1973). Correlación de la forma y función celular en la corteza visual del gato. Naturaleza, 241 (5389), 403-405.
Capítulo 5 Revisión
- Definir impulso nervioso.
- ¿Cuál es el potencial de reposo de una neurona y cómo se mantiene?
- Explicar cómo y por qué ocurre un potencial de acción.
- Describa cómo se transmite una señal de una célula presináptica a una célula postsináptica en una sinapsis química.
- ¿Qué determina generalmente los efectos de un neurotransmisor en una célula postsináptica?
- Identificar tres tipos generales de efectos que los neurotransmisores pueden tener sobre las células postsinápticas.
- Explicar cómo una señal eléctrica en una neurona presináptica provoca la transmisión de una señal química en la sinapsis.
- ¿El flujo de qué tipo de ion en la neurona resulta en un potencial de acción?
- ¿Cómo entran estos iones en la célula?
- ¿Qué hace este flujo de iones con la carga relativa dentro de la neurona en comparación con el exterior?
- La bomba de sodio-potasio:
- se activa por un potencial de acción
- requiere energía
- no requiere energía
- bombea iones de potasio fuera de las células
- Verdadero o Falso. Algunos potenciales de acción son mayores que otros, dependiendo de la cantidad de estimulación.
- Verdadero o Falso. Las vesículas sinápticas de la célula presináptica ingresan a la célula postsináptica.
- Verdadero o Falso. Un potencial de acción en una célula presináptica puede causar finalmente que la célula postsináptica se inhiba.
- Nombrar tres neurotransmisores.
Capítulo 5 Preguntas de discusión
- ¿Qué estructuras de una neurona son los principales canales de entrada y salida de esa neurona?
- ¿Qué significa la afirmación de que la comunicación dentro y entre celdas es un proceso electroquímico?
- ¿Cómo aumenta la mielina la velocidad y eficiencia del potencial de acción?
- ¿Cómo contribuyen la difusión (gradientes de concentración) y la presión electrostática (gradientes eléctricos) al potencial de la membrana en reposo y al potencial de acción?
- Describir el ciclo de comunicación dentro y entre neuronas.
Capítulo 5 Vocabulario
Potencial de acción (impulso nervioso)
Una corriente eléctrica positiva transitoria de todo o nada (despolarización) que se conduce hacia abajo del axón cuando el potencial de membrana alcanza el umbral de excitación (umbral de activación igual a aproximadamente 55 milivoltios negativos).
Axon
Parte de la neurona que se extiende fuera del soma, dividiéndose varias veces para conectarse con otras neuronas; salida principal de la neurona.
Membrana celular
Una capa bilipídica de moléculas que separa la célula del fluido extracelular circundante.
Dendrita
Parte de una neurona que se extiende alejándose del cuerpo celular y es la principal entrada a la neurona.
Difusión (a lo largo de un gradiente de concentración o desigualdad)
La fuerza sobre las moléculas para pasar de áreas de alta concentración a áreas de baja concentración.
Presión electrostática (gradiente de carga)
La fuerza sobre dos iones con carga similar para repelerse entre sí; la fuerza de dos iones con carga opuesta para atraer el uno al otro.
Potenciales postsinápticos excitatorios (EPSP)
Una corriente postsináptica despolarizante graduada que hace que el potencial de la membrana se vuelva más positivo y se mueva hacia el umbral de excitación (umbral de activación). Los EPSP y los IPSP se pueden sumar entre sí.
Potenciales postsinápticos inhibitorios (IPSP)
Una corriente postsináptica hiperpolarizante graduada que hace que el potencial de la membrana se vuelva más negativo y se aleje del umbral de excitación (umbral de activación). Los EPSP y los IPSP se pueden sumar entre sí.
Canales de iones
Proteínas que atraviesan la membrana celular, formando canales por los que pueden fluir iones específicos entre el espacio intracelular y extracelular (las “puertas” en el paso 5 en la lista de 8 pasos de transmisión sináptica anterior; ver Figura 5.3.2).
Receptor ionotrópico
Receptor (recibe moléculas transmisoras y se une a ellas) y su canal iónico asociado que se abre para permitir que los iones permeen la membrana celular en condiciones específicas, como la presencia de un neurotransmisor (canal controlado químicamente) o un potencial de membrana específico (canal controlado por voltaje).
Envoltura de mielina
Sustancia alrededor del axón de una neurona que sirve como aislante para permitir que el potencial de acción conduzca rápidamente hacia los botones terminales.
Neurotransmisores o transmisores
Sustancia química liberada por el botón terminal presináptico que actúa sobre la célula postsináptica.
Núcleo
Colección de células nerviosas que se encuentran en el cerebro que normalmente cumplen una función específica.
Potencial de membrana en reposo
El voltaje dentro de la celda relativo al voltaje fuera de la celda mientras que la celda es un descanso (aproximadamente -70 mV).
Bomba de sodio-potasio
Un canal iónico que utiliza la energía de la neurona (trifosfato de adenosina, ATP) para bombear tres iones Na+ fuera de la célula a cambio de traer dos iones K+ dentro de la célula.
Soma
Cuerpo celular de una neurona que contiene el núcleo y la información genética, y dirige la síntesis de proteínas.
Espinas
Protusiones en la dendrita de una neurona que forman sinapsis con botones terminales del axón presináptico.
Sinapsis
Unión entre el botón terminal presináptico de una neurona y la dendrita, axón o soma de otra neurona postsináptica.
Brecha sináptica o espacio sináptico
También conocida como hendidura sináptica; el pequeño espacio entre el botón terminal presináptico (terminación axónica) y la columna dendrítica postsináptica, axón o soma.
Vesículas sinápticas
Grupos de neurotransmisores empaquetados juntos y ubicados dentro del botón terminal.
Botón de terminal o terminación de axón
La parte del extremo del axón que forma sinapsis con dendrita postsináptica, axón o soma.
Umbral de activación (Umbral de excitación)
Potencial específico de membrana que la neurona debe alcanzar para iniciar un potencial de acción.
Recursos Externos
Video Resumen: https://www.stem.org.uk/system/files...he%20brain.mp4
Atribuciones
1. Capítulo 5, Comunicación dentro del Sistema Nervioso, Módulo 5.3. Neuronas y transmisión sináptica por Kenneth A. Koenigshofer, PhD, Chaffey College, está licenciado bajo CC BY 4.0
2. “Tipos de Neurotransmisores” y “Glia Release Transmisores Too” adaptado por Kenneth A. Koenigshofer de Sukel, K (2019) Neurotransmisores de https://dana.org/article/neurotransmitters/
2. Figura 5.3.3, Vocabulario, Preguntas de Discusión, Recursos Externos, y algún texto adaptado por Kenneth A. Koenigshofer de Furtak, S. (2021). Neuronas. En R. Biswas-Diener & E. Diener (Eds), serie de libros de texto Noba: Psicología. Champaign, IL: Editores DEF. Recuperado a partir de http://noba.to/s678why4
Licencia Creative Commons



 Neuronas de Sharon Furtak está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Los permisos más allá del alcance de esta licencia pueden estar disponibles en nuestro Acuerdo de licencia.
Neuronas de Sharon Furtak está bajo una Licencia Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional. Los permisos más allá del alcance de esta licencia pueden estar disponibles en nuestro Acuerdo de licencia.
3. Figuras 5.3.1, 5.3.2 y algún texto adaptado por Kenneth A. Koenigshofer de: General Biology (Boundless), Capítulo 35.2C, El sistema nervioso; transmisión sináptica, https://bio.libretexts.org/Bookshelv...gy_(Boundless); el contenido de LibreTexts está licenciado por CC BY-NC-SA 3.0. Jurídico.
4. Figuras 5.2.2 y “Capítulo 5 Revisión” adaptadas por Kenneth A. Koenigshofer del Capítulo 11.3 (Neuronas y Células Gliales), 11.4 (Impulsos Nerviosos) en Libro: Biología Humana (Wakim & Grewal) - LibreTextos de Biología por Suzanne Wakim & Mandeep Grewal, bajo licencia CC BY- NC
Recursos Externos para el Capítulo 5
- Serie de videos: Neurobiología/Biopsicología - Animaciones tutoriales de potenciales de acción, potenciales de membrana en reposo y transmisión sináptica.
- http://www.sumanasinc.com/webcontent/animations/neurobiology.html
- Video: Una animación y una explicación de un potencial de acción
- Video: Una animación de las acciones de los neurotransmisores en la sinapsis
- Video: Una animación interactiva que permite a los estudiantes observar los resultados de manipulaciones a potenciales postsinápticos excitatorios e inhibitorios. También incluye animaciones y explicaciones de transmisión y circuitos neuronales.
- https://apps.childrenshospital.org/clinical/animation/neuron/
- Video: Otra animación de un potencial de acción
- Video: Otra animación de las acciones de los neurotransmisores en la sinapsis
- Video: Potencial de acción dominó: Esta actividad práctica ayuda a los estudiantes a comprender el complejo proceso del potencial de acción, así como a familiarizarse con las características de transmisión (por ejemplo, respuesta de todo o ninguno, período refractario).
- Video: Por perspectiva sobre técnicas en neurociencia para mirar dentro del cerebro
- Video: El cerebro comportándose es el tercer programa de la serie DISCOVERING PSICHOLOGY. Este programa analiza la estructura y composición del cerebro humano: cómo funcionan las neuronas, cómo se recopila y transmite la información, y cómo las reacciones químicas se relacionan con el pensamiento y el comportamiento.
- http://www.learner.org/series/discoveringpsychology/03/e03expand.html
- Video: Se pueden cultivar nuevas células cerebrales. Aquí\\'s cómo. - ¿Podemos, como adultos, crecer nuevas neuronas? La neurocientífica Sandrine Thuret dice que podemos, y ofrece investigación y consejos prácticos sobre cómo podemos ayudar a nuestros cerebros a realizar mejor la neurogénesis, mejorando el estado de ánimo, aumentando la formación de memoria y previniendo el declive asociado con el envejecimiento en el camino.
- Web: Para más información sobre el Premio Nobel compartido por Ramón y Cajal y Golgi
- http://www.nobelprize.org/nobel_prizes/medicine/laureates/1906/